- Wollhaarmammut
-
Wollhaarmammut 
Skelettrekonstruktion eines Wollhaarmammuts im Mammutheum in Siegsdorf Bayern.
Systematik Klasse: Säugetiere (Mammalia) Überordnung: Afrotheria Ordnung: Rüsseltiere (Proboscidea) Familie: Elefanten (Elephantidae) Gattung: Mammuts (Mammuthus) Art: Wollhaarmammut Wissenschaftlicher Name Mammuthus primigenius Blumenbach, 1799 Das Wollhaarmammut, auch Woll- oder Fellmammut (Mammuthus primigenius), ist eine ausgestorbene Art aus der Familie der Elefanten. Diese Art der Mammuts entwickelte sich im Übergang vom Alt- zum Mittelpleistozän vor etwa 800.000 bis 600.000 Jahren in Sibirien und bewohnte die kaltzeitlichen Steppen im nördlichen Eurasien und Nordamerika. Es stellt das Endglied der plio- und pleistozänen Entwicklung der Mammuts dar. Am Ende des Pleistozäns starb das Wollhaarmammut in weiten Teilen seines Verbreitungsgebietes aus, überlebte aber regional in einigen Restpopulationen bis ins mittlere Holozän.
Inhaltsverzeichnis
Verbreitung und Lebensraum
 Lebensrekonstruktion eines Wollhaarmammuts.
Lebensrekonstruktion eines Wollhaarmammuts.
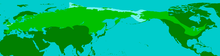 Maximale Verbreitung des Wollhaarmammuts während der Weichsel-Kaltzeit.
Maximale Verbreitung des Wollhaarmammuts während der Weichsel-Kaltzeit.Verbreitungsgebiet
Das Wollhaarmammut war ein an die Kälte angepasstes Tier und bevorzugte boreales bis subarktisches Klima. Es entstand vor rund 800.000 bis 600.000 Jahren im Innern Sibiriens und breitete sich im späten Mittelpleistozän vor rund 300.000 bis 250.000 Jahren in Eurasien aus. In Mitteleuropa ist es in der Saale-Kaltzeit (vor 300.000 bis 126.000 Jahren) nachgewiesen, u. a. aus dem Geiseltal (Sachsen-Anhalt). Am Ende dieser Vereisungsperiode zog sich die Tierart aber recht schnell wieder in ihre Ursprungsgebiete zurück.[1]
Im Ausgang der Eem-Warmzeit (vor 126.000 bis 115.000 Jahren) begann das Wollhaarmammut eine erneute Expansionsphase Richtung Süden, Südwesten und Südosten aber auch in den Norden. Dass diese zweite Ausbreitungswelle sehr zügig vonstatten gegangen sein muss, zeigen Funde aus Averley (Essex, England), wo es bereits in späten warmzeitlichen Sedimenten nachgewiesen ist. In der darauf folgenden Weichsel-Kaltzeit (vor 115.000 bis 11.700 Jahren) erreichte das Verbreitungsgebiet des Wollhaarmammuts seine maximale Ausdehnung. So trat die Tierart im westlichen Eurasien flächendeckend von West- über Mittel- bis Ost- und den südlichen Teil Nordeuropas auf, Ausnahmen blieben die eisbedeckten Gebiete wie der nördliche Teil von Fennoskandinavien. Darüber hinaus drang es auch nach Südeuropa vor, wo es auf der Iberischen Halbinsel in Granada (Spanien) bei 37° nördlicher Breite seinen südlichsten Punkt in Europa erreichte. Weiterhin ist es aber auch bis auf die Apennin-, die Balkanhalbinsel und bis zum Schwarzen Meer vorgestoßen.[2][3]
In Westasien erreichte das Wollhaarmammut die südliche Kaukasusregion, während es in Zentralasien unter Umgehung der Wüstenregionen der Kysylkum auch in Kasachstan auftrat. Weiter im Osten besiedelte es auch den nördlichen Teil der Mongolei. In Ostasien drang es in den Nordosten Chinas vor, wo seine südlichsten Fundstellen am Huang He etwa bei 35° bis 36° nördlicher Breite liegen, was gleichzeitig die weltweit südlichste Verbreitungsgrenze dieser Rüsseltierart darstellt. Im äußersten Osten Asiens sind Funde vom nördlichen Teil der koreanischen Halbinsel, von Kamtschatka, Sachalin (beides Russland) und von Hokkaido (Japan) bekannt.[1][2]
Weiterhin erfolgte eine Nordexpansion, in derem Zuge Jakutien und der hohe Norden und Nordosten Asiens besiedelt wurde, wie etwa die Taimyr-Halbinsel und die Wrangel-Insel. Die Taimyr-Halbinsel stellt dabei die oberste Verbreitungsgrenze des Wollhaarmammuts dar, welche ungefähr beim 76. nördlichen Breitengrad lag.[4] Die Tschuktschen-Halbinsel muss das Wollharmammut bereits vor rund 100.000 Jahren erreicht haben. Diese weite nordosteurasische Ausdehnung war notwendig, um über die Beringstraße nach Nordamerika einzuwandern.[5]
In Nordamerika war das Wollhaarmammut vor allem in Alaska und im nördlichen Kanada verbreitet, während es im Osten die Großen Seen erreichte. Von dort aus drang es in die Präriegebiete und die Großen Ebenen vor. Hier befinden sich auch die südlichsten amerikanischen Verbreitungsgrenzen, die etwa bei 36° bis 38° nördlicher Breite lagen. In den westlichen Teil Nordamerikas konnte das Rüsseltier nicht vordringen, weil die Kordilleren damals eisbedeckt waren. Ursprünglich aus Mexiko berichtete Reste des Wollhaarmammuts stellen offensichtlich Verwechslungen mit dem Präriemammut (Mammuthus columbi) dar.[2]
Am Ende der Weichsel-Kaltzeit zog sich das Wollhaarnashorn wieder aus seinen neuen Refugien zurück, starb aber alsbald sowohl in Eurasien als auch in Nordamerika weitgehend aus. Einige Restpopulationen haben allerdings noch bis weit in das Holozän verstreut überlebt. Unter diesen Populationen bestand eine isolierte auf der Wrangel-Insel, eine weitere auf der Sankt-Paul-Insel (Alaska), die einen Teil der Beringstraße bildet, und zwei weitere auf der Tschuktschen-Halbinsel bzw. der Taimyr-Halbinsel des eurasischen Festlandes.[6]
Lebensraum
Das Wollhaarmammut bevorzugte offene Landschaften und war an die Hartgras- und Strauchvegetation der kaltzeitlichen Lösssteppe und Steppentundra angepasst. Aus diesem Grund besiedelte es überwiegend Flachlandregionen und Niederungen, kam unter für ihn günstigen Klimabedingungen auch in Hochlagen und gebirgigem Gelände vor, mied aber weitgehend Wüstengebiete. Besonders häufig trat es in wasserreichen Arealen, wie Flusstälern oder Seeufern auf.[7][8] Aufgrund der hohen Verbreitung während der letzten Kaltzeit lebte es in seinen südlichen Verbreitungsgebieten unter Umständen auch in einem stärker bewaldeten oder parkähnlichen, aber immer noch deutlich steppenartigen Biotop.[2][8] Während wärmeren Klimaphasen der letzten Vereisungsperiode bewohnte es im westlichen Russland ebenfalls ein waldsteppenartiges Habitat.[9]
Das Wollhaarmammut war nicht nur Namensgeber sondern auch ein wichtiger Bewohner der Mammutsteppe, die weite Teile Eurasiens und Nordamerikas während der Kaltzeiten einnahm. Diese Mammutsteppe, eine Mischform aus Steppen- und Tundrenvegetation, war charakterisiert durch eine hohe Sonneneinstrahlung und eine, hervorgerufen durch die nahe Lage an den Gletscherrändern, vorherrschende lang andauernde Hochdrucklage. Dies resultierte in der Ausbildung einer nährstoffreichen Pflanzendecke. In diesem unter den heutigen Klimabedingungen nicht mehr existierenden Landschaftsraum lebte der in den Kaltzeiten des späten Mittel- und des Jungpleistozäns auftretende s. g. Mammuthus-Coelodonta-Faunenkomplex, dessen Charaktertier neben dem Wollnashorn auch das Wollhaarmammut war. Andere Begleitelemente dieses Faunenkomplexes waren der Bison, das Ren, die Saiga-Antilope und der Moschusochse.[10][7]
Bedingt durch den langen Zeitrahmen und dem großen Verbreitungsgebiet koexistierte das Wollhaarmammut gelegentlich mit andern Mammut- und Rüsseltierarten. Vor allem zu Beginn der Saale-Kaltzeit im westlichen Eurasien trat es noch zusammen mit dem Steppenmammut (Mammuthus trogontherii) auf, welches allerdings bald darauf hier ausstirbt.[1] Da sich diese ältere Mammutform im östlichen Eurasien, vor allem in China, jedoch noch bis ins späte Jungpleistozän gehalten zu haben scheint - hier wird sie in ihrer Spätform gelegentlich, aber fälschlicherweise Mammuthus sungari genannt - kommen sowohl Wollhaar- als auch Steppenmammut immer wieder an einzelnen Fundstellen gemeinsam vor .[11] In Nordamerika traten überwiegend an den südlichen Verbreitungsgrenzen des Wollhaarmammuts Überschneidungen zum Präriemammut auf, welches aber allgemein einen eher westlicheren und südlicheren Lebensraum hatte.[2][5] Darüber hinaus gab es hier auch eine gemeinsame Habitatnutzung mit dem ebenfalls bis ins Jungpleistozän vorkommenden Amerikanischen Mastodon (Mammut americanum), welches jedoch keinen Vertreter der Elefanten darstellt, sondern der wesentlich urtümlicheren Rüsseltierform der Echten Mastodonten angehört.[12]
Ob das Wollhaarmammut auch mit dem an Warmzeiten angepassten Europäischen Waldelefanten (Elephas antiquus) zumindest in den Spätphasen dieser Klimaabschnitte gemeinsam auftrat, ist unklar. Die rasche Wiederbesiedlung des westlichen Eurasiens durch das Wollhaarmammut in der späten Eem-Warmzeit macht wahrscheinlich, dass die äußersten Besiedlungsgrenzen beider Rüsseltiere geographisch nicht sehr weit auseinanderlagen. Ebenso möglich ist, das bei seiner südlichen Ausbreitung das Wollhaarmammut wiederum sehr nah an die letzten Rückzugsgebiete des Europäischen Waldelefanten in Südeuropa heranrückte.[1]
Funde des Wollhaarmammuts
Fundstellen allgemein
 Skelettrekonstruktion eines Wollhaarmammuts im Museum für Geologie in Warschau.
Skelettrekonstruktion eines Wollhaarmammuts im Museum für Geologie in Warschau.Fundstellen des Wollhaarmammuts liegen aufgrund des großen Verbreitungsgebietes mannigfaltig vor. Meist werden nur die widerstandsfähigsten Skelettelemente, wie die Zähne gefunden, seltener sind es Skelettpartien oder gar vollständige Tiere. Dabei kommen viele Funde in Kies-, Sand- und Tongruben vor, deren Ablagerungen ehemalige Flussläufe oder Seebecken aufzeigen. Gelegentlich sind sie auch in Höhlen oder Erdfällen zu finden oder submarin am Boden der Nord- und Ostsee bzw. des küstennahen Arktischen Ozeans, deren Schelfgebiete während der letzten Kaltzeit aufgrund des niedrigeren Meeresspiegels trocken gefallen waren.
Ein sehr wichtiger und für die Mammutforschung hervorragender Fundplatz stellt der Mammutfriedhof in einer Flussschlinge des Bjorjoljoch in Sibirien dar, der 1970 entdeckt und auf rund 12.000 Jahre datiert wurde. Der gesamte Friedhof enthielt mehr als 8.800 Mammutknochen von insgesamt 156 Individuen. Herausragend dabei ist ein 175 cm langes Hinterbein eines Wollhaarmammuts, das noch komplett mit Haut und Haaren bedeckt war. Warum hier so viele Tiere starben, ist strittig, da aber mehr weibliche Tiere vorliegen, geht man davon aus, dass mehrere Herden oder ein großer Herdenverband bei der Überquerung des Flusses ertranken. Da vollständige Skelette fehlen, fand das Ereignis höchstwahrscheinlich an einer anderen Stelle flussauf statt.[13][14] Ein ähnlicher, nicht ganz so umfangreicher Friedhof kam 1988 am Ufer der Sewa südlich von Moskau zu Tage, wo insgesamt 4.000 Knochenfragmenten von 10 bis 15 Individuen aller Altergruppen gefunden wurden. Das Alter der Fundstelle wird auf 13.950 Jahre datiert.[5]
Auch aus Deutschland sind zahlreiche Fundstellen bekannt, es liegen aber insgesamt nur sechs vollständige Skelette vor. Das forschungsgeschichtlich älteste stammt aus Klinge bei Cottbus (Brandenburg) und wurde 1903 gefunden. Ein weiteres wurde 1909 bei Borna nahe Leipzig (Sachsen) geborgen. Es war im dortigen Völkerkundemuseum ausgestellt, während des Zweiten Weltkrieges aber zerstört worden. Das Skelett eines 3,2 m hohen Wollhaarmammuts kam 1910 bei Ahlen (Nordrhein-Westfalen) zum Vorschein und steht heute im Geologisch-Paläontologischen Museum der Westfälischen Wilhelms-Universität in Münster, während ein weiteres 1936 an der Autobahn Koblenz - Trier bei Polch ausgegraben wurde. Das Mammut von Pfännerhall kam 1953 bei Tagebauarbeiten im Geiseltal zum Vorschein und ist heute im Landesmuseum für Vorgeschichte in Halle (Sachsen-Anhalt) ausgestellt. Der jüngste Fund eines 3,6 m hohen Wollhaarmammuts stammt aus Siegsdorf bei Traunstein (Bayern) und wurde 1975 entdeckt, aber erst zehn Jahre später vollständig ausgegraben. Es steht heute im Museum Mammutheum am Fundort.[10][15]
Funde mumifizierter Wollhaarmammutkadaver
 Die Dermoplastik des Berjosowka-Mammuts in Fundstellung, ausgestellt im Zoologischen Museum in Sankt Petersburg.
Die Dermoplastik des Berjosowka-Mammuts in Fundstellung, ausgestellt im Zoologischen Museum in Sankt Petersburg. Das Mammutkalb „Dima“ am Fundort in der einstigen Mammutsteppe von Beringia
Das Mammutkalb „Dima“ am Fundort in der einstigen Mammutsteppe von BeringiaIm Eis des seit der letzten Vereisungsphase nicht oder kaum aufgetauten Permafrostbodens jenseits des 60. nördlichen Breitengrades sind sowohl im nördlichen und nordöstlichen Sibirien (hauptsächlich in Jakutien) als auch im nördlichen Nordamerika (Alaska, Kanada) konservierte Wollhaarmammuts gefunden worden. Solche Funde sind besonders wichtig, um das Aussehen, die Ernährungs- und auch die Lebensweise dieser pleistozänen Rüsseltierart zu rekonstruieren.
