- Nervatur
-
Das Blatt ist neben der Sprossachse und der Wurzel eines der drei Grundorgane der höheren Pflanzen und wird als Organtyp Phyllom genannt. Blätter sind seitliche Auswüchse an den Knoten (Nodi) der Sprossachse. Die ursprünglichen Funktionen der Blätter sind Photosynthese (Aufbau organischer Stoffe mit Hilfe von Licht) und Transpiration (Wasserverdunstung, wichtig für Nährstoffaufnahme und -transport).
Blätter treten nur bei Sprosspflanzen auf, das heißt bei farnartigen Pflanzen (Pteridophyta) und Samenpflanzen (Spermatophyta). Dagegen fehlen sie bei Moosen und Algen, an deren Thallus allerdings blattähnliche Gebilde auftreten können, die jedoch nur als Analogien der Blätter zu betrachten sind.
Der Reichtum an Blattformen ist enorm. In einigen Fällen entstanden im Laufe der Evolution auch Blattorgane, die mit der ursprünglichen Funktion des Blattes, nämlich der Photosynthese und Transpiration, nichts mehr zu tun haben: zum Beispiel Blütenblätter, Blattdornen und Blattranken, sowie Knospenschuppen (siehe Metamorphosen des Blattes).
 Nadelblätter einer Douglasie (Pseudotsuga menziesii)
Nadelblätter einer Douglasie (Pseudotsuga menziesii) Laubblatt einer Linde (Tilia spec.)
Laubblatt einer Linde (Tilia spec.)Inhaltsverzeichnis
Anatomie
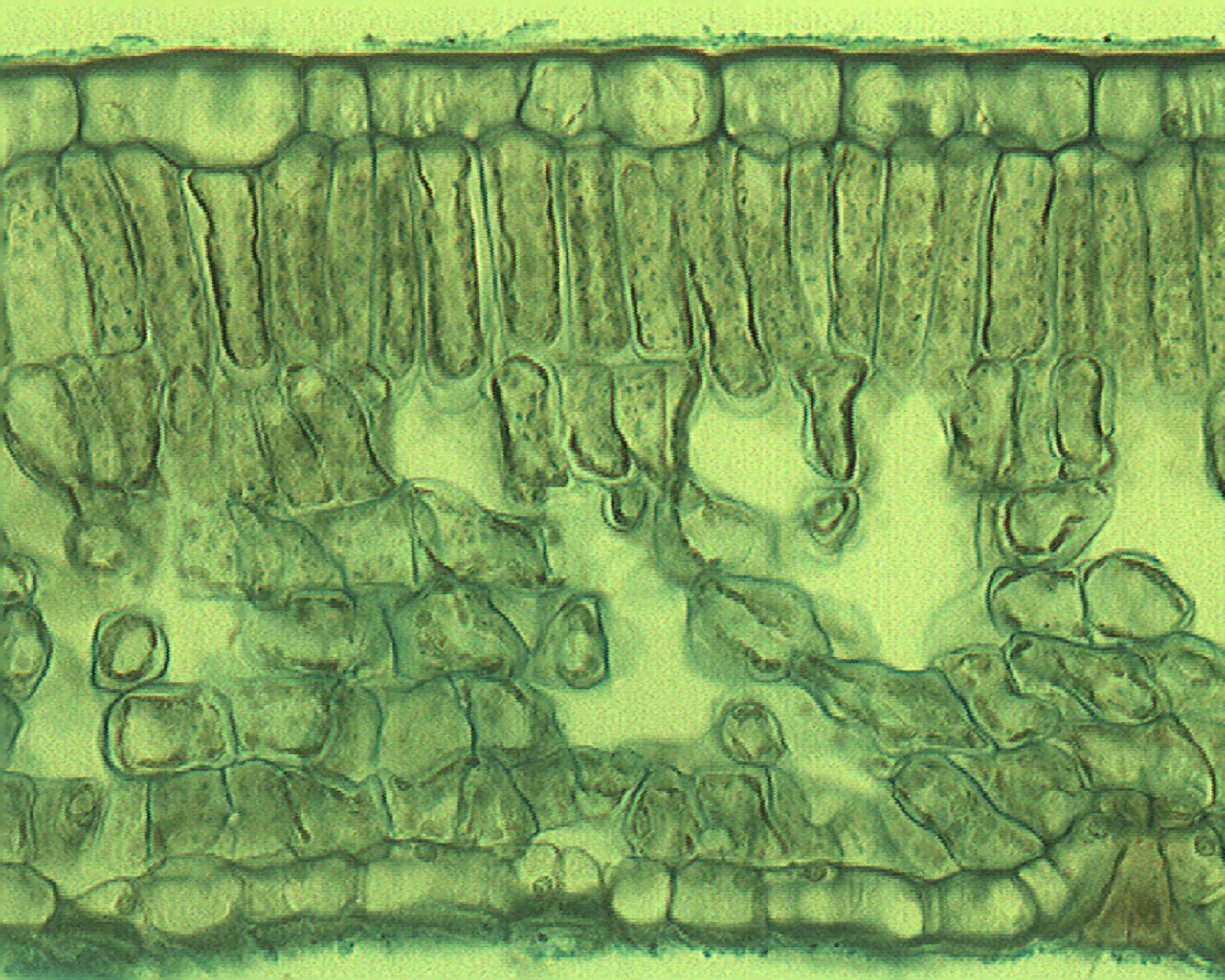
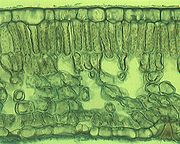 Querschnitt eines Laubblattes im Mikroskop (Foto: Philip Frank, Alicante)
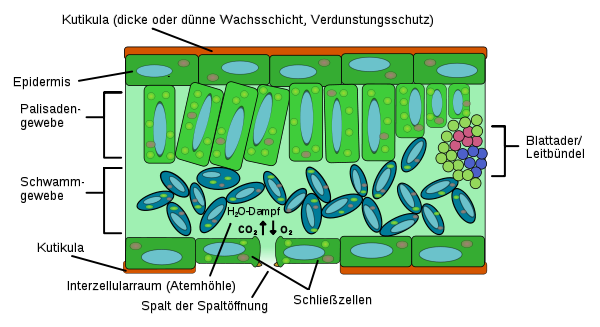
Querschnitt eines Laubblattes im Mikroskop (Foto: Philip Frank, Alicante)Der hier beschriebene anatomische Aufbau gilt für ein bifaziales Laubblatt, den häufigsten Laubblatt-Typ. Für alle Blätter charakteristisch sind die Elemente Epidermis, Mesophyll und Leitbündel.
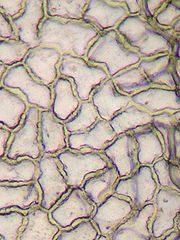
Epidermis
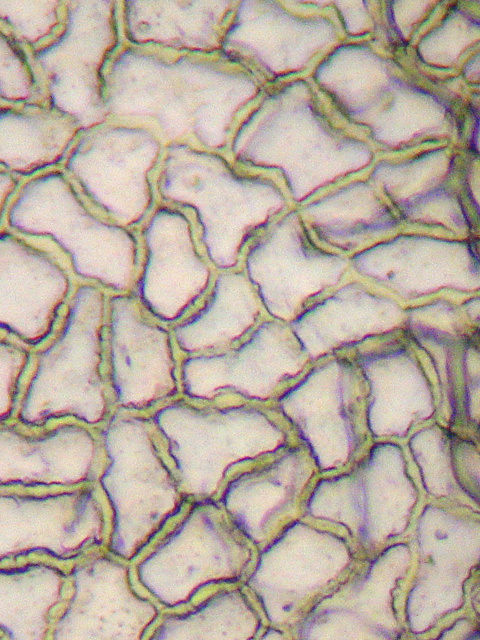
 Epidermiszellen
EpidermiszellenDas Blatt schließt nach außen mit einem Abschlussgewebe, der Epidermis, ab, die aus nur einer Zellschicht besteht. Die Epidermis besitzt nach außen eine wasserundurchlässige Wachsschicht Cuticula, die eine unregulierte Verdunstung verhindert. Die Zellen der Epidermis besitzen in der Regel keine Chloroplasten (die Zellbestandteile, in denen die Photosynthese stattfindet). Ausnahmen davon sind die Epidermis von Hygro-, Helo- und Hydrophyten und teilweise Schattenblätter, besonders aber die Schließzellen der Spaltöffnungen (Stomata), die immer Chloroplasten enthalten. Die Stomata dienen der Regulation des Gasaustausches, primär der Wasserdampfabgabe. Nach der Verteilung der Stomata unterscheidet man hypostomatische (Stomata auf der Blattunterseite, häufigste Form), amphistomatische (Stomata auf beiden Blattseiten) und epistomatische Blätter (Stomata auf der Blattoberseite, z. B. bei Schwimmblättern).
Die von der Epidermis gebildeten Anhänge werden Haare (Trichome) genannt. Sind an der Bildung auch subepidermale Zellschichten beteiligt, spricht man von Emergenzen: Beispiele sind Stacheln oder Drüsenzotten.
