- Nuklearpolyhedrose-Virus
-
Baculovirus  Baculoviren
BaculovirenSystematik Reich: Virus Baltimore K. (dsDNA-Viren) I Familie: Baculoviridae (Baculoviren) Wissenschaftlicher Name Baculovirus Gattungen - Granulovirus
- Nucleopolyhedrovirus
Die Familie der Baculoviren umfasst doppelsträngige, filamentöse DNA-Viren. Sie befallen ausschließlich Wirbellose, ihr Hauptwirt sind Mottenlarven, jedoch sind an die 600 Wirte beschrieben. Baculoviren werden in der Gentechnologie zur Erzeugung von komplexen Eukaryotenproteinen in Zellkulturen sowie zur Vektorklonierung verwendet. Auch werden sie in der Landwirtschaft zur Kontrolle von Schadinsekten eingesetzt.
Der Name "Baculo" geht auf lat. baculum = Stock, Stab zurück und bezieht sich auf die Morphologie des Nukleocapsids [1]. Innerhalb der Familie werden zwei Genera unterschieden: Nucleopolyhedrovirus (NPV) und Granulovirus (GV). Beim Granulovirus kommt pro Hülle nur ein Nukleokapsid, beim NPV kommen entweder einzelne (SNPV) oder mehrere (MNPV) Nukleokapside pro Hülle vor. Die Virionen (Nukleokapsid und Hülle) sind nun abermals in eine Granulin (GV)- oder Polyhedrin (NPV)-Matrix eingebettet. Auch hat die Art der Granuloviren nur ein Virion pro Granulinmatrix, bei Nucleopolyhedroviren sind mehrere Virionen in der Polyhedrinmatrix.
Inhaltsverzeichnis
Verbreitung
Sie sind weltweit verbreitet und wurden erstmals im 16. Jahrhundert beschrieben, als "Welkkrankeit" bei Seidenraupen. In den 1940er Jahren wurden sie als Biopestizide in Getreidefeldern eingesetzt, um Schadinsekten zu dezimieren. Durch spätere Untersuchungen wurden sie in den 1990er Jahren zur Produktion von Eukaryonten-Proteinen eingesetzt, da sie einfach in Zellkulturen vermehrt werden können. Die Viren sind an Wirbellose angepasst mit über 600 möglichen Wirtsarten. Hauptsächlich befallen sie Mottenlarven, aber sie wurden auch bei Pflanzenwespen, Moskitos und Shrimps gefunden.
Lebenszyklus
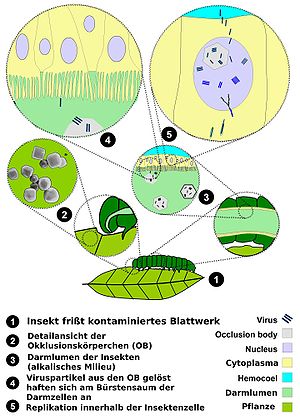 Lebenszyklus der Baculoviren
Lebenszyklus der BaculovirenIn seinem Lebenszyklus kommt das Virus in zwei Formen vor, dem ODV (Occlusion Derived Virus), mit Proteinhülle, und dem BV (Budding Virus). Die ODV-Form ist für die Erstinfektion notwendig, später werden dann BV-Partikel gebildet.
Der Hauptinfektionsweg verläuft über kontaminierte Blätter, die von Insekten gefressen werden. Das Virus gelangt in den Verdauungstrakt, wo sich die Proteinhülle (ODV-Form) aufgrund der basischen Umgebung auflöst. Die freien Viren heften sich nun an die Darmepithelzellen und werden über Endocytose in ein Endosom aufgenommen, aus dem sie sich befreien und nun als Nukleokapside, vermutlich durch Aktin-Filamente, in den Zellkern transportiert werden. Hier erfolgt die Replikation des Virus. Nun werden unbehüllte Viren (BV) erzeugt und das Virus infiziert die umliegenden (basolateralen) Epithelzellen. Die BV-Partikel umhüllen sich jedoch auch locker mit Zellmembran, die Glycoproteine des Virus enthält.
Die Infektion kann in drei Phasen unterteilt werden: die frühe Phase (0–6 Stunden), die späte Phase (6–24h) und die sehr späte Phase (18–24 bis 72h nach der Infektion). Bis zur späten Phase werden noch unbehüllte BV Formen freigesetzt, in der letzten Phase jedoch nur ODV Formen, die durch die Kernmembran knospen und mit Viren gefüllte Partikel bilden, die mit OB-(Occlusion Body)-Protein gefüllt sind. Diese besitzen nun eine größere Resistenz gegen Umweltfaktoren als die BV-Form und dienen der Verbreitung des Virus auf den nächsten Wirt. Da beim Austritt der Viren die Zelle lysiert wird, kommt es im gesamten Insekt zu einer regelrechten Verflüssigung, wobei nur die Chitinhülle intakt bleibt. Diese platzt später auf und verstreut die Viren auf der gesamten Blattoberfläche. Diese Eigenschaft gab der durch dieses Virus verursachten Erkrankung auch den englischen Namen "wilting disease" (Welkkrankheit).