Einer der frühesten wissenschaftlich untersuchten Kadaver wurde 1799 vom tungusischen Elfenbeinhändler Ossip Schumachow im Delta der Lena gefunden. Dieses war während eines Erdrutsches teilweise, im Sommer 1805 durch weitere Erosion aber erst vollständig freigelegt worden. Schumachow beraubte den Kadaver der Stoßzähne und verkaufte sie dem jakutischen Händler Roman Boltunow. Dieser reiste zum Fundort und fertigte eine Skizze des Kadavers an, der abgesehen von Raubtierfraß an Rüssel und Ohren vollständig war. Diese Skizze gelangte letztendlich an die Russische Akademie der Wissenschaften in Sankt Petersburg, wo sie der englische Naturforscher Henry Michail Adams einsah. Adams organisierte eine Expedition zum Fundgebiet, zu dem ihn Schumachow 1806 führte. Der Mammutkadaver war zu diesem Zeitpunkt bereits zur Hälfte Opfer von Raubtieren geworden, die linke Körperseite wies aber noch eine gute Haar- und Hautkonservierung auf. Bei der Bergung des Skelettes gingen aber bis auf die Kopfhaut und die Fußsohlen alle Weichteile verloren. Das Skelett wurde anschließend nach Sankt Petersburg gebracht und dort seit 1808 in der Kunstkammer aufgestellt. Es ist somit das erste montierte Wollhaarmammut-Skelett weltweit und wird nach seinem Ausgräber als Adams-Mammut bezeichnet.[13][5]
Erst 1846 wurde von russischen Landvermessern am Indigirka-Fluss in Sibirien erneut ein gut erhaltener Kadaver gefunden, an dem sich noch Fellreste befanden und dessen Mageninhalt erhalten war. Der Kadaver war durch das Frühjahrshochwasser freigespült worden und steckte mit den Hinterbeinen noch im Erdreich. Bei der Bergung der Karkasse allerdings ging durch den sofort einsetzenden Verwitterungsprozeß ein Großteil der Weichteile verloren.[16] Im Jahre 1900 fand man das nach seinem Fundort an einem Nebenfluss der Kolyma in Nordostsibirien benannte Berjosowka-Mammut. Eine im folgenden Jahr von der Akademie der Wissenschaften ausgestattete Expedition konnte das Mammut erfolgreich bergen, welches in sitzender Haltung im Erdreich steckte. Außer der durch Wölfe, Bären oder Füchse angefressene Rüssel und Kopf war es vollständig und besaß neben der Zunge auch noch bis zu 15 kg Nahrungsreste im Magen. Eine Dermoplastik sowie das Skelett des Mammutbullen sind heute im Zoologischen Museum der Russischen Akademie der Wissenschaften in Sankt Petersburg zugänglich.[13]
In den folgenden Jahren gelangen mehrere Funde von Mammutkadavern. Eines wurde zwischen 1901 und 1903 auf den Ljachow-Inseln geborgen und kurz bevor ein kaiserlicher Erlass die Ausfuhr von Mammutknochen aus Russland verbot nach Paris geschafft, wo es im Jardin des Plantes ausgestellt ist. Eugen Pfitzenmayer, der auch schon an der Bergung und anschließenden Restaurierung des Berjosowka-Mammuts beteiligt war, fand 1908 einen weiteren allerdings schwer beschädigten Kadaver am Sanga Jurach in Jakutien. Dieses Exemplar stellt das erste mit vollständig erhaltenem Rüssel dar.[5] Schon ein Jahr später wurde die Entdeckung eines vollständigen Kadavers im Flusstal der Mochowaja auf der Taimyr-Halbinsel gemeldet und einige Hautfetzen geborgen. Eine 1913 dorthin gesendetete Expedition unter Leitung von G. N. Kutomanow fand aber nur noch die Hälfte vor, die andere war zuvor von einheimischen Jägern als Hundenahrung verwendet worden. Der Rest dieses heute als Kutomanow-Mammut bekannten Fundes wurde geborgen und nach Sankt Petersburg gebracht.[4]
Ein vorzüglich erhaltenes partiell mumifiziertes Skelett, bei dem nur unwesentliche Knochen fehlten, wurde 1948 an einem linken Nebenfluss (Reka Mamonta „Fluss des Mammuts“) der Schrenk im Nordosten der Taimyr-Halbinsel entdeckt und im folgenden Jahr geborgen. Weil das Blumenbachs Erstbeschreibung zugrunde liegende Typusmaterial nicht mehr verfügbar war, wurde dieses als Tajmyr-Mammut bekannt gewordene Exemplar seiner guten Überlieferungsqualität und typischen Morphologie wegen zum Neotypus von Mammuthus primigenius erklärt.[16] Im selben Jahr entdeckte man in Alaska in der Nähe von Fairbanks eines der am besten erhaltenen jungen Wollhaarmammuts. Konserviert waren nur Kopf, Vorderbein und Schulter des Tieres, aber insbesondere die Haut und die Muskeln waren sehr gut erhalten. DNA-Untersuchungen an diesem Fund haben wesentlich zur Feststellung der Evolution dieser Tierart beigetragen. Das Exemplar, bekannt unter dem Namen Effie, ist heute im American Museum of Natural History in New York ausgestellt.[5]
Das 1972 am Schandrin, einem Nebenfluss der Indigirka, entdeckte nahezu vollständige Skelett des Schandrin-Mammuts besaß zwar keine gute Erhaltung der äußeren Weichteile, dafür waren die Inneren Organe hervorragend überliefert und erlaubten erstmals einen Einblick in den inneren Aufbau der Mammuts und einen Vergleich zu den rezenten Elefanten. Weiterhin enthielt der Kadaver, der zu einem alten, aber nicht sehr großen Bullen gehörte, 291 kg Nahrungsüberreste. Im Jahr 1977 wurde weiterhin der Kadaver eines jungen weiblichen Tieres am rechten Ufer der Juribei auf der Gydan-Halbinsel gefunden, während im selben Jahr in Schwemmsanden am linken Ufer der Chatanga in Nordsibirien von Rentierhirten ebenfalls ein als Chatanga-Mammut bekannt gewordener Kadaver entdeckt und in einer zweijährigen Grabungskampagne ergraben werden konnte.[4]
Im Kolyma-Becken am Fluss Kirgiljach im Rajon Sussuman in der Oblast Magadan entdeckte ein Arbeiter ebenfalls 1977 bei der Goldgewinnung die Eismumie eines vollständig erhaltenen männlichen Mammutkalbs. Das berühmt gewordene Kirgiljach- oder Magadan-Mammutbaby, das den Namen Dima erhielt, starb vor etwa 39.000 Jahren im Alter von sechs bis acht Monaten.[16] Das Kalb war 115 Zentimeter lang und 104 Zentimeter hoch und wog zum Todeszeitpunkt etwa 100 Kilogramm. An den distalen Enden der Beine waren noch Haarreste erhalten, denn diese Körperpartien frieren nach Eintritt des Todes als erste ein. Die inneren Organe „Dimas“ unterscheiden sich kaum von denen rezenter Elefanten, seine Ohrmuscheln jedoch haben nur ein Zehntel der Fläche eines gleichaltrigen Afrikanischen Elefanten (Loxodonta africana). [17]
Hervorzuheben ist weiterhin das 1988 am Ufer des Juribetejach auf der Jamal-Halbinsel aufgefundene Mammutkalb Mascha, welches in Alter und Größe mit Dima vergleichbar ist, dem aber lediglich der Rüssel fehlt. Ein nur drei Monate altes Kalb mit erhaltenen Schädel samt Haut, kleinen Stoßzähnen, Rüssel und Ohren und Teilen der Gliedmaßen entdeckten Jäger 1990 am Ufer der Indigirka. Es wurde nach der Provinz, aus der es stammt, als Abyi-Mammut bezeichnet.[14] Im Jahr 1997 fanden Angehörige der Dolganen auf der Taimyr-Halbinsel nahe der Großen Balachnija zwei Mammutstoßzähne. Bei Grabungen 1998 und 1999 an der Stelle wurde ein Mammutkadaver mit guter Weichteilkonservierung entdeckt. Dieser wurde daraufhin in einem vollständigen Block geborgen und in eine künstliche Höhle am Chatanga ausgeflogen, um das Mammut dort bei kontrollierten Temperaturen von -15 °C vollständig freizulegen und auch das Bodensediment umfassend zu untersuchen. Der Fund erhielt nach seinen Entdeckern den Namen Jarkow-Mammut.[18][4]
Bereits 1990 war am Ufer der Oberen Taimyra im zentralen Teil der Taimyr-Halbinsel ein Kadaver bekannt geworden, der alsbald wieder überflutet wurde. Zehn Jahre später entdeckte ihn jedoch S. Pankewitsch beim Fischen wieder, weshalb er als Fischhaken-Mammut bezeichnet wird. Der Fund wurde anschließend teilweise ausgegraben, der hintere Teil zusammen mit den inneren Organen und dem Mageninhalt allerdings in einem Block geborgen und zu Studienzwecken in dieselbe künstliche gekühlte Höhle an der Chatanga wie das Jarkow-Mammut ausgeflogen.[4][19][20] Das Jukagir-Mammut wurde 2002 von V. Gorochow am der Maksunuocha im Norden Jakutiens entdeckt und im Sommer 2004 von einem internationalen Team bestehend aus Russen, Jakuten und Japanern ausgegraben. Es zeichnet sich vor allem durch einen exzellent erhaltenen Schädel mit Weichteilbedeckung und beiden vollständig erhaltenen Ohren aus.[21] Einer der jüngsten Funde stammt aus dem Jahr 2007 und wurde von einem Rentierzüchter am Oberlauf des Juribei auf der Jamal-Halbinsel geborgen. Es handelt sich hierbei um ein nur einen Monat altes Kalb, das den Namen Ljuba erhielt und eine hervorragende Erhaltung aufweist.[22]
Außerhalb der Permafrostgebiete stammen konservierte Mammutfunde aus Starunia (Ukraine). Diese wurden 1907 zusammen mit dem mumifizierten Kadaver eines Wollnashorns in einer Ölschiefergrube gefunden, in der sie im mit Salzen durchsetzten Erdwachs in einer Tiefe von 12 bis 17 m unter der Erdoberfläche erhalten geblieben waren. Allerdings war das Fell bereits vergangen.[23]
Aussehen und Lebensweise
Erscheinungsbild

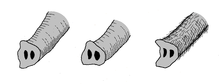 Rüsselenden verschiedener Elefantenarten. Links: Afrikanischer Elefant (Loxodonta afrikana), Mitte: Asiatischer Elefant (Elephas maximus), rechts: Wollhaarmammut (Mammuthus primigenius).
Rüsselenden verschiedener Elefantenarten. Links: Afrikanischer Elefant (Loxodonta afrikana), Mitte: Asiatischer Elefant (Elephas maximus), rechts: Wollhaarmammut (Mammuthus primigenius).Aufgrund der großen Verbreitung des Wollhaarmammuts und der damit verbundenen hohen Fundanzahl, die Knochen, Zähne oder vollständige Skelette umfassen, aber auch der erhaltenen mumifizierten Kadaver oder der Darstellungen in Höhlenmalereien durch den jungpaläolithischen Menschen ist diese Tierart eine der bekanntesten und am besten studierten aus dem Pleistozän. Daher ist das Aussehen und die Lebensweise des Wollhaarmammuts überaus umfassend rekonstruiert.
Wollhaarmammuts waren nicht so riesig wie oft vermutet wird. Ausgewachsene Tiere erreichten eine Widerristhöhe von 2,8 bis 3,75 m, nicht viel mehr als bei heutigen Elefanten. Ein relativ kleines erwachsenes Individuum aus Rottweil (Baden-Württemberg) wies nur eine Höhe von 2,5 m auf.[24] Wie bei den rezenten Elefanten gab es aber einen ausgeprägten Geschlechtsdimorphismus. Männliche Tiere brachten es im Durchschnitt auf 2,7 bis 3,4 m Schulterhöhe, während jene der Kühe oft nur zwischen 2,6 und 2,9 m lag. Die Tiere waren aber kompakter und massiger als Elefanten, so dass das Gewicht zwischen 5 und 6 t, gelegentlich auch bis zu 8 t betrug. Die jüngeren Vertreter der Art waren in der Regel kleiner als die älteren, die in ihrer Körpergröße noch etwas an das mächtige Steppenmammut erinnerten, aus dem die Art hervorging. Dennoch war das Wollhaarmammut im Jungpleistozän das größte Tier der Mammutsteppe. [5][25] Die kleinsten Vertreter der Wollhaarmammuts waren die letzten Angehörigen ihrer Art und lebten auf der Wrangel-Insel. Sie erreichten aufgrund von Inselverzwergung gerade 1,8 m Schulterhöhe bei einem Gewicht von rund 2 t.[26]
Im Gegensatz zu den rezenten Elefanten hatte das Wollhaarmammut kürzere und massige Beine und war insgesamt länger. Der Kopf war sehr hoch mit einem deutlichen Kranialdom, hinter dem eine Halseinbuchtung lag. Auf dem vorderen Rücken trug es einen Buckel, der häufig als Fettpolster gedeutet wird. Die weitere Rückenlinie war markant stark abfallend.