Mesophyll (Blattparenchym)
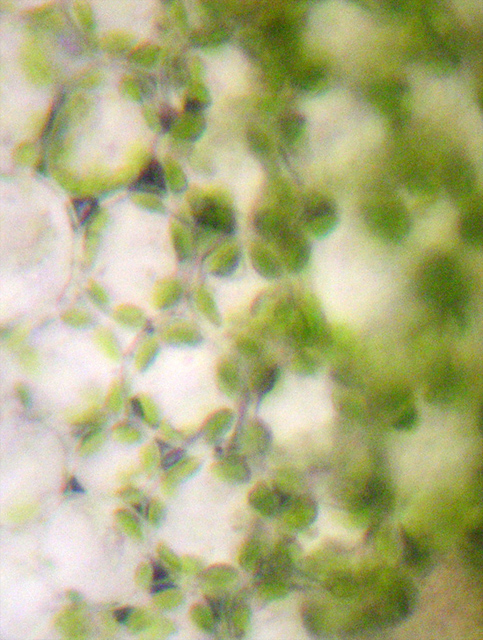
 Palisadenparenchym (Querschnitt)
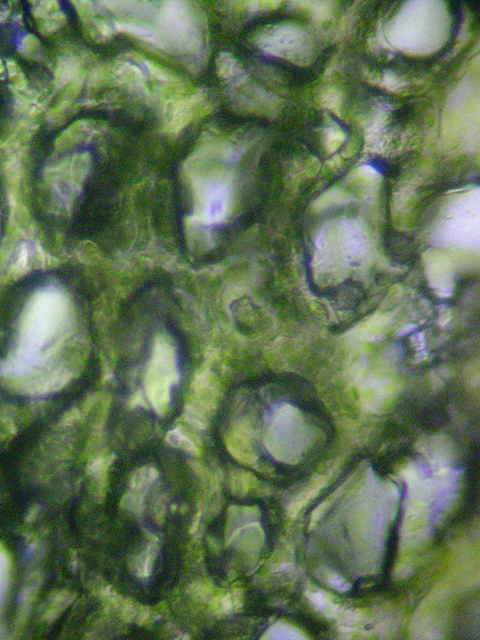
Palisadenparenchym (Querschnitt)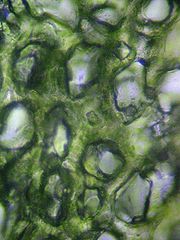 Schwammparenchym
SchwammparenchymAls Mesophyll bezeichnet man das Assimilationsgewebe. Es ist meist in das unter der oberen Epidermis gelegene Palisadenparenchym und das darunter gelegene Schwammparenchym gegliedert. Das Palisadenparenchym besteht aus ein bis drei Lagen langgestreckter, senkrecht zur Blattoberfläche stehender, chloroplastenreicher Zellen. Im Palisadenparenchym, dessen Hauptaufgabe die Photosynthese ist, befinden sich rund 80 Prozent aller Chloroplasten. Das Schwammparenchym besteht aus unregelmäßig geformten Zellen, die aufgrund ihrer Form große Interzellularräume bilden. Die Hauptaufgabe des Schwammparenchyms ist es, die Durchlüftung des parenchymatischen Gewebes zu gewährleisten. Die Zellen sind relativ arm an Chloroplasten.
Leitbündel
Die Leitbündel befinden sich oft an der Grenze zwischen Palisaden- und Schwammparenchym im oberen Schwammparenchym. Der Aufbau gleicht dem der Leitbündel in der Sprossachse und ist meist kollateral. Die Leitbündel zweigen von der Sprossachse ab und gehen durch den Blattstiel ohne Drehung in die Spreite über. Dadurch weist das Xylem zur Blattoberseite, das Phloem zur Blattunterseite.
Große Leitbündel sind oft von einer Endodermis umgeben, die hier Bündelscheide genannt wird. Die Bündelscheide kontrolliert den Stoffaustausch zwischen Leitbündel und Mesophyll. Die Leitbündel enden blind im Mesophyll. Dabei wird das Leitbündel immer stärker reduziert, das heißt zunächst werden die Siebröhren weniger und fallen aus, dann verbleiben im Xylem-Teil nur Schraubentracheiden, die schließlich blind enden. Das gesamte Blatt ist in der Regel so dicht mit Leitbündeln durchzogen, dass keine Blattzelle weiter als sieben Zellen von einem Leitbündel entfernt ist. Die sich daraus ergebenden kleinen Felder zwischen den Leitbündeln heißen Areolen oder Interkostalfelder.
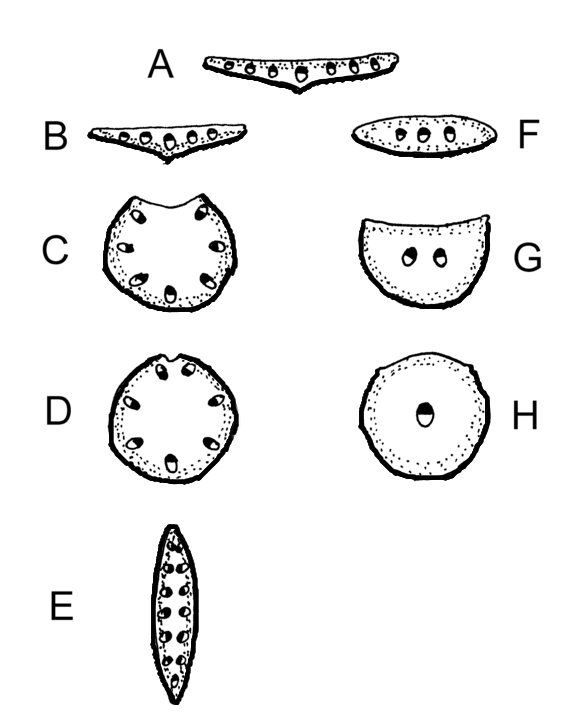
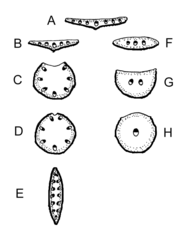 Blatt-Typen im Querschnitt: Punktiert = Palisadenparenchym; Holzteile der Leitbündel sind schwarz; Blattunterseite dicke Linie; A = Normales bifaziales Blatt; B = invers bifaziales Blatt (Bärlauch); C,D Ableitung des unifazialen Rundblattes (Allium sativum, Juncus effusus); E = unifaziales Schwertblatt (Schwertlilien); F = äquifaziales Flachblatt; G = äquifaziales Nadelblatt; H = Äquifaziales Rundblatt (Sedum).
Blatt-Typen im Querschnitt: Punktiert = Palisadenparenchym; Holzteile der Leitbündel sind schwarz; Blattunterseite dicke Linie; A = Normales bifaziales Blatt; B = invers bifaziales Blatt (Bärlauch); C,D Ableitung des unifazialen Rundblattes (Allium sativum, Juncus effusus); E = unifaziales Schwertblatt (Schwertlilien); F = äquifaziales Flachblatt; G = äquifaziales Nadelblatt; H = Äquifaziales Rundblatt (Sedum).Die Funktion der Leitbündel ist der Antransport von Wasser und Mineralien ins Blatt (über das Xylem) sowie der Abtransport von Photosyntheseprodukten aus dem Blatt (über das Phloem).
Festigungsgewebe
In der Nähe der Leitbündel oder auch an den Blatträndern befinden sich oft Sklerenchymstränge, die der Festigung des Blattgewebes dienen. Demselben Zweck dienen bei manchen Arten subepidermale Kollenchymschichten.
Einteilung nach anatomischen Gesichtspunkten
Nach der Lage des Palisadenparenchyms im Blatt werden verschiedene Blatt-Typen unterschieden.
- Die meisten Blätter sind bifazial gebaut, d. h. es wird eine Ober- und Unterseite ausgebildet.
- Bei normal bifazialen (= dorsiventralen) Blättern (A) liegt das Palisadenparenchym oben (= dorsal), das Schwammgewebe unten (= ventral).
- Bei invers bifazialen Blättern (B) liegt das Palisadenparenchym unten (z. B. beim Bärlauch).
- Bei äquifazialen Blättern (F, G) sind Ober- und Unterseite gleich mit Palisadenparenchym versehen, dazwischen liegt das Schwammparenchym. Ein typisches Beispiel ist das Nadelblatt der Kieferngewächse (G).
- Bei unifazialen Blättern (C, D) geht die Ober- und Unterseite nur aus der Unterseite des Blattprimordiums hervor. Sie leiten sich formal von invers bifazialen Blättern ab, bei denen die Blattoberseite reduziert wird. Bei unifazialen Blättern liegen die Leitbündel im Blattquerschnitt in einem Kreis oder Bogen angeordnet, das Phloem zeigt nach außen. Blattstiele sind oft unifazial, aber auch die Blätter vieler Einkeimblättriger, wie etwa Binsen, deren Blätter oft sprossachsenähnlich sind. Ein Spezialfall sind die Blätter der Schwertlilien (E), deren unifaziales Blatt sekundär wieder flach wurde, aber durch Abflachung in der Achsenrichtung, sodass reitende Blätter, auch Schwertblätter genannt, entstanden.
Morphologische Gliederung
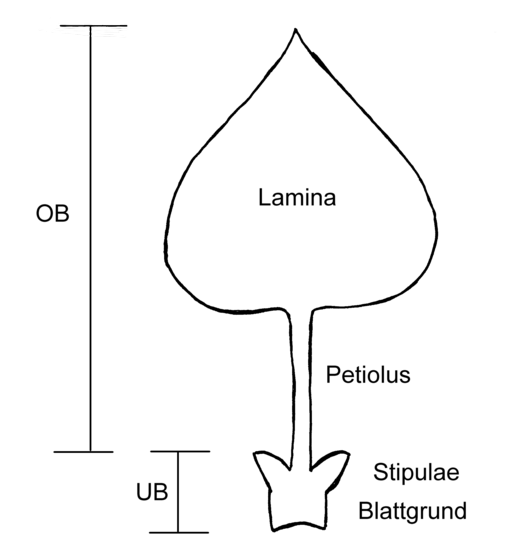
 Gliederung des Blattes. OB = Oberblatt, UB = Unterblatt, Lamina = Spreite, Petiolus = Stiel, Stipulae = Nebenblätter
Gliederung des Blattes. OB = Oberblatt, UB = Unterblatt, Lamina = Spreite, Petiolus = Stiel, Stipulae = NebenblätterEin Blatt ist unterteilt in das Unterblatt (Hypophyll), bestehend aus dem Blattgrund und den Nebenblättern (Stipulae), und in das Oberblatt (Epiphyll), das sich wieder in Blattspreite (Lamina) und Blattstiel (Petiolus) gliedert. Nicht bei allen Blättern sind alle Teile ausgebildet, alle Teile unterliegen einer mannigfachen Variation.
- Zur Beschreibung der Blattform in der botanischen Literatur siehe den Hauptartikel Blattform
Unterblatt
Blattgrund
Der Blattgrund oder die Blattbasis ist der unterste Teil, mit dem das Blatt der Sprossachse ansitzt. Als Blattachsel bezeichnet man den Winkel zwischen Sprossachse und davon abzweigendem Blatt. Er ist meist nur wenig verdickt, nimmt aber manchmal den ganzen Umfang der Sprossachse ein. Im letzteren Fall spricht man von einem stängelumfassenden Blatt. Bei gegenständiger Blattstellung sind bisweilen die Basen der beiden Blätter vereinigt (wie beispielsweise bei der Heckenkirsche). Bisweilen zieht der Blattgrund beiderseits als ein flügelartiger Streifen weit am Stängel herab; solche Stängel nennt man geflügelt.