Struktur des Virions
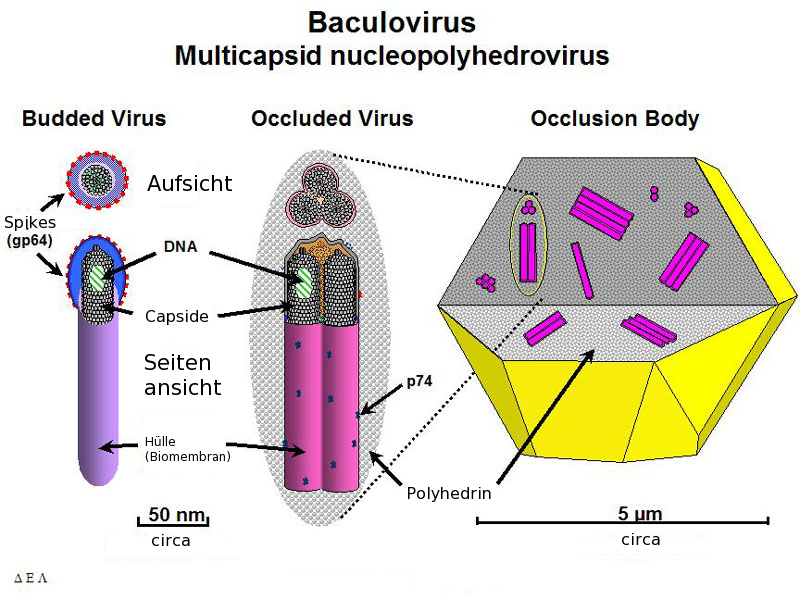
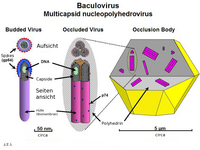 Formen der Baculoviren
Formen der BaculovirenDas bestuntersuchte Baculovirus ist Autographa californica multicapsid nucleopolyhedrovirus (AcMNPV). Es wurde erstmals aus einer Gammaeulenart (Autographa californica) isoliert, die auf Luzerne lebt. Das Genom hat 134 kBp (kilo Basenpaare) und besitzt 154 ORF. Das wichtigste Kapsidprotein ist VP39, es bildet mit einigen kleineren Proteinen das Nukleokapsid (21 nm x 260 nm), das die DNA umhüllt, die wiederum vom Protein p6.9 umgeben ist.
Die BV Partikel benötigen für eine Infektion das Glycoprotein gp64 (Genprodukt 64), welches an den Enden des fadenförmigen Virions vorkommt. Dieses Protein findet sich nicht in den ODV-Partikeln, hier gibt es einige Proteine die nur bei ODV vorkommen. Unterschiede gibt es auch in der Zusammensetzung der Lipide der Hülle, bei BV kommt Phosphatidylserin vor, ODV enthält Phosphatidylcholin und Phosphatidylethanolamin.
Hüllprotein gp64
Im Laufe der Evolution des Virus hat sich das gp64 verändert. Ld139, (auch Baculovirus F-Protein) aus dem Schwammspinner Lymantria dispar (LdMNPV) ist vermutlich ein ursprüngliches Membranfusionsprotein (zur Freisetzung des Virus aus der Wirtszelle), das durch gp64 als nicht "gleichwertiges" Protein ersetzt wurde, da Ld139 und gp64 im Genom vieler Baculovirus Arten vorkommen (Maulbeerspinner, Gammaeule, Douglasien-Bürstenspinner (Familie der Trägspinner)).
Gp64 selbst ist ein homotrimeres Protein das an den Polen der filamentösen BV Partikeln sitzt. Es besteht aus 512 Aminosäuren, mit vier Glycosilierungsstellen an Asparagin Resten und eine N-Terminale Signalsequenz von 20 Aminosäuren, weiters Oligomerisations und Fusionsdomänen, sowie eine hydrophobe Transmembrandomäne am C-Terminalen Ende von sieben Aminosäuren. Es wird in der frühen und in der Späten Phase der Infektion gebildet mit einem Maximum nach 24 bis 26 Stunden nach der Infektion. Die Trimerisation (Zusammenlagerung von je 3 gp64 Einheiten) über interne Cystinbindungen ist wichtig für den Transport zur Zelloberfläche, da nur 33 Prozent des gp64 die Oberfläche erreichen und Monomere innerhalb der Zelle abgebaut werden.
Dieses Protein ist essentiell für die Knospung des Virions aus der Zelle, sowie für die Infektion von weiteren Zellen im Infektionszyklus. Seine Hauptaufgabe ist die über den pH-Wert regulierte Fusion mit dem Endosom. Obwohl gp64 notwendig ist für die Virusvermehrung können Mutanten denen gp64 fehlt, über Substitution mit Ld130, oder dem G-protein des "Vesikulären Stomatitis" Virus wieder funktionsfähig gemacht werden.
Quellen


Wikimedia Foundation.