Namensgebend für diese Art ist das grobe Außenfell, das im Winter aus bis zu 90 Zentimeter langen Haaren bestand. Ähnlich wie beim Moschusochsen bildete dieses Fell an Bauch und Flanke eine Schürze. Auch Rüssel, Schwanz und Ohren waren als Kälteschutz mit Fell bedeckt, während die Haare auf dem Kopf einen charakteristischen Pony formten. Unter diesen Deckhaaren befand sich zunächst eine nicht so derbe, flauschige Fellschicht mit 40 bis 50 cm langen Haaren, während die dichte Unterwolle aus 10 bis 20 cm langen Haaren bestand. Das Sommerfell war wesentlich kürzer und weniger dicht als das Winterfell. Die Fellfarbe der Eisleichen vom Wollhaarmammut variiert von rötlich, gelblich über bräunlich bis schwarz. Die häufig rötlichen und relativ hellen Farbtöne gehen dabei meist auf posthume Oxydationsprozesse zurück, während die Naturfarbe eher Richtung Dunkelbraun oder Schwarz tendiert haben dürfte. Durch Forschungen wurde belegt, dass es zudem blonde Wollhaarmammuts gab.[25][27][28]
Die Haut des Wollhaarmammuts erreichte eine Dicke von durchschnittlich 3 cm, an den Fußsohlen war sie jedoch mit 5 bis 6 cm deutlich stärker. Zusätzlich befanden sich zwischen den Zehen noch dicke Hautpolster. Unter der Haut lag eine fast 10 Zentimeter dicke Fettschicht, die für eine zusätzliche Wärmeisolierung sorgte. Da der Tierart aber Talgdrüsen fehlten, mit denen sie ihr Fell hätten einfetten können, war sie feuchten Wetterbedingungen schutzlos ausgeliefert.[25][5]
Der Rüssel des Wollhaarmammuts war als kaltklimatische Anpassung relativ kurz, dafür aber wesentlich dicker und massiver als bei heutigen Elefanten. Markant ist das Rüsselende, das aus einem "Finger" und einem schaufelförmigen Zipfel bestand. Der Afrikanische Elefant hingegen weist an seinem Rüsselende zwei "Finger" auf, während der Indische Elefant (Elephas maximus) nur einen hat. Die Finger dienten weitgehend dem Greifen der Nahrung und funktionierten dabei ähnlich wie die menschliche Hand.[10] Ebenfalls eine Klimaadaptation stellen die sehr kleinen Ohren dar. Sie hatten eine ovale Form mit einer Länge von 38 cm und einer Breite von 18 bis 28 cm. Dadurch erreichten die Ohren ausgewachsener Tiere nur etwa ein Zwanzigstel der seiner heutigen afrikanischen Verwandten,[21] bei Jungtieren wie Dima immerhin noch ein Zehntel verglichen mit gleichaltrigen Elefantenkälbern. Auch der Schwanz war mit rund 40 cm relativ kurz und besaß am Ansatz einen Hautlappen, der als Analklappe diente. Das Schwanzende trug eine ausgeprägte Quaste aus bis zu 60 cm langen Haaren.[25][10]
Schädel-, Gebiss- und postcraniale Skelettmerkmale
 Das Mammut „Millie“ im „Celtic and Prehistoric Museum“, Ventry, Irland
Das Mammut „Millie“ im „Celtic and Prehistoric Museum“, Ventry, IrlandDer Schädel war groß und sehr hoch mit einem teils ausgeprägten quer verlaufenden Scheitelkamm und einem massiven Unterkiefer. Der Scheitelkamm diente als Ansatzstellen einerseits für die benötigte starke Rückenmuskulatur, die den Schädel mit den Stoßzähnen halten musste, andererseits auch für den mächtigen Kauapparat. Um das Gewicht des Schädels gering zu halten, bestand das Innere, wie bei allen Elefanten, aus einer wabenartigen Struktur mit dünnen Knochenlammellen.[10]
Charakteristisch sind die Alveolen der Stoßzähne, die beim Wollhaarmammut fast parallel oder in einem geringen Winkelabstand zueinander verliefen. Dies unterscheidet ihn z. B. vom ebenfalls mittel- und jungpleistozänen Europäischen Waldelefanten, wo diese sich in einem stärkeren bis zu 80° messenden Winkel zueinander befanden.[29]
Das Wollhaarmammut hatte wie alle echten Elefanten ein stark reduziertes Gebiss, das nur die Backenzähne und die aus den oberen Schneidezähnen gebildeten Stoßzähne umfasste. Pro Kiefernbogen besaß das Wollhaarmammut einen Backenzahn, der beginnend mit dem ersten Milchmolar fünfmal ausgetauscht werden konnte (also drei Milchmolaren und drei Dauermolaren). Die Backenzähne waren langoval, oft über 20 cm lang und erreichten von allen Elefanten die höchsten Zahnkronen. Auffallend ist die hohe Anzahl von Schmelzlamellen, die beim dritten Molaren zwischen 21 und 30 betrug, mit einer durchschnittlichen Schmelzfaltendicke von 0,9 bis 2 mm. Dies ist die höchste Anzahl bei allen bekannten Vertretern der Elefanten. Teilweise dient dieser Wert als taxonomisches Unterscheidungsmerkmal, wobei hohe Abkauungsgrade eine genaue Artbestimmung beeinträchtigen können.[1]
Das postcraniale Skelett des Wollhaarmammuts besitzt nur wenige Unterschiede zu denen anderer Elefanten. Charakteristisch sind die säulenförmigen Gliedmaßen. Auffällig ist auch der kurze Schwanz, der aus nur 21 Wirbeln gebildet wird, während der der rezenten Rüsseltiere zwischen 28 und 33 umfasst.[5] Lange Zeit hegten Paläontologen die Hoffnung, einzelne oder isolierte Skelettelemente den verschiedenen Elefantenarten zuweisen zu können, was sich jedoch nicht erfüllt hat. Lediglich am ersten Halswirbel, dem Atlas, zeigt das Wollhaarmammut im Gegensatz zum Europäischen Waldelefanten ein niedrigeres und breiteres craniales Gelenk.[30]
Stoßzähne
 Querschnitt eines Mammutstoßzahns. Deutlich sind die ringförmigen Lamellen und die Schreger-Linien, vor allem in der zweiten und dritten äußeren Lamelle, zu erkennen.
Querschnitt eines Mammutstoßzahns. Deutlich sind die ringförmigen Lamellen und die Schreger-Linien, vor allem in der zweiten und dritten äußeren Lamelle, zu erkennen.Die Stoßzähne gehören zu den markantesten äußeren Merkmalen des Wollhaarmammuts und sind im Gegensatz zu den heute noch lebenden Elefantenarten deutlich spiralförmig nach oben gedreht, wobei die spitzen Enden meist aufeinander gerichtet sind. Durch die Drehung der Defensen sind diese seitlich sehr weit ausladend mit einer Spanne bis zu 1,7 m.[21] Stoßzähne ausgewachsener alter Bullen konnten eine Länge, von der etwa ein Viertel in den Alveolen steckte, von mehr als 4,50 m bei einem Gewicht von mehr als 100 kg erreichen. Über die Krümmung gemessen ist das größte bisher bekannte Exemplar 4,9 m lang. Im Durchschnitt sind die aufgefundenen Stoßzähne mit 2,50 m Länge und 45 kg Gewicht allerdings deutlich kleiner. An der Austrittsstelle aus den Alveolen wiesen sie einen Durchmesser vom bis zu 20 cm auf. Kühe hatten kürzere und dünnere Stoßzähne - der Durchmesser an den Alveolen liegt im Durchschnitt bei 9 bis 10 cm -, die nicht so deutlich spiralförmig sondern eher säbelartig gebogen waren. Im Allgemeinen waren die Defensen des Wollhaarmammuts länger als jene der heute lebenden Elefanten.[8][25]
Die Stoßzähne bestehen aus Elfenbein, einem Gemisch aus Carbonat-Hydroxyl-Apatit-Kristallen, die mit Kollagenfasern orientiert verwachsen sind, wobei die Kristalle für die notwendige Härte, das Kollagen aber für die Elastizität verantwortlich sind. Aufgrund von differierenden Kristallisationsgraden im Elfenbein der verschiedenen Elefantenarten lassen sich die Stoßzähne in Dünnschliffproben mit Hilfe der Infrarotspektroskopie unterscheiden. Ebenso zeigt das Kollagen in den Stoßzähnen der einzelnen Elefantenarten Abweichungen, so das unter Verwendung der Ultraviolett-Fluoreszenz-Spektrophotometrie diese zugewiesen werden können.[31]
Im inneren Aufbau weisen die Stoßzähne drei unterschiedliche Lagen auf. Die äußerste dünne Lage besteht aus Zement, darunter folgt das Dentin als Hauptbestandteil. Dieses besitzt eine faserige Struktur und ist von unzählige Kanälchen durchsetzt, die mit Kollagen gefüllt sind. Den innersten Bereich bildet die Pulpa, in der sich Blutgefäße und Nerven befinden. Auch gehen von ihr zahlreichen Dentinkanälchen nach außen aus. Stoßzähne haben im Gegensatz zu anderen Zähnen keinen Zahnschmelz. Neues Stoßzahnmaterial wird hauptsächlich in den Alveolen gebildet und erfolgt von Innen nach Außen. Dadurch ist der Stoßzahn im Querschnitt aus konzentrischen Ringen aufgebaut, die jeweilige Wachstumsschübe darstellen und den Jahresringen in den Stämmen der Bäume vergleichbar sind. Die innersten Ringe sind dabei am jüngsten. Im Längsschnitt hingegen zeigen sich parallel verlaufende abwechselnd helle und dunkle Linien, die ebenfalls Zuwachsraten anzeigen.[31][10]
Weiterhin können im Querschnitt Strukturen beobachtet werden, die über die Wachstumsringe hinausgehen und eine rosettenartige Maserung aus abwechselnd hellen und dunklen Linien bilden. Diese als Schreger-Linien bezeichneten Bildungen gehen auf einen regelmäßigen Wechsel des Kollagengehaltes im Dentin zurück. Die Winkel, mit denen sich die Linien regelmäßig treffen sind aufgrund der starken Krümmung des Mammutstoßzahns spitzer als bei den rezenten Elefanten und variieren zwischen 25 bis 40° je nach Lage im Stoßzahn.[32][10]
Die Funktion der Stoßzähne war sicher vielfältig. Zum Einen dienten sie zur Darstellung der Dominanz einzelner Tiere im Paarungs- oder Ritualkampf. Hierbei ist aber ein Aufspießen wie bei den rezenten Elefantenarten nicht möglich gewesen. Weiterhin könnten sie ein Warnsignal an Nahrungskonkurrenten im interspezifischen Wettbewerb oder aber auch an große Raubtiere gewesen sein. Sehr häufig wurden die Stoßzähne aber für die Nahrungssuche eingesetzt. Dies zeigen Schliffspuren, die sich überwiegend an der Unterkante, gelegentlich aber auch auf der Oberseite und der Spitze befinden.[33] Meist werden diese Schliffspuren mit dem Freischaufeln des mit Schnee bedeckten Erdbodens bei der Suche nach Nahrung erklärt, doch könnten sie auch allgemein beim Herauswühlen von Pflanzen oder Umbiegen von Sträuchern entstanden sein. Solche Spuren sind auch bei rezenten Elefanten häufig zu finden.[5]
Ernährungsweise
Das Wollhaarmammut war ein an Steppengebiete angepasstes Tier, wie der gedrungene Körperbau und vor allem die kräftigen, auf lange Wanderungen spezialisierten Gliedmaßen zeigen. Da in Steppen hauptsächlich Gräser als Nahrungsquelle zur Verfügung standen, wurde forschungsgeschichtlich schon sehr früh vermutet, dass das Rüsseltier ein ausgewiesener Grasfresser war. Dafür gibt es auch anatomische Merkmale, wie die sehr hochkronigen Backenzähne mit ihren zahlreichen Schmelzfalten. Da Gräser nicht sehr nährstoffhaltig sind, musste das Wollhaarmammut sehr große Mengen verzehren. Die hohen Zahnkronen glichen dabei die durch die hohe Nahrungsmenge verursachte größere Abkauung wieder aus. Da Gräser zusätzlich noch kieselsäurehaltig und damit sehr hart sind, verhinderte die hohe Schmelzfaltendicke ebenfalls einen stärkeren Abrieb.[25]
Einige der im sibirischen Permafrost entdeckten Mammutkadaver enthielten noch Mageninhalte, wie z. B. das Schandrin-Mammut oder das Berjosowka-Mammut, letzteres wies sogar noch Nahrungsreste auf der Zunge bzw. im Maul auf. Die Untersuchungen dieser Nahrungsreste bestätigten die anatomischen Hinweise. So überwiegen vor allem Gräser, wie Fuchsschwanzgräser, Gerste, Straußgräser, Salzschwaden und Seggen, darüber hinaus sind Hahnenfuß und Moose nachgewiesen. In geringerem Umfang kamen auch Reste von Weiden- und Lärchenzweigen vor, ebenso wie Teile von Weiden, Erlen und Kiefern. Das Vorkommen von Gehölzpflanzen in der Nahrung des Wollhaarmammuts zeigt, dass die Steppe nicht vollständig strauch- und baumfrei war. Die Aufnahme solcher Pflanzenarten war überdies wichtig, um an Nährstoffe zu gelangen, die für das Wachstum benötigt wurden. Die große Vielfalt an verzehrten Pflanzen deckte gleichzeitig auch den Aminosäurebedarf, den das Wollhaarmammut im Gegensatz zu den großen Wiederkäuern vollständig über die Nahrungsaufnahme regeln musste.[25][5]
Ein einzelnes Tier benötigte je nach Jahreszeit 150 bis 300 kg Nahrung täglich, wobei die Menge der aufgenommenen Pflanzen abhängig vom Trocknungsgrad war, d. h. im Frühjahr mit frisch gewachsenem Gras brauchte ein Tier weniger als im Sommer und Herbst mit trockenem Pflanzenwuchs. Auf den allgemein hohen Nahrungsbedarf weist neben Vergleichen mit rezenten Elefanten auch der Mageninhalt des Schandrin-Mammuts hin, der mehr als 290 kg umfasste. Der Abbau der pflanzlichen Zellulose erfolgte beim Wollhaarmammut im Blinddarm. Dies ermöglichte ihm bei sinkendem Eiweiß- und steigendem Fasergehalt größere Mengen zu fressen und so den Großteil des Tages mit der Nahrungsaufnahme zu verbringen.[25][8]
Neben der pflanzlichen Nahrung war das Wollhhharmammut stark von Wasser abhängig. Im Durchschnitt verbrauchte es rund 70 bis 90 l am Tag. Die benötigte Wassermenge war wiederum abhängig von der Beschaffenheit der Pflanzennahrung. Bei saftigem frischen Gras kam das Wollhaarmammut mit weniger Wasser aus. Das erlaubte ihn auch sich weiter von den Wasserstellen wie Fluss- und Seeufer in die umliegenden Gebiete zu entfernen. Im Sommer und Herbst, wenn die meisten Pflanzen ausgetrocknet waren, war der Wasserbedarf entsprechend höher. Es ist möglich, dass sich in diesen Jahreszeiten der Aufenthalt der Wollhaarmammuts an den Wasserstellen konzentrierte, wodurch die Wanderungsbewegung der Tierart stark saisonabhängig war.[8]
Individuelle Lebensweise
 Rekonstruktion des Wollhaarmammuts und seiner Umwelt
Rekonstruktion des Wollhaarmammuts und seiner UmweltWollhaarmammuts erreichten ein Alter von 60 bis 65 Jahren. Dies geht zum Einen aus der Größe der Tiere hervor, zum Anderen auf den charakteristischen, allen Elefantenarten eigenen "horizontalen" Zahnwechsel zurück. Der letzte Molar, der dritte, ist ungefähr im Alter von 60 Jahren abgekaut und fällt aus. Alte Tiere können dann nicht mehr genügend Nahrung aufnehmen und sterben schließlich.