Bei einigen Pflanzenfamilien, etwa bei Süß- und Sauergräsern und Doldengewächsen, bildet der Blattgrund eine so genannte Blattscheide aus. Es handelt sich dabei um einen mehr oder weniger breiten, meist über der Basis des Blattes zu findenden, scheidenartig die Sprossachse umschließenden Teil. Meistens ist dabei die Scheide gespalten, d. h. die Ränder sind frei, nur übereinander gelegt. Dagegen haben die Blätter der Sauergräser geschlossene Scheiden oder solche, an denen keine freien Ränder vorhanden sind. Bei vielen Blättern aber ist der Scheidenteil nur angedeutet oder fehlt ganz.
Nebenblätter
 Bei der Echten Nelkenwurz sind die Nebenblätter laubblattförmig.
Bei der Echten Nelkenwurz sind die Nebenblätter laubblattförmig.Die Nebenblätter (Stipulae oder Stipeln) sind seitliche, zipfel- oder blattartige Auswüchse des Blattgrundes. Sie sind meist klein, bei vielen Pflanzenarten fehlen sie oder werden bereits beim Blattaustrieb abgeworfen. Je nach Bau des Blattstieles treten zwei Arten auf. Bei bifazialem Blattstiel treten Lateralstipeln auf, die stets paarig seitlich am Blattgrund sitzen. Diese Form ist charakteristisch für Zweikeimblättrige. Bei unifazialem Blattstiel treten Median- (Axillar-)stipeln auf, die nur in Einzahl auftreten und in der Mediane in der Achsel des Blattes liegen. Sie sind häufig kapuzenförmig und treten vor allem bei Einkeimblättrigen auf.
Bei einigen Familien sind die Nebenblätter stark entwickelt, so bei den Schmetterlingsblütlern (wie der Erbse), den Rosengewächsen und den Veilchengewächsen. Sie können entweder frei (z. B. Wicken) oder scheinbar dem Blattstiel angewachsen sein (Rosen).
Bei etlichen Bäumen, wie Linden, Hainbuchen oder Pappeln sind die Nebenblätter als häutige, nicht grüne Schuppen ausgebildet, die schon während der Entfaltung der Blätter abfallen. Bei den Knöterichgewächsen sind die Nebenblätter zu einer Nebenblattscheide (Ochrea) umgebildet, einer häutigen Scheide, die den Stängel röhrenförmig einschließt. Das Blatthäutchen (Ligula) der Süß- und Sauergräser, das am Übergang von der Blattscheide in die Blattspreite sitzt, ist ebenfalls ein Nebenblatt.
Oberblatt
Blattstiel
Der Blattstiel (Petiolus) ist der auf den Blattgrund folgende, durch seine schmale, stielförmige Gestalt vom folgenden Teil des Blattes mehr oder minder scharf abgegrenzte Teil des Blattes. Nach dem anatomischen Aufbau unterscheidet man bifaziale und unifaziale Blattstiele. Bei den meisten Einkeimblättrigen und bei vielen Koniferen fehlt der Blattstiel. Blätter ohne Stiel nennt man sitzend. Es gibt auch Blätter, die nur aus dem Stiel bestehen, der dann flach und breit ist und an welchem die eigentliche Blattfläche ganz fehlt. Es handelt sich dabei um ein so genanntes Blattstielblatt (Phyllodium), z. B. bei manchen Akazien. Der Blattstiel ist meist nur bei Laubblättern ausgebildet.
Blattspreite, „Blattnervatur“
Die Blattspreite (Lamina) bildet in den meisten Fällen den Hauptteil des Blattes, den man oft als das eigentliche Blatt bezeichnet. Die Blattspreite ist im Normalfall die Trägerin der Blattfunktionen Photosynthese und Transpiration.
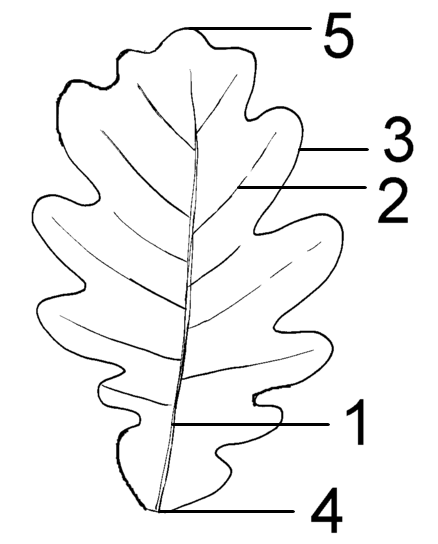
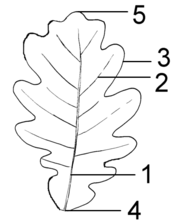 Teile der Spreite: 1 Mittelrippe, 2 Seitenrippe, 3 Blattrand, 4 Spreitengrund, 5 Spreitenspitze
Teile der Spreite: 1 Mittelrippe, 2 Seitenrippe, 3 Blattrand, 4 Spreitengrund, 5 SpreitenspitzeAn den meisten Blattspreiten fällt die sogenannte Nervatur auf, der Verlauf der Leitbündel. Große Leitbündel werden auch Rippen genannt, viele Blätter besitzen eine Mittelrippe (1) als scheinbare Verlängerung des Blattstieles, von der die Seitenrippen (2) abzweigen. Die Leitbündel werden volkstümlich meist als Nerven oder Adern bezeichnet, beides missverständliche Begriffe, da die Leitbündel weder eine Erregungsleitungs- noch eine Kreislauffunktion besitzen.
Es werden drei Formen von Nervatur unterschieden, die auch eine systematische Bedeutung haben. Bei den Einkeimblättrigen tritt hauptsächlich Parallelnervatur auf. Hier verlaufen die Hauptadern längs und parallel zueinander. Daraus ergibt sich der meist glatte Blattrand der Einkeimblättrigen. Besonders deutlich wird dies bei den Gräsern. Die Hauptadern und auch die vielen kleineren Parallel-Leitbündel sind jedoch durch kleine, meist mit freiem Auge sichtbare Leitbündel miteinander verbunden (transversale Anastomosen). Die parallele Anordnung der Leitbündel führt auch zu einer parallelen Anordnung der Spaltöffnungen.
Die meisten Zweikeimblättrigen besitzen eine kompliziertere Netznervatur. Daraus ergibt sich auch die fast beliebige Form der Spreite.
Bei Farnen und beim Ginkgo tritt die Gabel- oder Fächernervatur auf. Hier sind die Leitbündel dichotom (gabelförmig) verzweigt und enden blind am vorderen Blattende.
Besonders bei den Zweikeimblättrigen treten die Laubblätter in einer großen Formenvielfalt auf. Die Form und Beschaffenheit der Blätter sind daher wichtige Bestimmungsmerkmale zum Erkennen der Pflanzenarten. Die Beschaffenheit kann z. B. häutig, ledrig oder sukkulent (=fleischig) sein. Für die Oberfläche sind häufig auch Haare (Trichome) von Bedeutung. Bei der Gestalt sind wichtig:
- Die Gliederung der Blattspreite: Wenn die Spreite eine einzige zusammenhängende Gewebefläche darstellt, spricht man von einem „einfachen“ Blatt. Im Unterschied dazu gibt es auch so genannte „zusammengesetzte“ Blätter. Bei ihnen ist die Aufteilung der Blattfläche so weit fortgeschritten, dass die einzelnen Abschnitte als vollständig voneinander geschiedene Teile erscheinen. Diese werden – unabhängig von ihrer Größe – als Blättchen bezeichnet. Sie ahmen die Gestalt einfacher Blätter nach und sind häufig sogar mit einem Blattstielchen versehen.
- Die Anordnung der Abschnitte: Nach ihrer gegenseitigen Anordnung lassen sich grob drei Typen unterscheiden:
- gefiederte Blätter,
- handförmige Blätter und
- fußförmige Blätter.
Bei den ersteren heißt die Mittelrippe, d. h. der gemeinschaftliche Stiel, an welchem die einzelnen Fiederblättchen meist in Paaren sitzen, Blattspindel (Rhachis). Schließt letztere mit einem Endblättchen (Endfieder) ab, hat man ein unpaarig gefiedertes Blatt vor sich. Das endständige Fiederblättchen kann auch rankenförmig umgebildet sein wie z. B. bei den Erbsen. Dagegen spricht man von einem paarig gefiederten Blatt, wenn ein solches Endblättchen fehlt. Die handförmigen Blätter unterscheidet man nach der Anzahl der Teilblättchen als dreizählig, fünfzählig etc. Es gibt auch Blätter, die mehrfach zusammengesetzt sind; dies ist besonders häufig bei gefiederten Blättern der Fall. Die Abschnitte werden hier Fiedern genannt. Man spricht hier von „doppelt gefiederten“ Blättern.

Einfaches, ungeteiltes Blatt der Zitterpappel

Gefiedertes Blatt der Rose

Handförmiges Blatt der Rosskastanie

Fußförmiges Blatt der Schneerose
- Der Blattrand (3): Die sehr mannigfaltigen Formen des Blattrandes werden in der Botanik durch zahlreiche Begriffe bezeichnet, von denen nachfolgend einige aufgelistet sind: ganzrandig, gezähnt, gesägt, gebuchtet, gekerbt usw.
- Die Gestalt der Spreite oder Blättchen: Hier wird angegeben, ob das Blatt z. B. rundlich, elliptisch, linealisch, nierenförmig usw. ist.
- Der Spreitengrund (4), auch Spreitenbasis genannt, beschreibt, wie die Blattspreite in den Blattstiel übergeht: z. B. herzförmig, pfeilförmig.
- Der Spreiten-Apex (5, die Spitze) kann ausgerandet, abgerundet, spitz, stumpf usw. sein.
- Von Bedeutung ist auch der Spreitenquerschnitt (umgerollt, gefaltet, gerillt).
- Auch die dreidimensionale Form kann vom typischen Blatt abweichen (kugelig, röhrenförmig usw.)
Eine detaillierte Beschreibung der Blattformen wird im Artikel Blattform aufgezeigt.