Man geht davon aus, dass Wollhaarmammuts, ähnlich wie die heutigen Elefantenarten, in matriarchalisch geprägten Familienverbänden zusammen lebten. Das älteste Muttertier leitete dabei die Gruppe, die normalerweise aus zwei bis maximal 20 Tieren bestand. Bullen dagegen führten ein weitgehendes einzelgängerisches Leben oder formierten sich bei Bedarf in "Junggesellengruppen", mit den Kühen kamen sie nur während der Brunft zusammen. Diese war vermutlich aufgrund der starken jahreszeitlichen Temperaturschwankungen saisonabhängig und fand im Sommer statt, im Gegensatz zu den heutigen an das tropische Klima angepassten Elefanten mit einer ganzjährigen Paarungsbereitschaft. Nach 22monatiger Tragzeit wurde das Jungtier dann im Frühjahr geboren, wobei dieses wohl rund 90 kg wog. Junge Bullen wurden dann mit 9 bis 12 Jahren von der Herde vertrieben.[34][8] Wie bei den rezenten Elefanten ist auch beim Wollhaarmammut die subcutan am Kopf zwischen Auge und Ohr liegende Musth-Drüse nachgewiesen, die im jährlichen Zyklus Sekrete ausschied, welches eine erhöhte Aggressivität im Dominanz- und Paarungsverhalten verursachte.[35]
Ausgewachsene Wollhaarmammuts hatten keine natürlichen Feinde, Jungtiere konnten hingegen von großen Raubtieren wie dem Höhlenlöwen von der Herde getrennt und anschließend gerissen werden. Einzig der frühe Mensch war als aktiver Jäger mit Distanzwaffen dem Wollhaarmammut gefährlich. Darüber hinaus gab es aber verschiedenste Gefahren, die auf das Leben des Wollhaarmammuts einwirkten. Rutschige Hänge, hochwasserführende Flüsse im Frühjahr oder einbrechende Thermokarste konnten zu schweren Stürzen bis hin zum Tod einzelner Individuen. Gelegentlich zeugen einzelne gebrochene Knochen, wie z. B. am Schulterblatt von solchen Ereignissen. Möglicherweise sind ein Großteil der Eismumien auf solche zufälligen Ereignisse zurückzuführen.[5]
Wie bei den rezenten Elefanten ist zu vermuten, dass das Wollhaarmammut sehr emotional auf den Tod vom Artgenossen, vornehmlich Herdenmitgliedern, reagierte und oftmals bis zu mehreren Stunden am Sterbeplatz verweilte. Da die zurück gebliebenen Tiere später diese Orte meist mieden, kann dieses Verhalten auch Einfluss auf die Jagdstrategien und Sammeltätigkeiten der frühen Menschen Auswirkung gehabt haben.[8]
Des Weiteren können auch einzelne Krankheitserscheinungen beobachtet werden. Relativ häufig treten Backen- und Stoßzahnanomalien in Form von Wachstumsstörungen auf, ebenso wie Parodontitis und teilweise auch Karies. Bemerkenswert ist auch das vereinzelte Vorkommen überzähliger Backenzähne bei älteren Individuen. Gelegentlich wurden auch Krebsgeschwüre an Zähnen festgestellt.[36][37] An Knochenerkrankungen sind vor allem Arthritis an den Wirbeln bis hin zum Verwachsen ganzer Abschnitte der Wirbelsäule beobachtet worden. Auch konnte an einigen Funden Osteomyelitis nachgewiesen werden.[5][4]
Der frühe Mensch und das Wollhaarmammut
Das Wollhaarmammut als Nahrungs- und Rohstofflieferant
Wie bei vielen pleistozänen Großsäugetieren ist eine aktive Bejagung durch den frühen Menschen schwer nachweisbar, da die meist aus organischem Material bestehenden Jagdwaffen äußerst selten aufgefunden werden. Prinzipiell ließen sich alle Überreste eines Wollhaarmammuts verwerten, so natürlich Fleisch, Fett und Knochenmark als Nahrung, Knochen und Elfenbein als Rohmaterial für Geräte, Werkzeuge und Schmuck, das Fell als Kleidung oder Bedeckung für Wohnbauten wie Zelte und Sehnen für Fäden und Schnüre.[38]
Frühe Zusammenfunde von menschlichen Hinterlassenschaften und Wollhaarmammuts stammen aus dem Mittelpaläolithikum (vor 300.000 bis 40.000 Jahren) der ersten Hälfte der Weichsel-Kaltzeit. An der rund 60.000 Jahre alten Fundstelle Salzgitter-Lebenstedt (Niedersachsen) sind neben anderen Tierresten Knochen und Zähne von mindestens 16 Mammutindividuen zusammen mit rund 1.000 Feuersteinartfakten nachgewiesen. Ob die Mammuts aber von den damaligen Neandertalern auch erlegt wurden, ist nicht bekannt. Da hier das Ren mit etwa 80 Individuen dominiert, handelt es sich wohl eher um eine Gruppe spezialisierter Rentierjäger.[39] Auch in der rund 90.000 Jahre alten Fundschicht A von Königsaue (Sachsen-Anhalt) im Nordharzvorland wurden Reste von mindesten vier jüngeren Mammutindividuen gefunden, die mit Geräten des Micoquien vergesellschaftet sind.[40] Eine mögliche direkte Verwertung eines Wollhaarmammuts vor Ort oder gar eine aktive Bejagung könnte der Befund von Asolo (Italien) darstellen, der mehr als 50 Knochenreste eines erwachsenen weiblichen Mammuts in direkter Verbindung mit fünf Feuersteinartefakten, darunter zwei Levalloisspitzen, umfasst.[41] Auch in Sibirien sind mehrere Fundstellen aus dem Mittelpaläolithikum bekannt, an denen Mammutknochen mit menschlichen Hinterlassenschaften assoziiert sind, so im nördlichen Teil allein acht[42] und im südlichen Teil am Oberlauf des Jenissei wenigstens zwei.[43] Die Nutzung von Mammutknochen als Rohmaterial zeigt der bekannte beinerne rund 15 cm lange Faustkeil aus Rhede (Nordrhein-Westfalen)[44] oder einzelne bearbeitete Knochen aus der Kůlna-Höhle in Mähren.[45] Ein unikater Fund stammt aus Tata (Ungarn), wo eine Lamelle eines unteren Mammutmolars von Neandertalern vollständig poliert, die Kanten gerundet und die Oberflächen mit Ocker bedeckt wurden. Da dieses Objekt zwar überarbeitet, aber nicht benutzbar war, gehört es zu den seltenen Funden von non-utilitarian objects (nicht genutzte Objekte), die schon beim Neandertaler ein mögliches frühes Aufkeimen von künstlerischem Ausdruck aufzeigen.[46][47]
Im darauf folgenden Jungpaläolithikum (vor 40.000 bis 11.600 Jahren), dessen Träger der anatomisch moderne Mensch war, war das Wollhaarmammut ebenfalls eine wichtige Rohmaterialressource. Aber auch hier gibt es trotz des riesigen Verbreitungsgebietes nur ganz wenige Hinweise auf eine aktive Jagd auf dieses Großsäugetier.[43] Aus den aurignacienzeitlichen Fundschichten der Vogelherdhöhle (Baden-Württemberg) sind mehr als 3.500 Reste von wenigstens 28 Mammutindividuen bekannt. Dabei wurde sicher nicht nur das Fleisch verzehrt sondern auch Knochen und Elfenbein verarbeitet, wie Funde von Elfenbeinstäben zeigen, die - ausgehend von der natürlichen Krümmung der Stoßzähne - gerade gebogen wurden.[48][49] Ein ähnlicher etwa zeitgleicher Befund stammt aus aus der Istállóskö-Höhle im Bükk-Gebirge (Ungarn).[46] Während des nachfolgenden Gravettien stieg die Nutzung des Wollhaarmammuts als Ressource im südlichen und südöstlichen Mitteleuropa (hier auch Pavlovien genannt) deutlich an. An der Station Krems-Wachtberg (Niederösterreich) ist die Tierart die dominante Spezies. Vor allem an Wirbeln und Rippen zeigen sich deutliche Schnittspuren, die offensichtlich bei der Entfernung des Fleisches entstanden, einzelne Rippen sind auch zu Geräten weiterverarbeitet worden.[50] Hervorzuheben sind die Knochenanhäufungen von Wollhaarmammuts an der ebenfalls dem Gravettien angehörenden Fundstelle Dolní Věstonice (Mähren), wo auf einer Fläche von etwa 12 mal 45 m über 6.300 Knochen- und Zahnreste von mehr als 156 Mammutindividuen gefunden wurden. Ähnliche Befunde gibt es auch von Predmosti und Milovice (beide ebenfalls Mähren).[51][52] Besonders bedeutend war die Nutzung der Knochen und Stoßzähne des Wollhaarmammuts im östlichen Europa. Allein in Mezin (Ukraine) wurden Reste von mehr als 108 Mammutindividuen gefunden, während es in Mezhirich (ebenfalls Ukraine) 110 waren. Beide Fundstellen gehören dem osteuropäischen Epigravettien (entspricht dem Magdalénien Mitteleuropas) an und datieren um 15.000 BP. Spektakulär sind die Mammutknochenhäuser von beiden Fundstellen, aber auch von Dobranichevka und Kiev, Kirillovskaja Ulica (alle Ukraine). Allein in Mezin sind fünf Hüttenreste überliefert, der am besten erhaltene hatte einen Durchmesser von 5 m. An seiner Peripherie befanden sich 14 Schädel des Rüsseltiers nebst Lang- und Beckenknochen, während im Innern vor allem Schulterblätter und Unterkiefer lagen. Diese bildeten offensichtlich die Wandung der Rundhütte, welche später einstürzte.[53][38] Auch im östlichen Eurasien hatte das Wollhaarmammut als Rohstofflieferant Bedeutung, wenn auch nicht so markant wie in Osteuropa. So kommen im nördlichen Sibirien wenigstens an zehn Fundstellen Wollhaarmammut und Mensch gemeinsam vor.[43]
Nach dem letztkaltzeitlichen maximalen Eisvorstoß vor 20.000 bis 16.000 Jahren und im darauffolgendem Magdalénien tritt das Wollhaarmammut in West- und Mitteleuropa nur noch selten in Erscheinung. So sind z. B. an dem bedeutenden Lagerplatz von Gönnersdorf (Rheinland-Pfalz) Reste eines einzigen Individuums überliefert.[38] Auch im östlichen Eurasien, wo das Wollhaarmammut zu jener Zeit noch häufiger vorkam, geht die Bedeutung als Rohstoffquelle allmählich zurück. So sind Mammutreste in der Spätphase der Weichselkaltzeit in der Jenissei-Region nur an jeder dritten archäologischen Fundstation vertreten, während es in der vorhergehenden Zeit noch an fast zwei Drittel aller menschlichen Siedlungsplätze nachweisbar ist.[43] Aus anderen Regionen Sibiriens sind relativ wenige Funde bekannt. Bedeutend ist hier ein Siedlungsplatz menschlicher Jäger-Sammler-Gruppen am Ufer des Bjorjoljochs nur etwa 100 m von dem berühmten und zeitgleichen Mammutfriedhof entfernt, was annehmen lässt, das die Mitglieder dieser Gruppen diesen Friedhof als Rohstoffquelle nutzten.[42]
Das Wollhaarmammut in der jungpaläolithischen Kunst
 Mammutdarstellung aus der Höhle von Les Combarelles (Frankreich).
Mammutdarstellung aus der Höhle von Les Combarelles (Frankreich).Dass das Wollhaarmammut eines der beeindruckendsten Tiere der Mammutsteppe darstellte, zeigen auch die Höhlenmalereien des Jungpaläolithikums (seit etwa 32.000 BP, z. B. Grotte Chauvet, Ardeche). Auch in der mobilen Kunst gibt es Mammut-Darstellungen, zum Beispiel aus der Vogelherdhöhle und dem Geißenklösterle in Baden-Württemberg. So gewähren uns die frühen Jäger und Sammler des westlichen Eurasiens Einblicke in ihre Umwelt und helfen neben den biologischen und paläontologischen Analysen bei der Rekonstruktion dieses Lebewesens.
Die ältesten Mammut-Darstellungen der Frankokantabrischen Höhlenkunst aus der Grotte Chauvet (Frankreich) werden dem Aurignacien zugewiesen. Mindestens 34 Abbildungen, abwechselnd in roten oder schwarzen Pigmenten gehalten oder in den Fels eingraviert, sind überliefert und stellen die dritthäufigste Tierart nach dem Wollnashorn und dem Höhlenlöwen dar. Charakteristisch ist die fast hufeisenförmig gestaltete Linie, die den Bauch und die Beininnenseiten wiedergibt.[54] Besonders häufig ist das Wollhaarmammut aber in der südfranzösischen Höhle von Rouffignac dargestellt, deren Abbildungen der Kulturstufe des Magdalénien angehören und wo es mindestens 150 Zeichnungen dieses Rüsseltiers gibt. Die Wollhaarmammute sind teilweise sehr realistisch wiedergegeben, mit hohen Schädelbuckeln, schräg abfallender Rückenlinie und stark gekrümmten Stoßzähnen. Für die Anerkennung der Authentizität der Höhlenkunst von Rouffignac bedeutend war vor allem die Tatsache, dass die Darstellung der Afterklappe als anatomisches Detail erst nach der Entdeckung der Zeichnungen auch in der Paläozoologie wiederentdeckt worden ist.[55] Bezüglich anatomischer Details hervorzuheben sind der als Patriarch bezeichnete Bulle mit markanten Stoßzähnen sowie zwei Mammuts, die sich Stirn an Stirn gegenüberstehen und einen Ritualkampf oder eine Begrüßungszeremonie zu zelebrieren scheinen.[56] Weitere häufige Darstellungen des Wollhaarmammuts finden sich in Font-de-Gaume und Pech Merle mit je 23 bzw. in Les Combarelles mit 14 Abbildungen,[57] weswegen diese auch als "Mammutheiligtümer" bezeichnet werden. Insgesamt umfasst das Wollhaarmammut dabei 6 bis 7 % aller Tier- und Menschendarstellungen in den Höhlen Frankokantabriens.[56] Außerhalb dieses Kulturkreises sind vor allem die roten Darstellungen der Tierart aus der Höhle von Kapova im Ural bekannt.[58]

Figürliche Abbildungen des Wollhaarmammuts finden sich auch in der jungpaläolithischen Kleinkunst. Bedeutend sind die mehr als 60 Darstellungen des Rüsseltiers aus der magdalenienzeitlichen Siedlung von Gönnersdorf, die in Schieferplatten eingeritzt sind. Hier können zwei unterschiedliche Gruppen herausgestellt werden: Tiere mit einem Sattel hinter dem Schädelbuckel und abfallender Rückenlinie und solche mit konvexem Rückenverlauf, deren höchster Punkt etwa in der Mitte liegt. Während erstere als ausgewachsene Tiere gedeutet werden, sollen zweitere Jungtiere darstellen. Auch von der "Teufelsbrücke" bei Saalfeld (Thüringen) stammt ein Tonschiefergeröll mit einer allerdings nur den Kopf und Rüssel zeigenden Mammutdarstellung.[59][58][60] Weitere Abbildungen dieses Rüsseltiers wurden in La Madeleine (Frankreich), eine Fundstelle, die zur Definition des Magdaléniens beitrug, gefunden. Das hier mit erhobenen Schwanz erregt erscheinende Tier ist mit zahlreichen Details, wie dem charakteristischem Pony oder der Kehlbehaarung ausgefürt. Auch aus Malta nahe des Baikalsees (Russland) sind Darstellungen auf Knochenplättchen bekannt.[57]
Darüber hinaus gibt es auch Voll- und Halbplastiken, die das Wollhaarmammut wiedergeben. Zu den bekanntesten gehören jene aus der aurignacienzeitlichen Besiedlungsphase der Vogelherdhöhle und dem Geißenklösterle (beide Baden-Württemberg), eine etwas ältere Darstellung stammt aus Sungir (Russland). Aus dem folgenden Gravettien sind die Mammutfiguren aus Pavlov und Dolní Věstonice (beide Tschechien) hervorzuheben, die aus gebranntem Ton bestehen und zu den ältesten Keramikfunden der Welt gehören. Eher funktional waren Speerschleudern in Form von Mammutdarstellungen, wie sie aus Bruniquel oder Canecaude (beide Frankreich) überliefert sind und dem Magdalénien angehören.[58]
Ursprünglich waren aus Nordamerika keine Darstellungen des Mammuts bekannt. Kürzlich wurde von einem solchen Fund aber berichtet, der ein deutlich wiedergegebenes Rüsseltier mit hohem Kopfbuckel, abfallender Rückenlinie und gedrehten Stoßzähnen zeigt. Die nur 7,5 cm lange Darstellung ist in den Langknochen eines großen Säugetiers (Mammut, Mastoden oder Riesenfaultier) eingeritzt und weist ein Alter von ca. 13.000 Jahren auf. Da der Fund aus Florida (Hauptkanal von Vero Beach) stammt, wo das Wollhaarmammut selbst nicht nachgewiesen ist, handelt es sich offensichtlich um die Abbildung eines Präriemammuts, das hier vorgekommen ist.[61]
 Die Dame von Brassempouy (Frankreich) aus dem Gravettien.
Die Dame von Brassempouy (Frankreich) aus dem Gravettien.Des weiteren dienten Knochen und Stoßzähne auch als Rohmaterial für die jungpaläolithische Kunst und einige der bedeutendsten Kunsterzeugnisse jener Zeit sind aus diesen organischen Materialien hergestellt. So bestehen einige der bereits erwähnten Mammut-Figuralplastiken aus Elfenbein, wie jene aus Vogelherdhöhle, aber auch andere Tierfiguren, u. a. von einem Pferd, einer großen Raubkatze, einem Bären oder einem Wisents aus derselben Höhle. Von herausragender Bedeutung ist auch der aurignacienzeitliche Löwenmensch aus dem Hohlenstein-Stadel (Baden Württemberg), die Adorantenfigur aus dem Geisenklösterle, ebenso wie die Kopfdarstellungen der Dame von Brassempouy (Frankreich) und ein ähnlich geartetes Figürchen aus Dolní Věstonice. Weiterhin sind bemalte Knochen aus Mezin (Ukraine) oder solche mit komplexen Ritzverzierungen in Stoßzähnen aus Předmostí (Tschechien) überliefert.[58][57]
Auch Musikinstrumente wurden teilweise aus Elfenbein hergestellt, wie die Funde von Flötenfragmenten aus der Vogelherdhöhle, Geisenklösterle und dem Hohlen Fels, alle im Tal der Ach bei Blaubeuren gelegen, beweisen.[62] Schlussendlich fanden Schmuck- und Kunstgegenstände auch Verwendung im jungpaläolithischen Bestattungsritus. Herausragend sind hier der Grabfund eines Mannes und die Doppelbestattung zweier Kinder aus der früh- bis mitteljungpaläolithischen Station Sungir. Neben durchbohrten Elfenbeinscheiben wurden vor allem bei dem Männergrab über 3.500 Elfenbeinperlen verteilt über den Körper des Toten gefunden, die es ermöglichten, die Bekleidung genau zu rekonstruieren, während den beiden Jugendlichen je eine rund 2,4 m lange Elfenbeinlanze beigegeben wurde.[58] Daneben wurden auch vollständige Knochen vom Wollhaarmammut in den Gräbern beigelegt. Im Gravettien dienten so u. a. Schulterblätter zur Abdeckung der Bestattungen. Beispiele hierfür sind die Gräber von Dolni Vestonice, Pavlov oder Předmostí,[63] aber auch das erst vor wenigen Jahren entdeckte Kinder-Doppelgrab von Krems-Wachtberg.[64]
Stammesgeschichte
Ursprung und Entwicklung
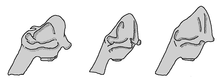 Schematische Darstellung der Änderung der Schädelmorphologie bei verschiedenen Mammutarten. Links: Südelefant (Mammuthus meridionalis), Mitte: Steppenmammut (Mammuthus trogontheriie), rechts: Wollhaarmammut (Mammuthus primigenius).
Schematische Darstellung der Änderung der Schädelmorphologie bei verschiedenen Mammutarten. Links: Südelefant (Mammuthus meridionalis), Mitte: Steppenmammut (Mammuthus trogontheriie), rechts: Wollhaarmammut (Mammuthus primigenius).Die Gattung Mammuthus aus Afrika, wo sie als Mammuthus subplanifrons schon im frühen Pliozän und als Mammuthus africanavus auch im Pleistozän nachgwiesen ist. Aus Mammuthus subplanifrons entwickelte sich dann der Südelefant (Mammuthus meridionalis), welcher vor knapp 3 Mio. Jahren auch als einer der ersten Vertreter der Elefanten eurasischen Boden betrat.[65] Von diesem spaltete sich dann vor rund 1,5 Mio. Jahren das Steppenmammut ab. Mit den einzelnen evolutiven Stufen gingen skelettanatomische Veränderung einher, die u. a. die Verlängerung des Hinterhauptes und damit die Ausbildung eines hohen Craniums, aber auch die allmähliche Krümmung der Stoßzähne und Änderung der Backenzahnstruktur umfassen. Letztere zeigen eine deutliche Zunahme der Schmelzlamellen bei gleichzeitiger Abnahme der Dicke des Zahnschmelzes. Hatte der Südelefant nur 13 bis 18 Schmelzlamellen mit einer durchschnittlichen Dicke von 2,0 bis 3,9 mm, so besaß sein phylogenetischer Nachfolger das Steppenmammut bereits zwischen 17 und 23 mit einer Dicke von 1,0 bis 3,5 mm. Die Zunahme der Schmelzfalten ist ein Anzeichen für eine stärkere Anpassung an offene Landschaftsverhältnisse und eine steigende Spezialisierung auf die daraus resultierende Grasnahrung.[66][67]
Das Steppenmammut hatte sein größtes Ausdehnungsgebiet im späten Altpleistozän vor rund 800.000 Jahren erreicht und kam damals von Ostasien bis nach Westeuropa vor. Mit einer Schulterhöhe von bis zu 4,5 m und einem Gewicht von 10 t gehörte es zu den größten Vertretern der Elefanten überhaupt. Aus dem Steppenmammut entwickelte sich letztendlich das Wollhaarmammut mit seinen charakteristischen Eigenschaften. Der genaue Prozess ist aber noch nicht abschließend geklärt. Der Ausgangspunkt für die Entwicklung des Wollhaarmammuts liegt wahrscheinlich im östlichen Eurasien. In Nordostsibirien, im Olyor-Faunenkomplex, treten vor 800.000 bis 600.000 Jahren die ersten fortgeschrittenen Mammuts auf, die sich durch eine höhere Lamellenzahl (22 bis 24) in den Backenzähnen und eine größere Zahnhöhe auszeichnen. Der Prozess ist dann vor 200.000 Jahren mit dem Auftreten des typischen Wollhaarmammuts abgeschlossen. Damit scheint diese Region eines der Zentren der Entstehung des Wollhaarmammuts zu sein.[68][66][1]
Im westlichen Eurasien unterzieht sich das Steppenmammut in derselben Zeit einer deutlichen Reduktion der Körpergröße. Sind in Süßenborn (Thüringen) und Mosbach vor 500.000 bis 600,000 Jahren noch relativ große Individuen nachgewiesen, so besitzen sie im späten Mittelpleistozän vor 200.000 Jahren schon eine wesentlich geringere Körpergröße, wie Funde von Ilford (England) und Ehringsdorf (Thüringen) zeigen. Einhergehend ist damit auch eine leichte Zunahme der Lamellenanzahl zu verzeichnen, die aber noch nicht die typische hohe Anzahl wie bei den späteren Wollhaarmammuts erreicht und bei durchschnittlich 19 Lamellen liegt. Auch erreichen die Zahnkronen noch nicht die Werte des späteren Wollhaarmammuts.[68][66]
Die dergestaltige morphologische Änderung des Steppenmammuts ließ viele Forscher in westlichen Eurasien vermuten, es handle sich hierbei um eine Misch- oder Übergangsform zum Wollhaarmammut. Deshalb wurde für die Mammutfunde aus dem späten Mittelpleistozäns häufig die Bezeichnung Mammuthus trogontherii-primigenius verwendet, um diese Zwischenstellung anzuzeigen, für die Reste aus Steinheim, die rund 300.000 Jahre alt sind und rekonstruiert ein 3,7 m hohes Skelett ergeben, wurde gar der Begriff Mammuthus primigenius fraasi eingeführt. Manche Forscher stellten sogar den Artstatus des Steppenmammuts in Frage, doch wurde dies von den meisten strikt abgelehnt. Heute geht man dazu über, die älteren Funde des Steppenmammuths als Mammuthus trogontherii trogontherii und die jüngeren als Mammuthus trogontherii chosaricus zu bezeichnen.[1]
Allerdings traten bereits in der frühen Saale-Kaltzeit vor rund 250.000 Jahren erste Wollhaarmammuts in Europa auf und lebten gemeinsam mit den Steppenmammuts, wie z. B. das Mammut von Pfännerhall aus dem Geiseltal. In der späten Saalekaltzeit vor rund 200.000 Jahren sind dann im westlichen Eurasien nur noch Mammutformen nachweisbar, die von den typischen weichselkaltzeitlichen nicht mehr zu trennen sind. Zu solchen spätsaalekaltzeitlichen Wollhaarmammutfunden gehören Balderton (England) und Zemst (Belgien). Im Laufe der Weichsel-Kaltzeit kommt es dann erneut zu einer Größenreduktiion, die meist als weitere klimatische Anpassung verstanden wird. Einige Forscher gehen auch von einer fluktuierenden Körpergröße aus, die größere Individuen in warmklimatischen Phasen (Interstadiale) und kleinere in kühleren Phasen (Stadiale) umfassen, doch fehlt hier eine Korrelation mit absoluten Alterswerten. Da die Wollhaarmammuts teilweise regional deutliche Populationsunterschiede zeigen, gibt es in der Literatur eine unübersichtlich große Anzahl an Unterarten.[66][1]
Aussterben
→ Hauptartikel: Quartäre Aussterbewelle
Zeitraum des Aussterbens
Lange Zeit wurde angenommen, die letzten Wollmammuts Eurasiens seien bereits gegen Ende der letzten Kaltzeit vor rund 12.000 Jahren ausgestorben.[69][70][71] Sein häufigstes Auftreten hatte das Wollhaarnmammut allerdings vor dem letztglazialen Kältemaximum. Während des Kältemaximums verschwand es aus weiten Teilen des westlichen Eurasiens, um bei der darauf folgenden Wiedererwärmung (Bölling-Interstadial) zurückzukehren. Die Populationsdichte war aber jetzt wesentlich geringer und es erreichte auch Südeuropa nicht mehr.[71][8] Nach dem Ausklingen dieser Warmphase war es in West- und Zentraleuropa nur noch äußerst selten anzutreffen. Die jüngsten Daten stammen bisher aus Gough's Cave (England) und Les Coudrays (Frankreich) und liegen bei 14.600 bzw. 14.700 (kalibrierten) Jahren vor heute. Allerdings muss es noch rund 1.000 Jahre später hier anzutreffen gewesen sein, da aus Gönnersdorf nicht nur Mammutreste, sondern auch zahlreiche Abbildungen dieser Tierart vorliegen. In Nordeuropa hat sich das Wollhaarmammut noch bis in die letzte Abkühlungsphase (Jüngere Dryas) am Ende der Weichsel-Kaltzeit gehalten, während es in Osteuropa offensichtlich noch bis in den Beginn des Holozäns überlebte. Junge Funde stammen hier aus Puurmani (Estland) mit circa 10.100 bis 10.200 BP und Tscherepowez in Nordrussland mit etwa 9.800 BP.[6][72][73]
In Westsibirien trat die Art ebenfalls noch im Allerød-Interstadial auf, verschwand dann aber auch während der Jüngeren Dryas. Das jüngste Datum stammt aber von Funden aus dem Juribei und sind mit 11.700 Jahren vor heute in den Übergang vom Pleistozän zum Holozän zu stellen. Weiter ins Holozän hineinreichend lassen sich Populationen von der Taimyr-Halbinsel und auf der Tschuktschen-Halbinsel nachweisen. Hier liegen die jüngsten Daten von Funden von der Unteren Taimyra bei etwa 9.600 Jahren vor heute.[6][72]
Die letzten Angehörigen der Art lebten bis vor 3.700 Jahren auf der Wrangel-Insel, überlebten also vergleichbar bis in die Zeit der ägyptischen Pharaonen oder der mitteleuropäischen Bronzezeit. Aus dieser Zeit stammen auch die ältesten Hinterlassenschaften von Menschen auf dieser Insel, was die Vermutung nährt, der Mensch habe die Tiere durch starke Bejagung ausgerottet.[74][26]
Auf dem nordamerikanischen Kontinent hat das Wollhaarmammut offensichtlich ebenfalls bis in das frühe Holozän überlebt. Die letzten Vertreter auf dem Festland lassen sich vor rund 10.700 Jahren am Gebiet des Yukon nachweisen.[75] Auf der Sankt-Paul-Insel, die zu den Pribilof-Inseln gehört und vor 13.000 Jahren von Alaska durch das steigende Meerwasser getrennt wurde, entdeckte man eine Population, die noch bis ins mittlere Holozän überlebte. Es handelt sich um relativ kleine Wollhaarmammuts, deren geringe Körpergröße jedoch nicht als eine vollständige Inselverzwergung wie auf der Wrangel-Insel angesehen wird. Die jüngsten Daten aus der Qagnax-Höhle von Sankt Paul liegen bei 5.725 BP (14C-Jahre).[76][77]
Theorien zum Aussterben
- Overkill-Hypothese: Das Wollhaarmammut war eines der Jagdtiere der Menschen im Jungpleistozän. Dies ist durch zahlreiche Höhlenmalereien und eine Vielzahl von Mammutknochen-Anhäufungen in archäologischen Fundstellen des Aurignacien, Gravettien und Epigravettien dokumentiert. Ob eine übermäßige Bejagung („Overkill-Hypothese“) das Aussterben der Tiere verursacht hat oder rasche Klimaveränderungen zum Ende der Eiszeit (Erwärmung im Allerød-Interstadial), ist bis heute umstritten. Da das Mammut und andere eiszeitliche Säuger schon vorher viele heftige Klimaschwankungen überlebt hatten, ist ein menschlicher Einfluss auf das vollständige Verschwinden plausibel. Eine Studie von C. Johnson deutet darauf hin, dass das Aussterben des Wollhaarmammuts und anderer pleistozäner Arten mit einer rapiden Abnahme der Fruchtbarkeit einherging.[78] Eine zu geringe Reproduktionsrate sieht er bei einer Reihe von Großsäugern in Australien, Eurasien, Amerika und Madagaskar als Hauptursache des Aussterbens, während er die „Overkill-Hypothese“ (bei ihm: „Blitzkrieg-Hypothese“) als ursächliches Szenario zurückweist. Da Arten mit zurückgehender Reproduktionsrate bei menschlicher Bejagung zusätzlichem Stress ausgesetzt sind, sei die Gleichzeitigkeit des Aussterbens mit der verstärkten Bejagung durch Jäger und Sammler-Populationen die logische Folge.