Evolution
Man unterscheidet generell zwei Typen von Blättern, die gemäß der Telomtheorie unabhängig voneinander entstanden sind:
- Mikrophylle sind kleine, oft nadelförmige Blätter mit nur einem Leitbündel. Das Mesophyll ist meist wenig differenziert. Ihre Entstehung in der Evolution deutet man als Reduktion der Telome. Die ältesten Gefäßpflanzen, die ab dem Obersilur bekannten Urfarngewächse wie Cooksonia und Rhynia hatten noch keine Blätter. Die ersten Mikrophylle sind von den Protolepidodendrales aus dem Unterdevon bekannt. Heute kommen die Mikrophylle bei den Bärlapppflanzen, den Schachtelhalmen und den Gabelblattgewächsen vor. Mikrophylle sind in der Regel klein, bei den Schuppenbäumen (Lepidodendron) erreichten sie jedoch eine Länge von rund einem Meter.
- Die Entstehung der Makro- oder Megaphylle wird durch die Einebnung (Planation) und anschließende Verwachsung der ursprünglich dreidimensional angeordneten Telome erklärt. Megaphylle treten erstmals bei den Farnen (Polypodiophyta) auf und werden hier meist Wedel genannt. Der Grundtyp des Megaphylls ist das gefiederte Laubblatt. Die übrigen Blattformen lassen sich – weitgehend auch fossil belegt – davon ableiten. Bei den fossilen Primofilices (Mitteldevon bis Unterperm) waren die Fiederabschnitte noch räumlich angeordnet (Raumwedel), wie auch heute noch bei den Natternzungengewächsen (Ophioglossaceae).[1]
Wachstum und Lebensdauer
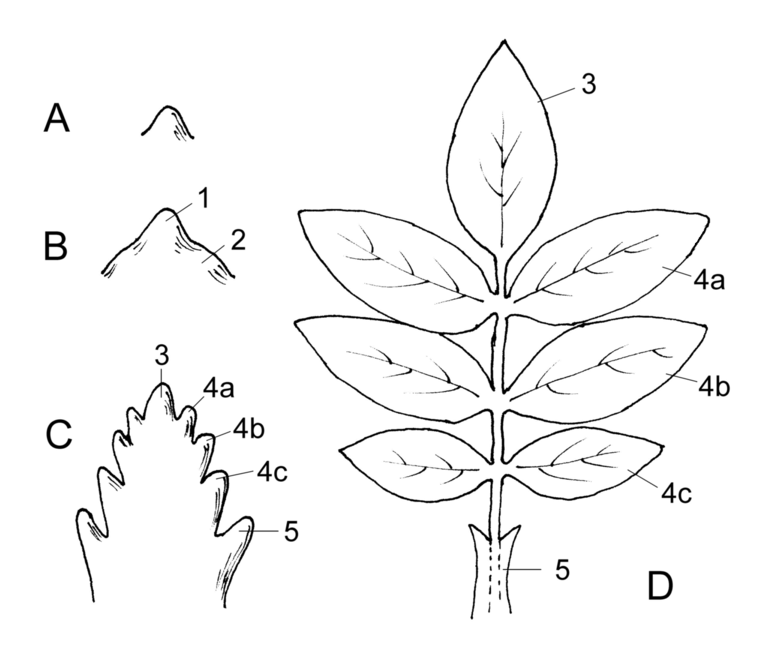
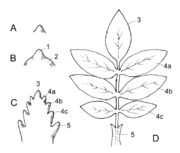 Entwicklung eines Fiederblattes. A Blatthöcker am Sprossscheitel, B Gliederung in Oberblatt (1) und Unterblatt (2), C Anlage der Fiederblätter, D fertiges Fiederblatt. 3 Endfieder, 4a,b,c Seitenfiedern, 5 Nebenblatt
Entwicklung eines Fiederblattes. A Blatthöcker am Sprossscheitel, B Gliederung in Oberblatt (1) und Unterblatt (2), C Anlage der Fiederblätter, D fertiges Fiederblatt. 3 Endfieder, 4a,b,c Seitenfiedern, 5 NebenblattBlätter entstehen aus wenigen Zellen aus den äußeren Zellschichten (Tunica) des Sprossmeristems, also exogen. Unterhalb des Apikalmeristems bilden sich in der Tunica seitliche Auswüchse. Aus einer zunächst schwachen Erhebung entsteht ein kleiner, meist stumpf konischer Zellgewebshöcker, das Blattprimordium oder die Blattanlage genannt.
Durch ein Signal des Sprossmeristems erfolgt die dorso-ventrale Organisation des Blattes. Unterbleibt dieses Signal – etwa indem das Blattprimordium vom Sprossmeristem getrennt wird – bildet sich eine radiärsymmetrische Struktur mit ventralen Differenzierungen. Die dorsale Entwicklung wird durch eine Gengruppe gefördert, zu der die Gene PHABULOSA (PHB), PHAVOLUTA (PHV) und REVOLUTA (REV) gehören, die für Transkriptionsfaktoren kodieren. Diese Gene werden schon in der Peripheren Zone des Sprossmeristems gebildet, also noch vor der Bildung des Blattprimordiums. Sobald das Primordium erkennbar ist, ist die Expression der Gene auf die dorsale Seite beschränkt. Auf der ventralen Seite des Blattprimordiums werden Gene der YABBY (YAB) Genfamilie (Transkriptionsfaktoren mit Zinkfinger-Domäne) und Gene der KANADI (KAN) Genfamilie (GARP Transkriptionsfaktoren) exprimiert. Auch diese Gene werden zunächst gleichmäßig im ganzen Blattprimordium exprimiert. Blattanlagen exprimieren also zunächst dorsalisierende (PHB) wie auch ventralisierende (YAB, KAN) Gene. Ein Signal vom Meristem aktiviert PHB Transkriptionsfaktoren, abhängig von der Lage reprimieren diese die YAB und KAN Gene und erhalten die eigene Expression aufrecht. Auf diese Weise entsteht die dorso-ventrale Gliederung. Auch die proximo-distale Blattentwicklung scheint dadurch gefördert zu werden.[2]
 Ein Kirschblatt in Herbstfärbung. Deutlich zu erkennen die Mittelrippe und die Seitenrippen, sowie die kleineren, netzartig verbundenen Leitbündel.
Ein Kirschblatt in Herbstfärbung. Deutlich zu erkennen die Mittelrippe und die Seitenrippen, sowie die kleineren, netzartig verbundenen Leitbündel.Aus der Blattanlage entwickelt sich der Blatthöcker, dieser differenziert sich durch eine Einschnürung in einen breiten, proximalen Abschnitt, das Unterblatt, und einen schmalen, distalen Abschnitt, das Oberblatt.
Das Wachstum erfolgt nur kurze Zeit mit der Spitze (akroplast). Die Spitze stellt sehr früh ihr Wachstum ein, das Wachstum erfolgt durch basale oder interkalare Meristeme (basiplastes bzw. interkalares Wachstum). Die Blattspreite (Lamina) entsteht meist durch basiplastes Wachstum, der Blattstiel (Petiolus) und die Spreiten der Gräser durch interkalares Wachstum. Eine Ausnahme bilden die Farne, deren Wachstum akroplast mittels einer Scheitelzelle bzw. einer Scheitelkante (aus mehreren Zellen) erfolgt.
Im weiteren Wachstumsverlauf passieren Zellteilungs- und Zellstreckungsvorgänge nicht im gesamten Blattkörper gleichmäßig, sondern nur innerhalb meristematisch (bzw. teilungs-) aktiver Zonen. Ob, zu welchem Zeitpunkt, und wie intensiv diese Zonen aktiv sind, ist genetisch festgelegt und führt zu einer charakteristischen Blattform.
Blätter haben in der Regel nur eine begrenzte Lebensdauer, nur bei wenigen mehrjährigen Arten bleiben die Blätter während der ganzen Lebensdauer der Pflanze erhalten (z. B. bei der Welwitschie). Nach der Lebensdauer unterscheidet man zwischen immergrünen Blättern (leben mindestens zwei Vegetationsperioden), wintergrünen (überwintern grün), sommergrünen (nur eine Vegetationsperiode lang) und hinfälligen Blättern (fallen sehr bald ab, z. B. Kelchblätter des Mohns).
Der Blattfall erfolgt durch Bildung einer eigenen Trennungszone (Abszissionszone) am Übergang von der Sprossachse zum Blatt (siehe Abszission).
Farbe und Farbänderung
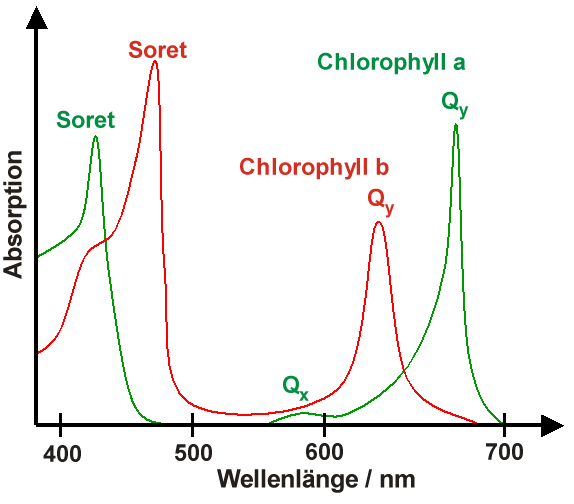
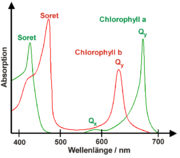 Absorptionsspektrum von Chlorophyll a und b
Absorptionsspektrum von Chlorophyll a und bDie Absorptionsspektren von in Lösungsmitteln gelösten Chlorophyllen besitzen immer zwei ausgeprägte Absorptionsmaxima, eines zwischen 600 und 800 nm und eines um 400 nm, das Soret-Bande genannt wird. Die Abbildung rechts zeigt diese Absorptionsmaxima für Chlorophyll a und b. Anhand dieser Spektren kann man leicht verstehen, warum Blätter – diese enthalten Chlorophyll a und b – grün sind. Zusammen absorbieren Chlorophyll a und b hauptsächlich im blauen Spektralbereich (400–500 nm) sowie im roten Spektralbereich (600–700 nm). Im grünen Bereich hingegen findet keine Absorption statt, so dass dieser Anteil von Sonnenlicht gestreut wird, was Blätter grün erscheinen lässt.