- Impakt-Hypothese: 2007 publizierten Richard B. Firestone et al. eine Hypothese, der zufolge der Einschlag (Impakt) eines oder mehrerer großer astronomischer Objekte geringer Dichte vor etwa 12.900 Jahren (12.900 ± 100 Kalenderjahren cal BP) im nördlichen Nordamerika die Ursache für die unvermittelt einsetzende starke Abkühlung der Jüngeren Dryas gewesen ist. Mittelbare Folgen seien das Aussterben der pleistozänen Megafauna in Nordamerika (einschließlich des Wollhaarmammuts) sowie der Untergang der prähistorischen Clovis-Kultur der Paläoindianer gewesen.[79] Eine 2008 veröffentlichte Untersuchung der demographischen Entwicklung der Paläoindianer für den fraglichen Zeitraum ergab jedoch keinen Hinweis auf den in der Hypothese genannten Bevölkerungsrückgang.[80]
Systematik
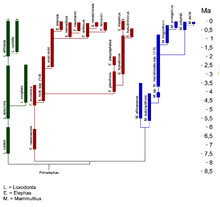 Evolution der Elephantidae
Evolution der ElephantidaeDas Wollhaarmammut als Vertreter der Gattung Mammuthus gehört zur Familie der Eigentlichen Elefanten, der weiterhin die rezenten Gattungen Elephas mit dem Asiatischen Elefanten, Loxodonta mit dem Afrikanischen Elefanten und dem Waldelefanten (Loxodonta cyclotis) und die ausgestorbene Gattung Primelephas zugewiesen werden. Allen Vertretern der Elefanten gemein ist der lamellenartige Aufbau und die ausgeprägte Hochkronigkeit (Hypsodontie) der Backenzähne bzw. die Bildung der Stoßzähne aus den oberen Schneidezähnen.[81][82]
Dabei ist das Wollhaarmammut aus dem Steppenmammut hervorgegangen und bildet mit ihm eine Schwesterlinie zum Präriemammut und dem Zwergmammut (Mammuthus exilis) auf dem amerikanischen Kontinent. Molekulargenetischen Untersuchungen zufolge trennten sich beide Linien vor etwa 2 Mio. Jahren.[76] Auch das Genom dieser Tierart, das aus über 4 Gb (Giga-Basenpaare) besteht, wurde untersucht und mit Hilfe neuer DNA-Sequenzierungsmethoden konnte ein großer Teil (3,3 Gb) davon entschlüsselt werden. Der daraus resultierende, geschätzte, genetische Unterschied zwischen Wollhaarmammut und Afrikanischem Elefant betrug weniger als 0,6 % und ist nur etwa halb so groß wie der zwischen Schimpanse und Mensch. Mit dem Asiatischen Elefanten ist das Wollhaarmammut noch näher verwandt.[83]. In absoluten Altersdaten ausgedrückt bedeutet dies, dass sich die Linien Elephas und Mammuthus vor 6,7 Mio. Jahren trennten, während Loxodonta sich schon vor 7,6 Mio. Jahren abgespaltet hatte. Die Echten Mastodonten, eine urtümliche Rüsseltierart, zu denen auch das bis ins Jungpleistozän überlebende Amerikanische Mastodon gehört, gingen bereits vor 26 Mio. Jahren ihren eigenen evolutiven Weg.[84][85][86]
Schon Anfang des 21. Jh. zeigten DNA-Untersuchungen an immerhin sechs verschiedenen Individuen, dass das Wollhaarmammut offenbar aus zwei genetischen Gruppen besteht.[87] Spätere Analysen an mindestens 18 sibirischen Funden - u. a. vom Jarkow-Mammut, dem Fischhaken-Mammut, Dima und vom Adams-Mammut - bestätigten diese Ansicht. Die beiden Wollhaarmammut-Linien wurden dabei Klade I und Klade II genannt. Während Vertreter von Klade I weit über das nördliche Eurasien und Nordamerika verbreitet waren, beschränkten sich jene der Klade II bisher auf die Region zwischen der Lena und der Kolyma. Außerdem scheint sie schon vor wenigstens 33.000 Jahren erloschen zu sein.[85] Was die Trennung der beiden Gruppen verursachte, ob Selektion oder Gendrift ist bisher nicht geklärt, anatomische oder funktionelle Änderungen scheinen sie nicht bewirkt zuhaben. Die Aufspaltung der beiden Kladen geschah nach bisherigen Untersuchungen bereits vor ein oder zwei Mio. Jahren.[85][83]
Darüber hinaus zeigten Analysen, die an der mitochondrialen DNA von 160 Mammutindividuen aus der gesamten Holarktis erfolgten, dass sich das Wollhaarmammmut offenbar nicht nur über die Beringstraße nach Nordamerika ausbreitete, sondern auch gelegentlich nach Eurasien zurückkehrte und so ein dynamisches Besiedlungsverhalten besaß. Dabei wurden fünf Haplogruppen (A bis E) identifiziert, von denen die Gruppe C ursprünglich bis vor 47.000 Jahren nur in Nordamerika beheimatet war und offensichtlich die ursprüngliche Einwanderungswelle darstellte. Bis vor 22.000 Jahren wanderten die Vertreter der vier anderen Gruppen ebenfalls über die Bering-Straße nach Nordamerika, während die dortige Population nun auch im nördlichen Eurasien anzutreffen war. Die gemischten Gruppen überlebten dann bis zum Ende der letzten Vereisungsphase, während die Vertreter der Gruppe E die letzten Wollhaarmammuts auf der Wrangel-Insel waren.[88]
Weiterhin wies der DNA-Code für das Hämoglobin eines 43.000 Jahre alten Wollhaarmammuts drei vom Hämoglobin eines Asiatischen Elefanten abweichende Sequenzen auf. Das Hämoglobin-Gen des Wollhaarmammuts wurde 2010 in Bakterien eingeschleust, worauf diese das Mammutprotein produzierten. Aus der unterschiedlichen Sauerstoff-Sättigung zwischen dem Hämoglobin des Wollhaarmammuts und dem des Asiatischen Elefanten erhofft man sich Erkenntnisse über die Kälteanpassung der Mammuts zu gewinnen.[89].
Forschungsgeschichte
 Johann Friedrich Blumenbach
Johann Friedrich BlumenbachFunde von Wollhaarmammuts wurden schon sehr früh im westlichen Eurasien entdeckt, häufig aber nicht als fossile Elefantenart erkannt. Die 1577 beim Kloster Reyden (Luzern) entdeckten Mammutknochen wurden vom damaligen Arzt F. Plater als Überbleibsel eines 19 Fuß großen Riesen gedeutet.[90] Ebenso wurde das 1663 bei Seveckenberg bei Quedlinburg (Sachsen-Anhalt) im Beisein des Naturforschers Otto von Guericke (1602–1686) ausgegrabene Skelett eines Wollhaarmammuts als Rest des mythischen Unicornu fossile angesehen - ähnlich erging es auch den rund 33 Jahre später geborgenen ersten Funden eines Europäischen Waldelefanten bei Tonna (Thüringen) - und später auch als solches rekonstruiert.[10] Im Jahr 1696 beschrieb dann der Russe Ludloff Reste eines in Sibirien vorkommenden Tieres, welches die einheimischen Jakuten und Tungusen als Mamantu bezeichneten und nach ihrer Meinung einem riesigen Maulwurf glich, der sterben müsste, sobald er das Sonnenlicht erblickt.[13]
Die Herkunft des Begriffes Mamantu oder Maimanto ist nicht geklärt. Häufig wird ein Ursprung im nenzischen oder estnischen Sprachraum gesucht. In letzterer Sprache bedeutet maa "Erde" und mutt "Maulwurf". Allerdings wird auch gelegentlich eine Verbindung zum arabischen Wort Behemoth - einem gewaltigen Ungeheuer mit gekrümmten Hörnern und Stoßzähnen - gezogen und mit dem seit dem 9. Jh. nachgwíesenen Handel von sibirischem Elfenbein durch Araber begründet. Auch wer den Namen letztendlich in Europa einführte, ist nicht hinreichend gesichert.[13][5]
Erstmals wissenschaftlich beschrieben wurde das Wollhaarmammut 1799 vom Göttinger Naturforscher und Anatomen Johann Friedrich Blumenbach (1752–1840) als Elephas primigenius anhand von Funden bei Osterode am Harz. Im selben Jahr wurde der Kadaver des Adams-Mammut an der Lena entdeckt. Die von Roman Boltunow 1806 angefertigte Skizze dieses Fundes leitete später Henry Michail Adams nach Göttingen an Blumenbach weiter, der darin seine kürzlich beschriebene Elefantenart erkannte.[13] Der Gattungsname Mammuthus stammt vom englischen Biologen Joshua Brookes (1761–1833), den er 1828 einführte.
Kulturgeschichte
Als ausgestorbenes Tier hatte das Wollhaarmammut schon früh in der menschlichen Geschichte der Nach-Eiszeit eine hohe Bedeutung. Die einheimischen Völker Sibiriens und des nördlichen Nordamerikas nutzten vornehmlich Mammutelfenbein für Schnitzereien. Seit dem 9. Jh. ist der Handel mit fossilem Elfenbein aus Jakutien durch Araber belegt, später wurde dieser durch das Russische Kaiserreich fortgesetzt. von 1800 bis 1914 sind Schätzungen zufolge zwischen 20 und 32 t Elfenbein jährlich gehandelt worden, was etwa einer Anzahl von 25.000 bis 46.000 Mammutindividuen insgesamt entspricht. Seit 1989 der Handel mit Elfenbein lebender Elefanten untersagt wurde, hat fossiles Elfenbein auch heute wieder eine erhebliche Bedeutung, u. a. für die Elfenbeinschnitzerei. In China wurden fossile Mammutstoßzähne als Zähne von "Eisratten" angesehen und ähnlich den als Drachenknochen eingeschätzten pleistozänen Tierresten zu Pulver zermahlen und als Arzenei verkauft. In Europa galten sie bis in die Neuzeit teilweise als Überreste des Einhorns und hatten ebenfalls zu Pulver zerkleinert als Heilmittel Bedeutung.[31]
Doch auch nach seiner wissenschaftlichen Beschreibung ist das Mammut in die menschliche Kultur eingegangen. Der Begriff Mammut steht heute für etwas großes oder kaum zu bewältigendes (so u. a. "Mammutaufgabe"). Es fand außerdem Einzug in die Heraldik und ist in den Wappen von Srednekolymsk (Republik Sacha, Russland) oder aber von Seedorf (Niedersachsen) vertreten. Darüber hinaus ist es ein beliebtes Element in der modernen Popkultur und tritt in Filmen (u. a. Am Anfang war das Feuer von Regisseur Jean-Jacques Annaud) und Romanen (so im Zyklus Kinder der Erde von Jean M. Auel auf.
Literatur
- Miles Barton: Wildes Amerika. Zeugen der Eiszeit. Vgs, Köln 2003, ISBN 3-8025-1558-7.
- Wighart von Koenigswald: Lebendige Eiszeit. Theiss-Verlag, Stuttgart Sigmaringen 2002, ISBN 3-8062-1734-3.
- Adrian Lister, Paul Bahn: Mammuts – Die Riesen der Eiszeit. Jan Thorbecke Verlag, 1997 ISBN 3-7995-9050-1.
- Ronald M. Nowak: Walker’s Mammals of the World. 6th edition. Johns Hopkins University Press, Baltimore MD u. a. 1999, ISBN 0-8018-5789-9.
Einzelnachweise
- ↑ a b c d e f g h Ralf-Dietrich Kahlke: Die Entstehungs-, Entwicklungs- und Verbreitungsgeschichte des oberpleistozönen Mammuthus-Coelodonta-Faunenkomplexes in Eurasien (Großsäuger). Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft 546 Frankfurt am Main, 1994
- ↑ a b c d e Diego J. Álvarez-Lao, Ralf-Dietrich Kahlke, Nuria García, and Dick Mol: The Padul mammoth finds — On the southernmost record of Mammuthus primigenius in Europe and its southern spread during the Late Pleistocene. Palaeogeography, Palaeoclimatology, Palaeoecology. 278, 2009, S. 57–70
- ↑ Diego J. Álvarez-Lao und Nuria García: Geographical distribution of Pleistocene cold-adapted large mammal faunas in the Iberian Peninsula. Quaternary International 233, 2011, S. 159-170
- ↑ a b c d e f Dick Mol, Alexei N. Tikhonov, Johannes van der Pflicht und Dmitry Yu. Bolshiyanov: Discoveries of woolly mammoth, Mammuthus primigenius (Proboscidea: Elephantidae) and some other Pleistocene mammals on the Taimyr Peninsula. Russian Journal of Theriology 2 (2), 2003, S. 77-95
- ↑ a b c d e f g h i j k l m n Adrian Lister und Paul Bahn: Mammuts – Die Riesen der Eiszeit. Sigmaringen, 1997
- ↑ a b c Yaroslav V. Kuzmin: Extinction of the woolly mammoth (Mammuthus primigenius) and woolly rhinoceros (Coelodonta antiquitatis) in Eurasia: Review of chronological and environmental issues. Boreas 39, 2010, S. 247-261
- ↑ a b Ute Koch: Lebensraum Mammutsteppe. In: Ulrich Joger und Ute Koch (Hrsg.): Mammuts aus Sibirien. Darmstadt, 1994, S. 55-73
- ↑ a b c d e f g h Rudolf Musil: Überlebenstrategien von Elefanten. In: Harald Meller (Hrsg.): Elefantenreich - Eine Fossilwelt in Europa. Halle/Saale, 2010, S. 323-336
- ↑ A. K. Markova, A. N. Simakova, A. Yu. Puzachenko und L. M. Kitaev: Eastern European mammoth distribution and environments during the Middle Valdai Briansk Interstade (33,000-24,000 BP). In: G. Cavarretta et al. (Eds.): The World of Elephants - International Congress. Consiglio Nazionale delle Ricerche Rom, 2001, S. 299-304
- ↑ a b c d e f g h Wighart von Koenigswald: Lebendige Eiszeit. Klima und Tierwelt im Wandel. Stuttgart, 2002, S. 54–61
- ↑ Wei Guang Biao, Hu Song Mei, Yu Ke Fu, Hou Ya Mei, Li Xin, Jin Chang Zhu, Wang Yuan, Zhao Jian Xin und Wang Wen Hua: New materials of the steppe mammoth, Mammuthus trogontherii, with discussion on the origin and evolutionary patterns of mammoths. Science China - Earth sciences 53 (7), 2010, S. 956-963
- ↑ G. Haynes und J. Klimowicz: Mammoth (Mammuthus spp.) and American mastodont (Mammut americanum) bonesites: what do the differences mean? Deinsea 9, 2003, S. 185-204
- ↑ a b c d e f Ulrich Joger: Geschichte(n) der Mammutfunde. In: Ulrich Joger und Ute Koch (Hrsg.): Mammuts aus Sibirien. Darmstadt, 1994, S. 9-23
- ↑ a b Gennady G. Boeskorov and Dick Mol: Quaternary Mammal Collections in the Museums of Yakutsk (Eastern Siberia, Yakutia, Russia). Cranium 21 (1-2), 2004, S. 19-32
- ↑ Volker Toepfer: Die Mammutfunde von Pfännerhall im Geiseltal. Veröffentlichungen des Landesmuseums für Vorgeschichte in Halle 16.Halle/Saale, 1957
- ↑ a b c Ralf-Dietrich Kahlke und Dick Mol: Eiszeitliche Großsäugetiere der Sibirischen Arktis. Die Cerpolex/Mammuthus-Expeditionen auf Tajmyr. E. Schweizerbart'sche Verlagsbuchhandlung (Nägele u. Obermiller), Stuttgart 2005
- ↑ C. R. Harington (Dezember 1995): „Woolly Mammoth“. Aus: Canadian Museum of Nature in Yukon Beringia Interpretive Centre.
- ↑ Dick. Mol, Y. Coppens, A. N. Tikhonov, L. D. Agenbroad, R. D. E. MacPhee, C. Flemming, A. Greenwood, B. Buigues, C. de Marliave, B. van Geel, G. B. A. van Reenen, J. P. Pals, D. C. Fisher und D. Fox: The Jarkov Mammoth: 20,000-Year-Old carcass of a Siberian woolly mammoth Mammuthus primigenius (Blumenbach, 1799). In: G. Cavarretta et al. (Eds.): The World of Elephants - International Congress. Consiglio Nazionale delle Ricerche Rom, 2001, S. 306-309
- ↑ Dick Mol und Ralf-Dietrich Kahlke: The fishhook mammoth - a Late Pleistocene (Early Sartanian) individual of Mammuthus primigenius with preserved gut contents from the Upper Taimyra River (Taimyr Peninsula, Arctic Siberia). Abstracts of the 18th International Senckenberg Conference 25. bis 30. April 2004 in Weimar
- ↑ Dick Mol, A. N. Tikhonov, R. D. E. MacPhee, C. Flemming, B. Buigues, C. de Marliave, Y. Coppens und L.D. Agenbroad: The Fishhook Mammoth: rediscovery of a Woolly Mammoth carcass by the CERPOLEX/Mammuthus Team, Taimyr Peninsula, Siberia. In: G. Cavarretta et al. (Eds.): The World of Elephants - International Congress. Consiglio Nazionale delle Ricerche Rom, 2001, S. 310-313
- ↑ a b c Dick Mol, Jeheskel (Hezy) Shoshani, Alexei Tikhonov, Bas van Geel, Shin-ichi Sano, Peter Lazarev, Gennady Boeskorov und Larry D. Agenbroad: The Yukagir Mammoth: Brief history, 14C Dates, individual age, gender, sze, physical and environmental conditions and storage. Scientific Annals, School of Geology Aristotle University of Thessaloniki (AUTH), special volume 98, 2006, S. 310-313
- ↑ Tom Mueller (Mai 2009): Ice Baby A near-perfect frozen mammoth resurfaces after 40,000 years, bearing clues to a great vanished species. National Geographic (abgerufen 30. April 2011)
- ↑ Maciej J. Kotarba, Marek Dzieniewicz, Włodzimierz J. Mościcki und enryk Sechman: Unique Quaternary environment for discoveries of woolly rhinoceroses in Starunia, fore-Carpathian region, Ukraine: Geochemical and geoelectric studies. Geology 36 (7), 2008, S. 567-570
- ↑ Reinhard Ziegler: An extraordinary small mammoth (Mammuthus primigenius) from SW Germany. Stuttgarter Beiträge zur Naturkunde Serie B 300, 2001, S. 1-41
- ↑ a b c d e f g h Ute Koch: Unser heutiges Bild vom Mammut. In: Ulrich Joger und Ute Koch (Hrsg.): Mammuts aus Sibirien. Darmstadt, 1994, S. 29-32
- ↑ a b Adrian M. Lister: Mammoths in miniature. Nature 362, 1993, S. 288-289
- ↑ http://www.badische-zeitung.de/bildung-wissen-1/ein-mammut-projekt--8100204.html 2006 gelang es ein Gen aus dem Zellkern eines Mammuts zu rekonstruieren, das für die Fellfarbe verantwortlich war. Dies belegte dass es auch blonde Mammuts gab.