Besonders auffällig ist die Blattverfärbung vor dem herbstlichen Laubfall. Dieser kommt dadurch zustande, dass in den Zellen das grüne Stickstoff-reiche Photosynthese-Pigment Chlorophyll abgebaut und der Stickstoff in die Sprossachse verlagert wird. Im Blatt verbleiben die bis dahin vom Grün überdeckten gelben Carotine und bei manchen Arten die roten Anthocyane, die für die bunte Herbstfärbung verantwortlich sind. Bei manchen Pflanzen dominieren die Anthocyane generell über das grüne Chlorophyll, so z. B. bei der Blutbuche. Andere Blätter sind grün-weiß gefleckt, panaschiert. Diese Formen sind im Zierpflanzenbereich sehr beliebt.
Blattfolge
Als Blattfolge bezeichnet man die Abfolge verschieden gestalteter Blätter an einer Pflanze. Eine typische Blattfolge ist Keimblätter – Primärblätter – Laubblätter – Blütenblätter. Dazwischen können noch Hoch- und Niederblätter zwischengeschaltet sein. Bei den Farnen bleibt, mit unbedeutenden Ausnahmen, die Gestalt der Blätter am gesamten Spross und an allen Zweigen ziemlich unverändert. Im Zuge der Blattfolge kann es zur so genannten Heterophyllie kommen, der unterschiedlichen Ausformung von Blättern an einer Pflanze. Heterophyllie findet sich z. B. am Efeu.
Keimblätter
 Keimblätter von Jacaranda mimosifolia (Palisanderbaum)
Keimblätter von Jacaranda mimosifolia (Palisanderbaum)Die Keimblätter (Kotyledonen) der Samenpflanzen sind die ersten, im Embryo angelegten Blätter und bereits im Samen erkennbar. Sie sind meist wesentlich einfacher gestaltet als die folgenden Blätter. Die Anzahl der Keimblätter dient auch als ein wichtiges systematisches Merkmal. Die Klasse der Einkeimblättrigen (Liliopsida) wurde nach ihrem einzigen Keimblatt benannt (monokotyl). Ihnen wurde bis vor wenigen Jahren die Klasse der Zweikeimblättrigen (Magnoliopsida) gegenübergestellt (dikotyl), die heute jedoch auf zwei Klassen aufgeteilt ist. Die Nacktsamer besitzen meist mehrere Keimblätter und werden deshalb als polykotyl bezeichnet. Je nachdem, ob die Keimblätter bei der Keimung die Erdoberfläche durchbrechen, spricht man von epigäischer (über der Erdoberfläche, unsere meisten Kulturpflanzen) oder hypogäischer (unterhalb der Erdoberfläche, z. B. bei der Erdnuss) Keimung.
Primärblätter
Bei vielen Pflanzen folgen auf die Keimblätter Laubblätter, die ebenfalls noch einfacher gestaltet sind als die später gebildeten. Dies sind die sogenannten Primärblätter.
Laubblätter
Dies sind die Blätter, die den Großteil der Blattmasse bei den meisten Pflanzen ausmachen und deren Hauptaufgabe die Photosynthese und Transpiration ist. Besonders für sie gilt der oben in den Abschnitten Anatomie und Morphologische Gliederung beschriebene Aufbau.
Blütenblätter
Morphologisch betrachtet, ist eine Blüte ein Kurzspross, die an diesem Kurzspross sitzenden Blätter sind zu den Blütenblättern umgebildet: die Blütenhüllblätter sind entweder unterschiedlich ausgebildet als Kelchblätter (Sepalen) und Kronblätter (Petalen) oder einheitlich als Perigonblätter (Tepalen); nach innen zu folgen die Staubblätter und Fruchtblätter.
Niederblätter
Niederblätter (Cataphylle) sind in der Regel klein und einfach gestaltet, vielfach schuppenförmig. Vielfach ist nur das Unterblatt ausgebildet. Meist sind sie nicht grün. An der Sprossachse stehen sie unterhalb der Laubblätter, daher der Name. Sie stehen entweder am Beginn des Grund- oder des Seitentriebes, bei Holzgewächsen stehen Niederblätter häufig als Knospenschuppen am unteren Ende des Jahrestriebes (nicht bei allen Gehölzen sind die Knospenschuppen jedoch Niederblätter). Hier wechseln sich Laubblatt- und Niederblattregion periodisch miteinander ab. Niederblätter finden sich auch an Rhizomen, unterirdischen Ausläufern. Auch die Zwiebelschuppen der Zwiebeln sind meist Niederblätter.
Hochblätter
Als Hochblätter (Brakteen) bezeichnet man bei Pflanzen Tragblätter, die in ihrer Blattachsel eine Einzelblüte, einen Blütenstand oder einen Teilblütenstand tragen. Ein Tragblatt einer einzelnen Blüte nennt man Deckblatt. Als Hüllblätter (Involukralblätter) bezeichnet man Hochblätter, die meist zu mehreren einen Blütenstand umgeben. Ihre Gesamtheit nennt man Hülle (Involukrum). Die am Blütenzweig direkt auf die Braktee folgenden Blätter nennt man Vorblätter (Brakteolen).
Häufig unterscheiden sich die Hochblätter von den normalen Laubblättern, z. B. durch eine auffällige Färbung. Von den Niederblättern sind sie nur durch die Stellung im Spross unterschieden. Häufig finden sich zwischen den Laub- und den Hochblättern Übergangsformen (Übergangsblätter).
Blattstellung
- Hauptartikel: Phyllotaxis

Blätter sind an der Sprossachse in gesetzmäßiger, artspezifischer Weise angeordnet. An jedem Knoten der Sprossachse können ein oder mehrere Blätter sitzen, es gibt vier Grundarten der Blattstellung:
- Bei der zweizeiligen oder distichen Blattstellung steht an jedem Knoten nur ein Blatt, Blätter aufeinander folgender Knoten sind um 180° verschoben, sodass sich an der Sprossachse zwei Längszeilen von Blättern ergeben. Vertreter sind viele monokotyle Pflanzen und Schmetterlingsblütler.
- Bei wechselständiger Blattstellung sitzt ebenfalls nur ein Blatt an jedem Knoten, der Winkel zwischen zwei Blättern ist aber von 180° verschieden, die Blätter stehen entlang einer Spirallinie. Diese Anordnung ist für dikotyle Pflanzen charakteristisch.
- Bei der gegenständigen Blattstellung stehen an jedem Knoten zwei Blätter. Bei der dekussierten oder kreuzgegenständigen Blattstellung sind aufeinander folgende Blattpaare jeweils um 90 Grad gedreht, stehen also im rechten Winkel übereinander. Es entstehen vier Längszeilen. Vertreter sind Lippenblütler, Nelkengewächse und Ölbaumgewächse.
- Bei quirliger Blattstellung stehen an jedem Knoten drei oder mehr Blätter, wobei die Blätter des nächstjüngeren Knotens auf Lücke stehen. Vertreter sind z. B. die Rötegewächse (Waldmeister).
Abwandlungen der Blätter
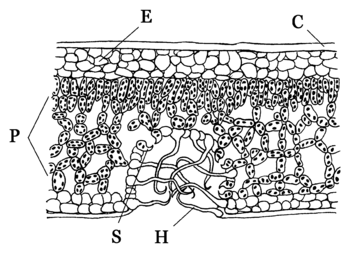 Blattanatomie von Xerophyten. Besonderheiten:
Blattanatomie von Xerophyten. Besonderheiten:
C = verdickte Cuticula,
E = mehrschichtige Epidermis,
H = tote, epidermale Blatthaare,
P = mehrschichtiges Palisaden- und Schwammgewebe,
S = eingesenkte SpaltöffnungenWie bei der Wurzel und der Sprossachse sind auch die Blätter vielfach durch Metamorphosen abgewandelt, um entweder ihre ursprüngliche Funktion an bestimmte Umweltbedingungen angepasst zu erfüllen oder überhaupt andere Funktionen zu übernehmen.
Sonnen- und Schattenblätter
Sonnenblätter, d. h. Blätter, die dem vollen Sonnenlicht ausgesetzt sind, bilden häufig ein mehrschichtiges, kleinzelliges Palisadenparenchym aus. Die Interzellularen im Schwammparenchym sind schwach ausgebildet.
Schattenblätter haben oft ein reduziertes Palisadenparenchym, die Blätter bestehen aus wenigen Zellschichten, die Zellen sind groß und besitzen wenige Chloroplasten. Das Interzellularensystem ist weiträumig, die Palisadenzellen sind kegelförmig. Die Wasserleitungsbahnen sind oft reduziert.
Besonders bei Bäumen (z. B. Rotbuche) treten Sonnen- und Schattenblätter an einer Pflanze auf. Sonnenblätter leiten aber auch zu den xeromorphen Blättern über, Schattenblätter zu den hygromorphen Blättern.
Xeromorphe Blätter
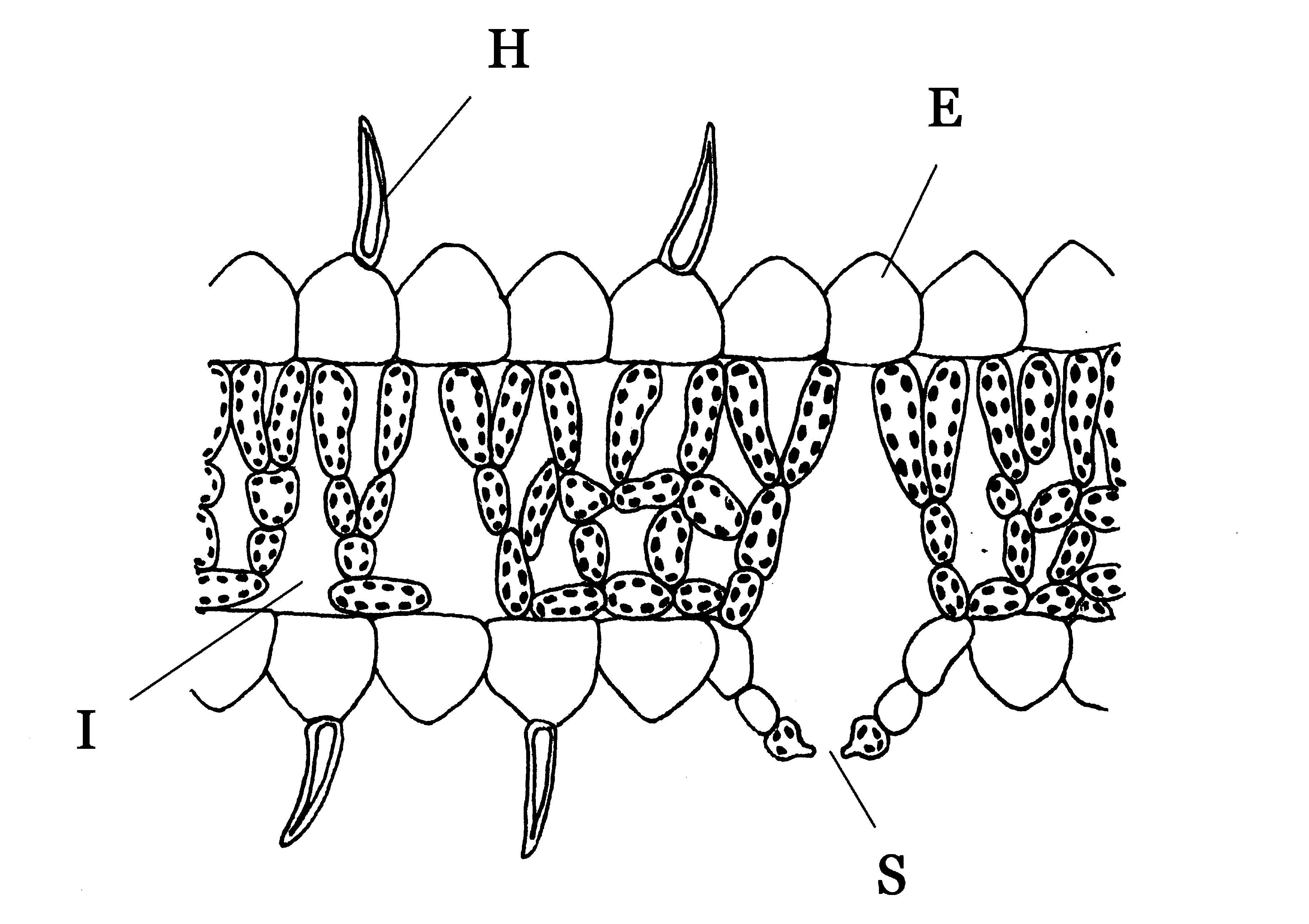
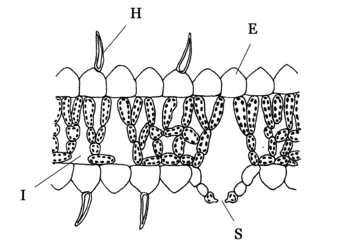 Blattanatomie von Hygrophyten. Besonderheiten:
Blattanatomie von Hygrophyten. Besonderheiten:
E = gewölbte, papillenartige Epidermiszellen,
H = lebende, epidermale Blatthaare,
I = große Interzellulare,
S = herausgehobene SpaltöffnungenViele Pflanzen trockener Standorte reduzieren ihre Blätter vollständig oder wandeln sie in Dornen um, wie z. B. die Kakteengewächse. Dadurch wird die Oberfläche der Pflanze wesentlich reduziert und damit auch die Transpiration.
Zahlreiche Xerophyten behalten jedoch ihre Blätter, deren Aufbau aber stark in Richtung Transpirations-Verminderung abgewandelt ist. Xeromorphe Blätter sind meist derb-lederig (Hartlaubgehölze, wie etwa Lorbeer, Myrte und Ölbaum). Die Spaltöffnungen sind tief in die Blattoberfläche eingesenkt, die dadurch entstehenden Vertiefungen (Krypten) sind mit Haaren versehen, die die Luftkonvektion weiter behindern. Der substomatäre Interzellularraum kann mit Wachs verschlossen sein. Vielfach werden bei Trockenheit die Blätter eingerollt und so die Spaltöffnungen weiter eingeschlossen (z. B. Stipa capillata).
Die Epidermis besitzt eine verdickte Cuticula mit starker Wachseinlagerung. Vielfach sind die Blätter dicht mit toten Haaren besetzt. Dies führt zu einem geringeren Luftaustausch und zu einem deutlich feuchteren Mikroklima direkt an der Blattoberfläche.
Xeromorphe Blätter sind oft äquifazial aufgebaut. Auch das Nadelblatt weist einen typisch xeromorphen Bau auf, da die Nadelgehölze im Winter oft starker Frosttrocknis ausgesetzt sind.
Da eine Verringerung der Transpiration jedoch zu einer Überhitzung führen kann, stellen manche Pflanzen ihre Blätter senkrecht zur Sonneneinstrahlung, wie etwa manche australischen Eukalypten, die „schattenlose Wälder“ bilden.
Hygromorphe Blätter
Hygromorphe Blätter sind eine Anpassung an immerfeuchte Standorte. Zusätzlich zu den Merkmalen der Schattenblätter besitzen sie große, dünnwandige Epidermiszellen, die häufig Chloroplasten führen und nur eine dünne Cuticula besitzen. Die Spaltöffnungen sind oft über die Epidermis emporgehoben, um die Transpiration zu erleichtern. Hygrophyten (Hygromorphe Blätter), die meist in tropischen Gebieten leben, haben nämlich die Schwierigkeit, wegen der hohen Luftfeuchtigkeit Wasser abzugeben, um somit neues (und damit auch Mineralien) aufzunehmen. Im Gegensatz zu Xerophyten die ihre Stoma nach innen gestülpt haben um möglichst wenig Wasser zu transpirieren, haben Hygrophyten ihre Spaltöffnung nach Außen vorgestülpt. Manchmal kommt auch aktive Wasserausscheidung (Guttation) über die Stomata vor, dann hängen Wassertropfen an der hervorgestülpten Stoma, die vom Wind weggeweht oder von Tieren durch Berührung zu Boden fallen. Guttation ist die Ausscheidung von Wasser, das nicht mehr in Gasform vorliegt wie bei der Transpiration.
Nadelblatt
 Ein Zweig mit Nadelblättern.
Ein Zweig mit Nadelblättern.
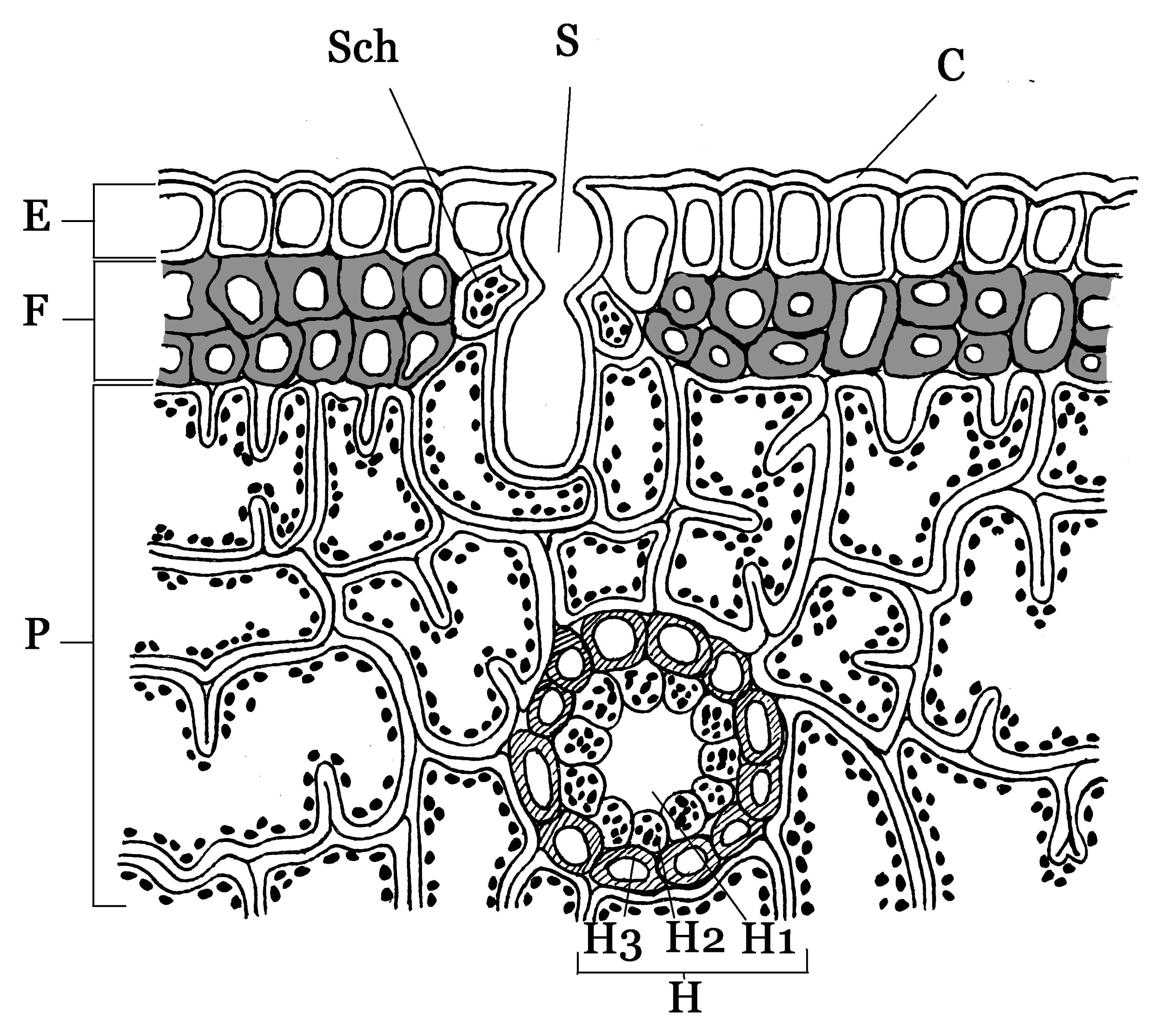
Picea glauca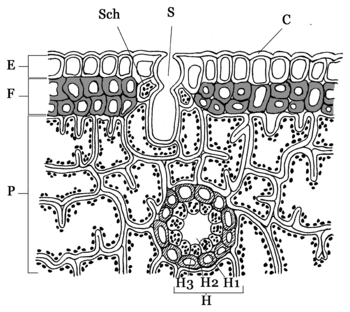 Blattanatomie eines Nadelblattes. Bezeichnung:
Blattanatomie eines Nadelblattes. Bezeichnung:
C = dicke Cuticula,
E = Epidermis,
F = totes Festigungsgewebe (Hypoderm),
H = Harzkanal,
H1 = Lumen (Hohlraum),
H2 = Drüsenepithel,
H3 = sklerenchymatische Scheide,
P = Armpalisaden-Parenchym,
S = eingesenkte Spaltöffnungen,
Sch = SchließzellenDie Nadelblätter der meisten Nadelholzgewächse (Pinophyta) sind großteils eine Anpassung an Trockenheit (Xeromorphie). Die meist immergrünen Bäume sind im Winter der Frosttrocknis ausgesetzt, d. h. durch den gefrorenen Boden kann die Pflanze kein Wasser aufnehmen und muss daher dem Wasserverlust über die Blätter entgegenwirken: Die Nadeln haben eine kleine Oberfläche, eine dicke Cuticula und die Spaltöffnungen sind in die Epidermis eingesenkt.