- ↑ http://www.wissenschaft.de/wissenschaft/news/267155.html Bild der Wissenschaft – Rüsseltiere mit Blondschopf
- ↑ Karol Schauer: Wie sahen sie aus? Zur Rekonstruktion des Elephas (Palaeoloxodon) antiquus aus den Seeablagerungen von Neumark-Nord. In: Harald Meller (Hrsg.): Elefantenreich - Eine Fossilwelt in Europa. Halle/Saale, 2010, S. 296-313
- ↑ Karlheinz Fischer: Die Waldelefanten von Neumark-Nord und Gröbern. In: Dietrich Mania u. a. (Hrsg.): Neumark-Nord - Ein interglaziales Ökosystem des mittelpaläolithischen Menschen. Veröffentlichungen des Landesmuseums für Vorgeschichte in Halle 62. Halle/Saale, 2010, S. 361-374
- ↑ a b c Arun Banerjee: Das Mammutelfenbein. In: Ulrich Joger und Ute Koch (Hrsg.): Mammuts aus Sibirien. Darmstadt, 1994, S. 38-42
- ↑ Martina Ábelová: Schreger pattern analysis of Mammuthus primigenius tusk: analytical aproach and utility. Bulletin of Geosciences 83 (2), 2008, S. 225-232
- ↑ Grant D. Zazula, Duane G. Froese, Alice M. Telka, Rolf W. Mathewes und John A. Westgate: Plants, bugs, and a giant mammoth tusk: Paleoecology of Last Chance Creek, Yukon Territory. Yukon Explorationand Geology, 2002, S. 252-258
- ↑ R. D. Guthrie: Reconstructions of Woolly Mammoth life history. In: G. Cavarretta et al. (Eds.): The World of Elephants - International Congress. Consiglio Nazionale delle Ricerche Rom, 2001, S. 276-279
- ↑ Jeheskel Shoshani: Understanding proboscidean evolution: a formidable task. Tree 13, 1998, S. 480-487
- ↑ Hans van Essen: A supernumerary tooth and an odontoma attributable to Mammuthus primigenius (BLUMENBACH, 1799) (Mammalia, Proboscidea) from The Netherlands, and various related finds. Acta zoologica cracoviensia, 47 (1-2), 2004, S. 81-121
- ↑ Irina V. Kirillova: Unusual tooth pathology in mammoth (Mammuthus primigenius) from Yakutia. Russian Journal of Theriology 8 (1), 2009, S. 29-36
- ↑ a b c Hans-Peter Hock: Die Welt der Mammutjäger. In: Ulrich Joger und Ute Koch (Hrsg.): Mammuts aus Sibirien. Darmstadt, 1994, S. 78-93
- ↑ Sabine Gaudzinski: Ein mittelpaläolithisches Rentierlager bei Salzgitter-Lebenstedt. In: Manfred Boetzkes, Ingeborg Schweitzer und Jürgen Vespermann (Hrsg.): EisZeit - Das große Abenteuer der Naturbeherrschung. Begleitbuch zur gleichnamigen Ausstellung. Hildesheim, 1999, S. 121-136
- ↑ Dietrich Mania und V. Toepfer: Königsaue. Gliederung, Ökologie und mittelpaläolithische Funde der letzten Eiszeit. Veröffentlichungen des Landesmuseums für Vorgeschichte Halle/Saale 26, Berlin, 1973
- ↑ Margherita Mussi und Paola Villa: Single carcass of Mammuthus primigenius with lithic artifacts in the Upper Pleistocene of northern Italy. Journal of Archaeological Science 35, 2008, S. 2606–2613
- ↑ a b Sergey A. Vasil’ev: Man and Mammoth in Pleistocene Siberia. In: G. Cavarretta et al. (Eds.): The World of Elephants - International Congress. Consiglio Nazionale delle Ricerche Rom, 2001, S. 363-366
- ↑ a b c d Yaroslav V. Kutzmin: Mammalian Fauna from Palaeolithic sites in the Upper Yenisei River Basin (Southern Siberia): Review of the current zooarchaeological evidence. International Journal of Osteoarchaeology 21, 2011, S. 218-228
- ↑ G. Tromnau: Ein Mammutknochen-Faustkeil aus Rhede, Kr. Borken (Westfalen). Archäologisches Korrespondenzblatt 13, 1983, S. 287-289
- ↑ Karel Valoch: Die Erforschung der Kůlna-Höhle 1961-1976. Anthropos 24, Brno, 1988
- ↑ a b V. T. Dobosi: Ex Proboscideis - Proboscidean remains as raw material at four Palaeolithic sites, Hungary. In: G. Cavarretta et al. (Eds.): The World of Elephants - International Congress. Consiglio Nazionale delle Ricerche Rom, 2001, S. 429-431
- ↑ Joachim Schäfer: Die Wertschätzung außergewöhnlicher Gegenstände (non-utilitarian objects) im Alt- und Mittelpaläolithikum. Ethnographisch-Archäologische Zeitschrift 36, 1996, S. 173-190
- ↑ Laura Niven: The role of mammoths in Upper Palaeolithic economies of southern Germany. In: G. Cavarretta et al. (Eds.): The World of Elephants - International Congress. Consiglio Nazionale delle Ricerche Rom, 2001, S. 323-327
- ↑ Laura Niven: From carcass to cave: Large mammal exploitation during the Aurignacian at Vogelherd, Germany. Journal of Human Evolution 53, 2007, S. 362e382
- ↑ F. A. Fladerer: The Krems-Wachtberg camp-site: mammoth carcass utilization along the Danube 27,000 years ago. In: G. Cavarretta et al. (Eds.): The World of Elephants - International Congress. Consiglio Nazionale delle Ricerche Rom, 2001, S. 432-438
- ↑ Bohuslav Klíma: Die große Anhäufung von Mammutknochen in Dolní Vestonice. Acta Scientiarum Naturalium Brno 3 (6), 1969, S. 1-52
- ↑ Martin Oliva: Some thoughts on pavlovian adaptions and their alternatives. In: Wil Roebroeks, Margherita Mussi, Jiri Svoboda und Kelly Fennema (Hrsg.): Hunters of the Golden Age. Leiden, 2000, S. 219-230
- ↑ I. G. Pidoplichko: Upper Palaeolithic dwellings of mammoth bones in the Ukraine. In: BAR International Series Nr. 712, 1998
- ↑ Jean-Marie Chauvet, Éliette Brunel Deschamps und Christian Hillaire: Grotte Chauvet bei Vallon -Pont-d'Arc. Altsteinzeitliche Höhlenkunst im Tal der Ardèche Sigmaringen, 1995
- ↑ Lothar Zotz: Die Wiederentdeckung der Merkwürdigkeiten von Rouffignac. In: Orion, Heft 3, 12, 1957, S. 169-174
- ↑ a b Michel Lorblanchet: Höhlenmalerei. Ein Handbuch. Sigmaringen, 1997
- ↑ a b c Hans-Peter Hock: Kunst der Eiszeit. In: Ulrich Joger und Ute Koch (Hrsg.): Mammuts aus Sibirien. Darmstadt, 1994, S. 94-100
- ↑ a b c d e Gerhard Bosinski: Die große Zeit der Eiszeitjäger. Europa zwischen 40.000 und 10.000 v. Chr. Jahrbuch des Römisch-Germanischen Zentralmuseums Mainz 34, 1987, S. 3-139
- ↑ Gerhard Bosinski: Das Aussehen des Mammuts nach den zeitgenössischen Darstellungen von Gönnersdorf. In: Ulrich Joger und Ute Koch (Hrsg.): Mammuts aus Sibirien. Darmstadt, 1994, S. 101-109
- ↑ Gerhard Bosinski: Die Kunst der Eiszeit in Deutschland und der Schweiz. Kataloge Vor- und Frühgeschichtlicher Altertümer 20, Bonn, 1982
- ↑ Barbara A. Purdy, Kevin S. Jones, John J. Mecholsky, Gerald Bourne, Richard C. Hulbert Jr., Bruce J. MacFadden, Krista L. Church und Michael W. Warren: Earliest Art in the Americas: Incised Image of a Mammoth on a Mineralized Extinct Animal Bone from the Old Vero Site (8-Ir-9), Florida. Congrès de l’IFRAO, septembre 2010 – Symposium : L’art pléistocène dans les Amériques (Pré-Actes) / IFRAO Congress, September 2010 – Symposium: Pleistocene art of the Americas (Pre-Acts), 2010, S. 3-12
- ↑ Nicholas J. Conard, Maria Malina2 und Susanne C. Mu¨nzel: New flutes document the earliest musical tradition in southwestern Germany. Nature 460, 2009, S. 737-740
- ↑ Jiři Svoboda, Vojen Ložek und Emanuel Vlček: Hunters between East and West. The Palaeolithic of Moravia. New York, London, 1996
- ↑ Marc Händel, Thomas Einwögerer und Ulrich Simon:: Krems-Wachtberg – A Gravettian Settlement Site in the Middle Danube Region. Wissenschaftliche Mitteilungen des Niederösterreichischen Landesmuseumeums 19, 2008, S. 91-108
- ↑ Adrian M. Lister und Hans van Essen: The earliest Mammoth in Europe. Abstracts of the 18th International Senckenberg Conference 25. bis 30. April 2004 in Weimar
- ↑ a b c d Adrian M. Lister, Andrei V. Sher, Hans van Essen und Guangbiao Wei: The pattern and process of mammoth evolution in Eurasia. Quaternary International 126–128, 2005, S. 49–64
- ↑ Marco P. Ferretti: Structure and evolution of mammoth molar enamel. Acta Palaeontologija Polonica 48 (3), 2003, S.: 383–396
- ↑ a b Adrian M. Lister und Andrei V. Sher: The Origin and Evolution of the Woolly Mammoth. Science 294, 2001, S. 1094-1097
- ↑ Anthony J. Stuart: Mammalian extinctions in the late Pleistocene of Northern Eurasia and North America. Biological Reviews 66, 1991, S. 453–562
- ↑ Anthony J. Stuart: Late Pleistocene megafaunal extinctions; a European perspective. In: R. D. E. MacPhee (Hrsg.): Extinctions in Near Time: Causes, Contexts and Consequences. Kluwer Academic/Plenum Publishers, New York, 1999.
- ↑ a b Anthony J. Stuart, The extinction of woolly mammoth (Mammuthus primigenius) and straight-tusked elephant (Palaeoloxodon antiquus) in Europe. Quaternary International 126–128, 2005, S. 171–177.
- ↑ a b Anthony J. Stuart, Leopold D. Sulerzhitsky, Lyobov A. Orlova, Yaroslav V. Kuzmin und Adrian M. Lister: The latest woolly mammoths (Mammuthus primigenius Blumenbach) in Europe and Asia: a review of the current evidence. Quaternary Science Reviews 21, 2002, S. 1559–1569
- ↑ Linas Daugnora: Mamutai Lietuvoje. Lietuvos Archeologija 25, 2004, S. 9-16
- ↑ David L. Fox, Daniel C. Fisher, Sergey Vartanyan, Alexei N. Tikhonov, Dick Mol, Bernard Buigues: Paleoclimatic implications of oxygen isotopic variation in late Pleistocene and Holocene tusks of Mammuthus primigenius from northern Eurasia. In: Quaternary International 169–170, 2007, S. 154–165
- ↑ Gary Haynes: Extinctions in North America’s Late Glacial landscapes. Quaternary International 2010, S. 1-10>
- ↑ a b J. M. Enk, D. R. Yesner, K. J. Crossen, D. W. Veltre und D. H. O’Rourke: Phylogeographic Analysis of the mid-Holocene Mammoth from Qagnax Cave, St. Paul Island, Alaska. Palaeogeography, Palaeoclimatology, Palaeoecology 273 (1-2), 2009, S. 184-190
- ↑ Douglas W. Veltre, David R. Yesner, Kristine J. Crossen, Russell W. Graham und Joan B. Coltrain: Patterns of faunal extinction and paleoclimatic change from mid-Holocene mammoth and polar bear remains, Pribilof Islands, Alaska. Quaternary Research 2008, S. 40-50
- ↑ C. N. Johnson: Determinants of loss of mammal species during the Late Quaternary ‘megafauna’ extinctions: life history and ecology, but not body size. In: Proceedings of the Royal Society of London, Band 269, 2002, S. 2221–2227.