Nadelblätter weisen weitere charakteristische Merkmale auf – die meisten Blätter sind äquifazial aufgebaut, Schwamm- und Palisadenparenchym sind nicht deutlich getrennt. Darüber hinaus gibt es einige bifaziale Nadelblätter, z. B. das der Weißtanne (Abies alba), die eine unterschiedliche Differenzierung nach Oberseite (Palisadenparenchym) und Unterseite (Schwammparenchym sowie weiße Wachsschicht mit Stomata) ähnlich einem Laubblatt aufweisen. Die Oberfläche der Mesophyllzellen einiger Pinusarten ist durch leistenförmige Wandeinstülpungen vergrößert (Armpalisaden-Parenchym). Zwischen diesem Parenchym und der Epidermis liegt ein sklerotisches (totes) Festigungsgewebe, die so genannte Hypodermis, aus extrem dicken Zellwänden. Die Epidermiszellen sind meistens mit sekundären und tertiären Wandverdickungen ebenfalls fast komplett ausgefüllt und weisen lediglich schmale Verbindungskanäle zur Nachbarzelle auf. Im Mesophyll verlaufen in Längsrichtung meist Harzkanäle. Die ein bis zwei unverzweigten Leitbündel sind von einer gemeinsamen Leitbündelscheide, der Endodermis, umgeben. Der Stofftransport zwischen Leitbündel und Mesophyll erfolgt durch ein spezielles Transfusionsgewebe (Strasburger-Zellen) sowie durch kurze tote Tracheiden. Das Leitbündel besteht wie in der Sprossachse und der Wurzel aus Xylem und Phloem. Dazwischen befindet sich eine dünne Kambiumschicht, die zur Neubildung von Siebzellen bei mehrjährigen Nadelblättern dient (Siebzellen sind sehr kurzlebig, siehe auch Bast). Xylem wird kaum neu gebildet.
Weitere Metamorphosen
Dornen
Dornen dienen den Pflanzen zur Abwehr von Tieren. Blattdornen sind ein- oder mehrspitzige Umbildungen von Blättern oder Blattteilen aus sklerenchymatischem Gewebe. Die Dornen der Berberitze sind Umwandlungen des gesamten Blattes, sie treten an den Langtrieben auf. Nebenblattdornen (Stipulardornen) treten immer paarig auf und sind z. B. bei der Robinie zu finden.
Ranken
Ranken dienen der Pflanze zum Halt an Stützen. Sie können von allen Grundorganen des Blattes abgeleitet sein. Bei der Erbse sind beispielsweise die Endfiedern der Fiederblätter umgebildet, während bei der Platterbse die Ranke durch die Blattspreite gebildet wird, während die Nebenblätter die Photosynthese übernehmen. Bei manchen Pflanzen wird der Blattstiel für das Ranken benutzt – diese winden sich um die Stütze (z. B. Kannenpflanzen oder Zaunwinde).
Speicherorgane
An wasserarmen Standorten sind Blätter häufig zu wasserspeichernden Organen umgewandelt. Solche Blätter sind häufig äquifazial gebaut. An der Wasserspeicherung können entweder die Epidermis und subepidermales Gewebe beteiligt sein, oder es findet im Mesophyll statt.
Wasserspeichernde Zellen besitzen immer sehr große Saftvakuolen. Sukkulente Blätter haben ein dickfleischiges, saftiges Aussehen. Pflanzen mit derartigen Blättern bezeichnet man als Blattsukkulenten. Wie auch bei der Sprosssukkulenz geht die Blattsukkulenz häufig mit dem CAM-Mechanismus einher. Typische Blattsukkulenten sind die Agaven oder die Hauswurz-Arten.
Auch Zwiebeln bestehen aus Blättern und dienen der Speicherung. Eine Zwiebel ist eine äußerst gestauchte, unterirdische Sprossachse, der schalenförmig übereinander liegende, dickfleischige Schuppenblätter aufsitzen. Diese Schuppenblätter sind Niederblätter oder gehen aus dem Blattgrund abgestorbener Laubblätter hervor und dienen der Speicherung von Reservestoffen. In den ungünstigen Jahreszeiten überdauert die Pflanze als Zwiebel. Zwischen den Schuppenblättern treiben Achselknospen bei Beginn einer neuen Vegetationsperiode zu neuen Vegetationskörpern aus und verbrauchen dabei die gespeicherten Reservestoffe. Neben der Küchenzwiebel sind Tulpen, Lilien und Narzissen weitere Beispiele für Zwiebelpflanzen. Zwiebeln kommen nur bei Monokotylen vor.
Phyllodien
Wenn der Blattstiel verbreitert ist und die Funktion der Blattspreite übernimmt, so spricht man von Phyllodien. In diesem Fall ist die Blattspreite meistens stark reduziert. Beispiele finden sich bei den Akazien, bei denen sich häufig mehrere Übergangsstadien von den typischen Fiederblättern bis hin zu spreitenlosen Phyllodien an einer Pflanze finden.

Zweig der Berberitze von unten. In den Achseln der Dornen sitzen die beblätterten Kurztriebe.

Sukkulente Blätter der Dach-Hauswurz

Die Zwiebel besteht aus Blättern

Phyllodien von Acacia confusa
Blätter fleischfressender Pflanzen
 Eine Kanne der fleischfressenden Nepenthes sibuyanensis
Eine Kanne der fleischfressenden Nepenthes sibuyanensisBei vielen fleischfressenden Pflanzen sind die Blätter zu Organen umgewandelt worden, mit denen Beute gefangen und absorbiert wird, je nach Gattung werden sie entweder als Klebe-, Klapp- oder Fallgrubenfallen bezeichnet. Dabei sind bei einigen Pflanzengattungen die Blätter auch zu, teils sehr schnellen, Bewegungen fähig (Sonnentaugewächse, Wasserschläuche). Alle fleischfressenden Pflanzen sind in der Lage, mit der Oberfläche ihrer Fallen die gelösten Nährstoffe der Beute zu absorbieren, die im strengen Sinne karnivoren Pflanzen sind zusätzlich noch mit Drüsen auf der Oberfläche der Fallen versehen, durch die sie Enzyme ausscheiden, die die Beute auflösen.
Blätter der Epiphyten
Epiphyten wachsen auf Bäumen oder anderen Pflanzen und sind daher für ihre Wasser- und Nährstoffversorgung rein auf Niederschläge und Luftfeuchtigkeit (Nebel) angewiesen. Viele Epiphyten bilden mit ihren Blättern trichterförmige Rosetten, in denen sich Regenwasser ansammelt. In den Trichtern der Nestfarne, z. B. (Asplenium nidus), sammelt sich mit der Zeit sogar Humus an, ebenso in den Mantelblättern der Geweihfarne (Platycerium). Die Gattung Dischidia (Asclepiadaceae) bildet schlauchförmige Blätter, in denen sich Ameisenkolonien ansiedeln, die Erde einschleppen. In diese „Blumentöpfe“ wachsen Adventivwurzeln ein. Ähnliches gilt auch für viele Lithophyten.
Die meisten (vor allem epiphytische) Bromeliengewächse, zum Beispiel Tillandsia-Arten, bilden spezielle Absorptionshaare (Saugschuppen) aus, mit deren Hilfe sie Wasser über das Blatt aufnehmen können.
Stoffaustausch über die Oberfläche
Die wichtigste Aufgabe der Blätter ist die Photosynthese, mit der der Austausch von Sauerstoff und Kohlendioxid mit der Umgebungsluft einhergeht, und die Transpiration, also die Abgabe von Wasser an die Atmosphäre. Diese Vorgänge werden in den jeweiligen Artikeln genauer beschrieben. Daneben gibt es noch eine Reihe weiterer Stoffe, die die Blätter über die Luft aufnehmen bzw. an die Luft abgeben können. [3]
Austausch über die Spaltöffnungen
Über die Spaltöffnungen werden vor allem gasförmige und sehr flüchtige Substanzen aufgenommen. Die wichtigsten sind Schwefeldioxid, Ammoniak und Stickstoffdioxid. Ammoniak kann in Gebieten mit intensiver Tierhaltung 10 bis 20 % des Pflanzenstickstoffs liefern. Die Aufnahme von Ammoniak durch die Spaltöffnungen steigt linear mit der Außenkonzentration. Dasselbe gilt für Stickstoffdioxid. Schwefeldioxid führt in hohen Konzentrationen zur Schädigung der Photosynthese, geringe Konzentrationen können besonders bei Schwefelmangel im Boden zu besserem Wachstum führen.
Pflanzen können aber über ihre Blätter auch Nährstoffe verlieren. So wurde der Verlust an Stickstoff durch die stomatäre Abgabe von Ammoniak für Reis auf 15 kg Stickstoff pro Hektar, für Weizen auf sieben kg Stickstoff pro Hektar berechnet, was in letzterem Fall 20 % der Düngergabe entsprach. Bei hoher Schwefeldioxid-Belastung geben Blätter Schwefelwasserstoff ab. Dies wird als Entgiftungsmechanismus gedeutet. Aber auch Pflanzen ohne Schwefeldioxid-Belastung geben flüchtige Schwefelverbindungen ab, für Hafer und Raps wurden Werte von zwei bis drei Kilogramm Schwefel pro Hektar und Jahr errechnet. Pflanzen mit hohem Selen-Gehalt geben ebenfalls flüchtige Selen-Verbindungen, wie etwa Dimethylselen ab.