- ↑ R. B. Firestone u. a.: Evidence for an extraterrestrial impact 12,900 years ago that contributed to the megafaunal extinctions and Younger Dryas cooling. In: Proceedings of the National Academy of Sciences, Band 104, 2007, S. 16016–16021. Abstract
- ↑ Briggs Buchanan, Mark Collard, Kevan Edinborough: Paleoindian demography and the extraterrestrial impact hypothesis. In: Proceedings of the National Academy of Sciences 105, 2008, S. 11651–11654. Abstract
- ↑ Jan van der Made: The evolution of the elephants and their relatives in context of a changing climate and geography. In: Harald Meller (Hrsg.): Elefantenreich - Eine Fossilwelt in Europa. Halle/Saale, 2010, S. 340-360
- ↑ Ute Koch: Das Mammut und seine Verwandten. In: Ulrich Joger und Ute Koch (Hrsg.): Mammuts aus Sibirien. Darmstadt, 1994, S. 33-37
- ↑ a b Webb Miller, Daniela I. Drautz, Aakrosh Ratan, Barbara Pusey, Ji Qi, Arthur M. Lesk, Lynn P. Tomsho, Michael D. Packard, Fangqing Zhao, Andrei Sher, Alexei Tikhonov, Brian Raney, Nick Patterson, Kerstin Lindblad-Toh, Eric S. Lander, James R. knight, Gerard P. Irzyk, Karin m. Fredrikson, Tomothy T. harkins, Sharon Sheridan, Tom Pringle &Stephan C. Schuster: Sequencing the nuclear genome of the extinct woolly mammoth. Nature 456, 2008, S. 387–390
- ↑ Nadin Rohland, Anna-Sapfo Malaspinas, Joshua L. Pollack, Montgomery Slatkin, Paul Matheus und Michael Hofreiter: Proboscidean Mitogenomics: Chronology and Mode of Elephant Evolution Using Mastodon as Outgroup. PLOSBiology 5 (August), 2007, S. 1663-1671
- ↑ a b c M. Thomas P. Gilbert, Daniela I. Drautz, Arthur M. Lesk, Simon Y. W. Ho, Ji Qi, Aakrosh Ratan, Chih-Hao Hsu, Andrei Sher, Love Dalén, Anders Götherström, Lynn P. Tomsho, Snjezana Rendulic, Michael Packard, Paula F. Campos, Tatyana V. Kuznetsova, Fyodor Shidlovskiy, Alexei Tikhonovj, Eske Willerslev, Paola Iacumin, Bernard Buigues, Per G. P. Ericson, Mietje Germonpré, Pavel Kosintsev, Vladimir Nikolaev, Malgosia Nowak-Kemp, James R. Knight, Gerard P. Irzyk, Clotilde S. Perbost, Karin M. Fredrikson, Timothy T. Harkins, Sharon Sheridan, Webb Miller und Stephan C. Schuster: Intraspecific phylogenetic analysis of Siberian woolly mammoths using complete mitochondrial genomes. PNAS 105 (24), 2008, S. 8327-8332
- ↑ Cristian Capelli, Ross D. E. MacPhee, Alfred L. Roca, Francesca Brisighelli, Nicholas Georgiadis, Stephen J. O’Brien und Alex D. Greenwood: A nuclear DNA phylogeny of the woolly mammoth (Mammuthus primigenius). Molecular Phylogenetics and Evolution 40, 2006, S. 620–627
- ↑ G. S. Rautian und I. A. Dubrovo: Genetic data indicating that Mammuthus primigenius includes two species. In: G. Cavarretta et al. (Eds.): The World of Elephants - International Congress. Consiglio Nazionale delle Ricerche Rom, 2001, S. 552-556
- ↑ Regis Debruyne, Genevieve Chu, Christine E. King, Kirsti Bos, Melanie Kuch, Carsten Schwarz, Paul Szpak, Darren R. Gröcke, Paul Matheus, Grant Zazula, Dale Guthrie, Duane Froese, Bernard Buigues, Christian de Marliave, Clare Flemming,8 Debi Poinar, Daniel Fisher, John Southon, Alexei N. Tikhonov, Ross D.E. MacPhee, und Hendrik N. Poinar1: Out of America: Ancient DNA Evidence for a New World Origin of Late Quaternary Woolly Mammoths. Current Biology 18, 2008, S. 1–7
- ↑ Kevin L Campbell, Jason E. E. Roberts, Laura N. Watson, Jörg Stetefeld, Angela M. Sloan, Anthony V. Signore, Jesse W. Howatt, Jeremy R. H. Tame, Nadin Rohland, Tong-Jian Shen, Jeremy J. Austin, Michael Hofreiter5, Chien Ho, Roy E. Weber und Alan Cooper: Substitutions in woolly mammoth hemoglobin confer biochemical properties adaptive for cold tolerance. In: Nature Genetics 42, 2010, S. 536–540 doi:10.1038/ng.574
- ↑ Hans-Dietrich Kahlke: Das Eiszeitalter. Leipzig, Jena, Berlin, 1981
Weblinks
 Commons: Wollhaarmammut – Sammlung von Bildern, Videos und Audiodateien
Commons: Wollhaarmammut – Sammlung von Bildern, Videos und Audiodateien- Caitlin Sedwick (2008): „What Killed the Woolly Mammoth?“. In: PLoS Biol 6(4): e99 doi:10.1371/journal.pbio.0060099
Kategorien:- Ausgestorbenes Säugetier
- Rüsseltiere
- Fauna des Pleistozäns


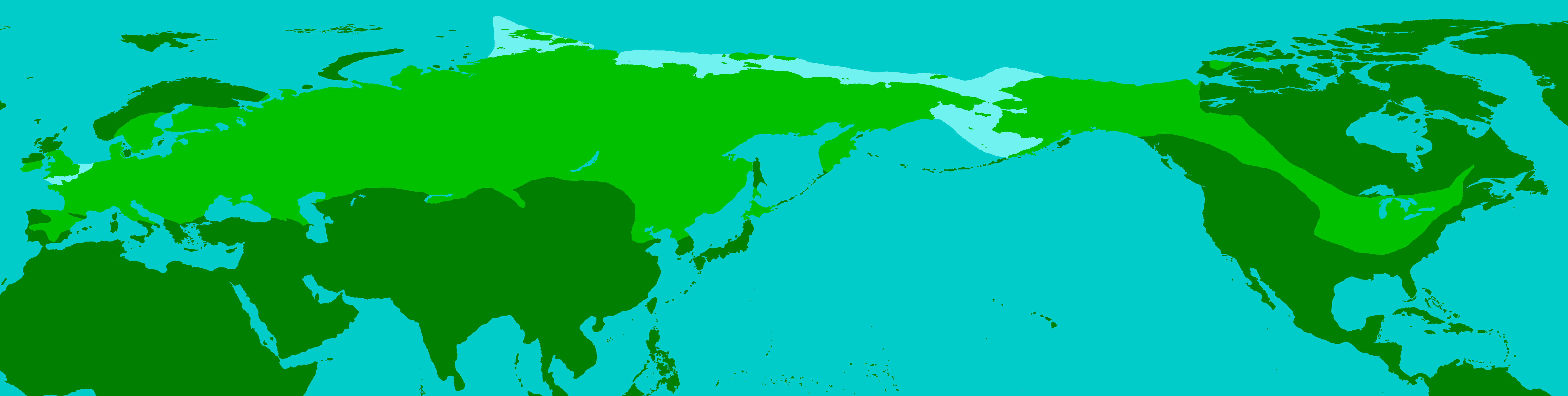




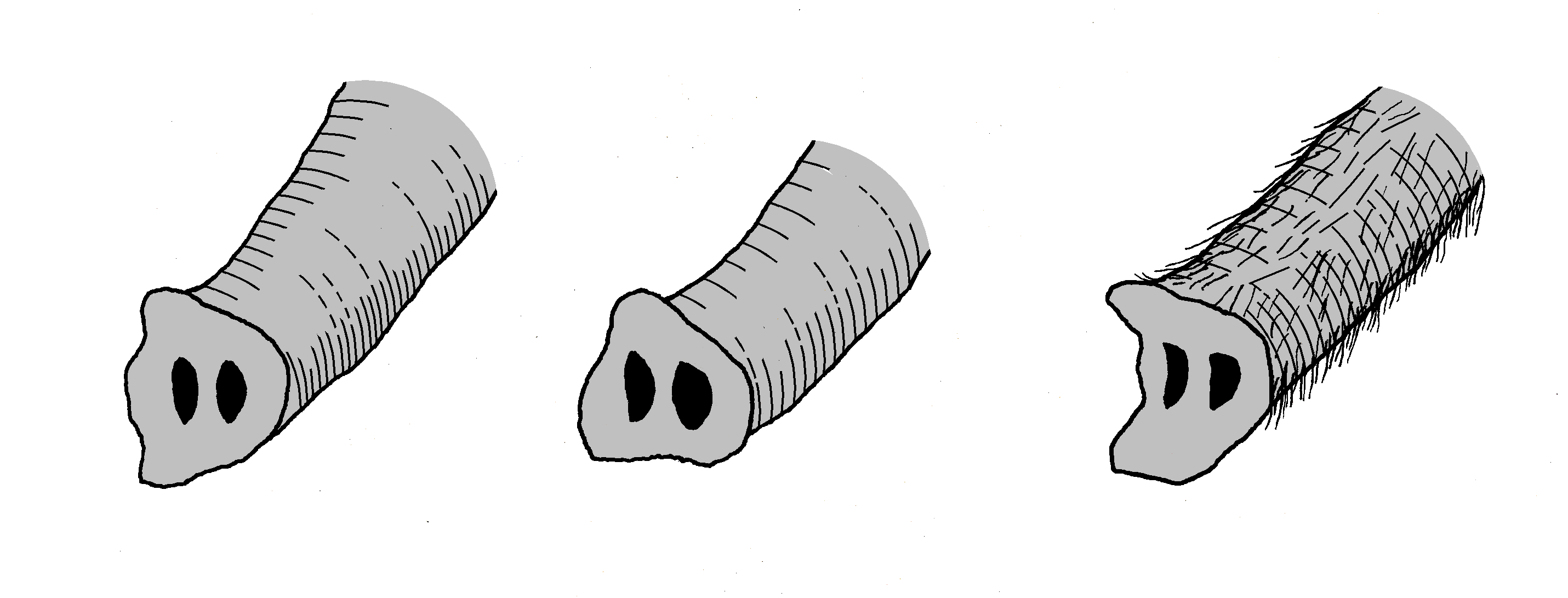






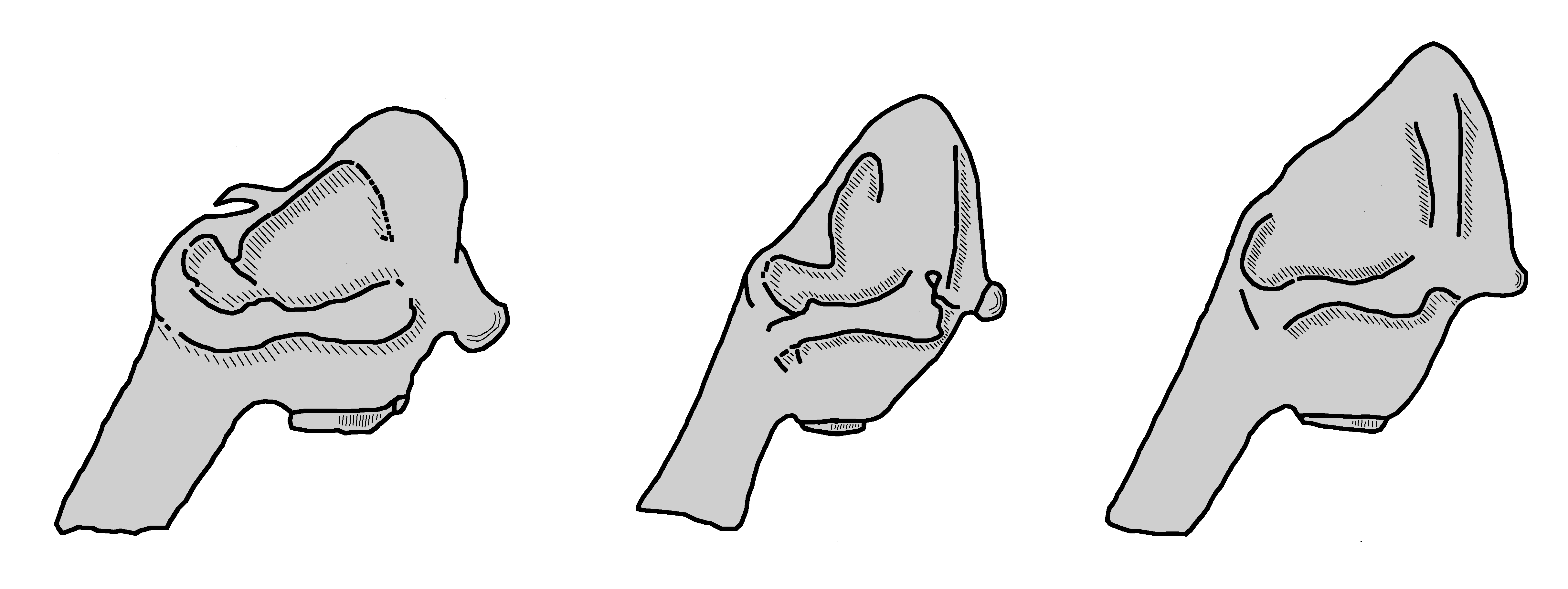
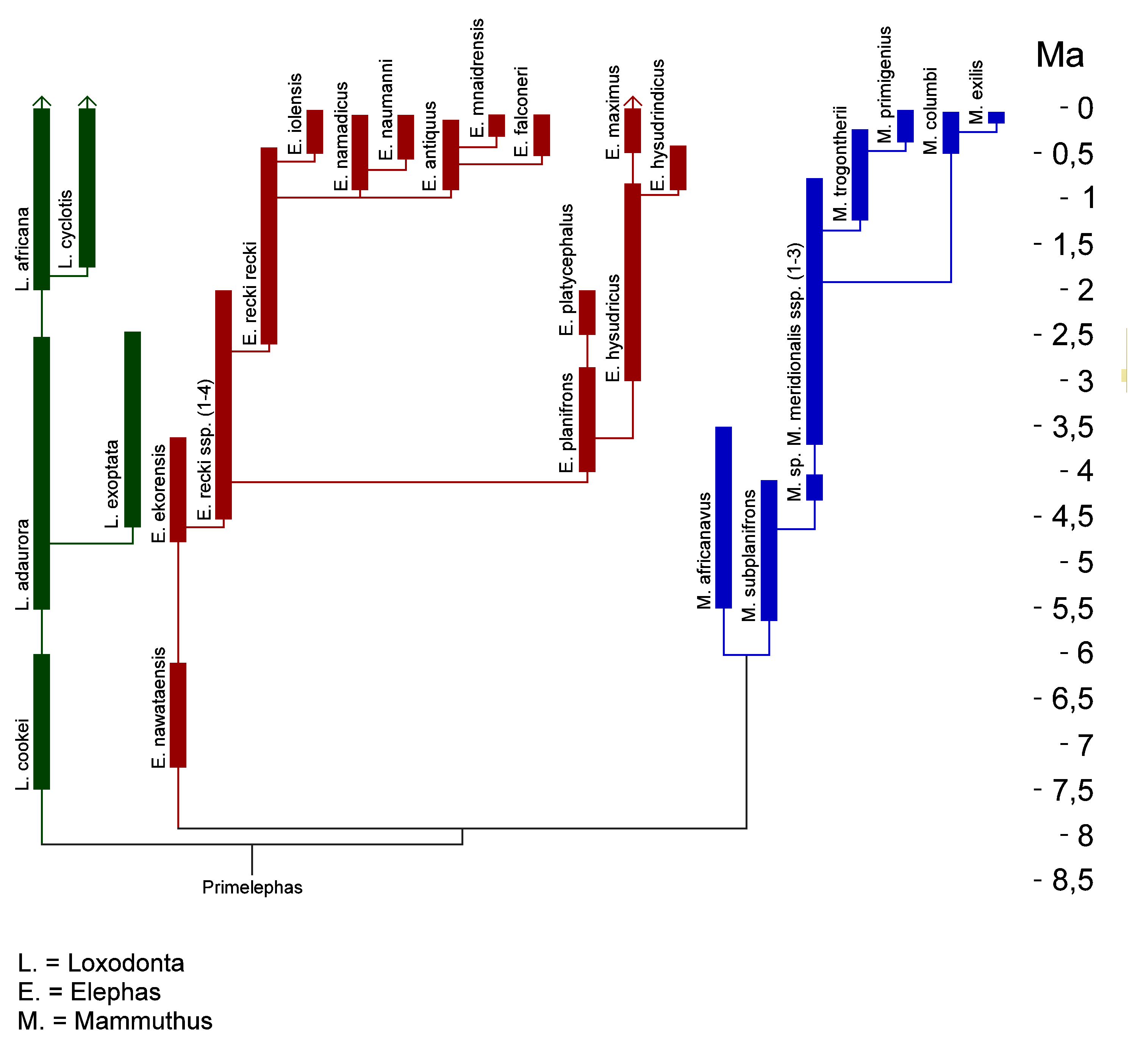

Wikimedia Foundation.