Aufnahme von gelösten Stoffen, Blattdüngung
Die Aufnahme von gelösten Stoffen über die Blätter ist bei Landpflanzen durch die Cuticula der Epidermis stark eingeschränkt. Niedermolekulare Verbindungen wie Zucker sowie Mineralstoffe und Wasser können durch hydrophile Poren die Cuticula passieren. Diese Poren haben einen Durchmesser von einem Nanometer, dadurch kann z. B. Harnstoff (Durchmesser 0,44 Nanometer) leicht passieren. Die Poren sind negativ geladen, so dass Kationen leichter passieren können als Anionen. Damit wird z. B. Ammonium rascher aufgenommen als Nitrat. Poren treten besonders häufig in der Zellwand der Schließzellen auf, womit die häufig beobachtete positive Korrelation zwischen der Anzahl der Stomata und der Nährstoffaufnahme aus flüssigem appliziertem Dünger erklärt werden kann.
Die weitere Aufnahme in die Zelle verläuft gleich wie bei der Nährstoffaufnahme der Wurzeln über den Apoplasten. Die Aufnahmerate ist bei gleicher externer Nährstoffkonzentration jedoch bei Blättern aufgrund des zusätzlichen Engpasses der Cuticula wesentlich geringer als bei der Wurzel. Im Gegensatz zu Wurzeln wird die Ionenaufnahme von Blättern durch Licht gefördert. Die Aufnahmerate ist auch abhängig von der internen Nährstoffkonzentration, d. h. die Aufnahme ist bei Nährstoffmangel rascher.
In natürlichen Ökosystemen ist die Aufnahme von Nährstoffen nur bei Stickstoff und Schwefel von Bedeutung.
Blattdüngung führt den Pflanzen die Nährstoffe in der Regel rascher zu als herkömmliche Bodendüngung. Daher wird sie trotz mancher Nachteile in vielen Bereichen eingesetzt.
Zu den Nachteilen zählen:
- Abperlen von der hydrophoben Blattoberfläche
- Abwaschen durch Regen
- Bestimmte Nährstoffe wie Kalzium können von den Blättern nicht mehr in andere Pflanzenteile transportiert werden.
- Mit einer Blattdüngung kann nur eine begrenzte Menge an Nährstoffen aufgebracht werden (Ausnahme ist Harnstoff).
- Es kann zu Schäden am Blatt führen: Nekrosen und Verbrennungen.
Unter bestimmten Bedingungen ist die Blattdüngung dennoch von großer praktischer Bedeutung:
- Nährstoffmangel im Boden: Auf Kalkböden, die Eisen immobilisieren, kann Blattdüngung mit Eisen vor Chlorosen schützen. Dasselbe gilt für Mangan-Mangel. Bei Obstbäumen kann eine im Herbst applizierte Blattdüngung mit Bor vor Bormangel schützen.
- Trockene Oberböden: In semiariden Gebieten ist die Nährstoffverfügbarkeit durch die Austrocknung des Oberbodens oft drastisch reduziert. In solchen Fällen ist Blattdüngung effektiver als Bodendüngung.
- Während der Samenfüllung ist bei vielen Pflanzen die Wurzelaktivität reduziert. Auch hier kann Blattdüngung zu höheren Nährstoffgehalten und auch Ernteerträgen führen.
Bei der Bewässerung mit salzhaltigem Wasser kann es zu stark erhöhter Aufnahme von Chlorid und Natrium kommen. Dieser Effekt ist bei dieser Bewässerungsart stärker als es bei der Tröpfchenbewässerung der Fall ist.
Leaching
Der Verlust von organischen und anorganischen Stoffen durch Flüssigkeiten, besonders Regen und Bewässerung, wird meist mit dem englischen Begriff Leaching (Lecken, Auswaschen) bezeichnet. Man unterscheidet vier Arten:
- Aktive Exkretion von Lösungen, z. B. die Exkretion von Salz durch Salzdrüsen in Halophyten.
- Exkretion von inorganischen Lösungen an Blattspitzen und -rändern durch Wurzeldruck: Guttation.
- Leaching aus verletzten Blattbereichen.
- Leaching aus dem Apoplasten von intakten Blättern.
Von wesentlicher ökologischer Bedeutung sind die letzten beiden Arten. Der Verlust ist höher in alten Blättern und unter Stress (Trockenheit, hohe Temperatur, Ozon). Auch ein niedriger pH-Wert des Regens (saurer Regen) erhöht das Leaching. Die Kationen des Blattes werden wie in einem Ionenaustauscher durch Protonen ersetzt.
Mit Ausnahme von Stickstoff und Schwefel überwiegt in natürlichen Ökosystemen das Leaching. Besonders hoch ist der Verlust in Gebieten mit starken Regenfällen. Für tropische Regenwälder wurden folgende Jahreswerte berechnet (in Kilogramm pro Hektar): Kalium 100–200, Stickstoff 12–60, Magnesium 18–45, Kalzium 25–29, und Phosphor 4–10. In gemäßigten Breiten fällt – verglichen mit den internen Blattgehalten – die hohe Leaching-Rate von Kalzium und Mangan auf. Diese Elemente sind nicht phloemmobil, d. h. sie sammeln sich in den Blättern an. Das starke Leaching wird als Strategie der Pflanzen gedeutet, zu hohe Konzentrationen zu vermeiden.
Neben Mineralstoffen können auch größere Mengen an organischen Verbindungen durch Leaching verloren gehen. Für Wälder der gemäßigten Breiten wurden Werte von 25 bis 60 Kilogramm Kohlenstoff pro Hektar und Jahr errechnet, für tropische Wälder schätzt man die Menge auf mehrere hundert Kilogramm.
Das Blatt als Lebensraum
 Minen in einem Rosskastanien-Blatt
Minen in einem Rosskastanien-BlattBlätter enthalten als physiologisch sehr aktive Pflanzenteile (Photosynthese) in der Regel sehr viele Nährstoffe und sind daher eine sehr wichtige Nahrungsquelle für eine Unzahl von Tierarten. Etliche Tiergruppen benutzen jedoch die Blätter zugleich auch als Lebensraum. Hierzu zählen etwa die Blattminierer wie z. B. die Rosskastanienminiermotte. Dies sind Insekten, deren Larven Gänge im Inneren der Blätter fressen. Weitere Beispiele sind Blattroller (Familie Attelabidae), deren Weibchen Blätter einrollen und darin die Eier ablegen, so dass die Larven geschützt sind und Gallwespen, die mit der Eiablage die Bildung sogenannter Gallen, Wucherungen des Pflanzengewebes, auslösen, von denen sich die Larven ernähren.
Blätter werden auch von einer Vielzahl von Pilzen befallen, wie etwa von Mehltau-, Brand- und Rostpilzen, die in landwirtschaftlichen und gartenbaulichen Kulturen große Schäden anrichten können. In Blättern leben auch oft endophytische Pilze, die zu keiner erkennbaren Schädigung der Pflanze führen.
Auf Blättern können wiederum andere Pflanzen leben, man nennt diese Lebensform Epiphyllie. Epiphylle Moose und Flechten sind besonders häufig in den tropischen Regen- und Nebelwäldern.
Quellenangaben
- ↑ vgl. Lexikon der Biologie. Bd 3. Spektrum Akademischer Verlag, Heidelberg 2000, S. 1. ISBN 3-8274-0328-6; Sitte u. a., 2002, S. 717–750.
- ↑ Wilhelm Seyffert (Hrsg.): Lehrbuch der Genetik. Spektrum Akademischer Verlag, Heidelberg ²2003, S. 712f. ISBN 3-8274-1022-3
- ↑ Der Abschnitt folgt: Horst Marschner: Mineral Nutrition of Higher Plants. Academic Press, London ²1995, S. 116–30. ISBN 0-12-473543-6
Literatur
- W. Braune, A. Leman, H. Taubert: Pflanzenanatomisches Praktikum. Bd I. Zur Einführung in die Anatomie der Vegetationsorgane der Samenpflanzen. Gustav Fischer, Jena 61991, bes. S. 176–220, ISBN 3-334-60352-0
- Manfred A. Fischer (Hrsg.): Exkursionsflora für Österreich, Liechtenstein und Südtirol. Oberösterreichisches Landesmuseum, Linz 2005, bes. S. 72–84, ISBN 3-85474-140-5
- Stefan Klotz, Dieter Uhl, Christopher Traiser, Volker Mosbrugger: Physiognomische Anpassungen von Laubblättern an Umweltbedingungen. in: Naturwissenschaftliche Rundschau. Stuttgart 58.2005,11, S. 581–586, ISSN 0028-1050
- U. Lüttge, G. Kluge, G. Bauer: Botanik – Ein grundlegendes Lehrbuch. VCH Verlagsgesellschaft, Weinheim 1989 (Nachdr.), ISBN 3-527-26119-2
- Klaus Napp-Zinn: Anatomie des Blattes. T II. Blattanatomie der Angiospermen. B: Experimentelle und ökologische Anatomie des Angiospermenblattes. in: Handbuch der Pflanzenanatomie. Bd 8 Teil 2 B. Borntraeger, Stuttgart 1988 (2. Lieferung), ISBN 3-443-14015-7
- Schmeil, Fitschen: Flora von Deutschland und angrenzender Länder. Quelle & Meyer, Heidelberg/Wiesbaden 891993, ISBN 3-494-01210-5
- P. Sitte, E. W. Weiler, J. W. Kadereit, A. Bresinsky, C. Körner: Lehrbuch der Botanik für Hochschulen. Begründet v. E. Strasburger. Spektrum Akademischer Verlag, Heidelberg 352002, ISBN 3-8274-1010-X
Weblinks
- Aufbau eines typischen Laubblattes
- Blattquerschnitt (Übersicht)
- Blattformen und Blattstellungen
- Blatt-Bilder aus dem Bildarchiv der Universität Basel
- Blattnervatur-Bilder aus dem Bildarchiv der Universität Basel
- spektrumdirekt: Der Ursprung der Blätter
- Beispiele für fossile, pliozäne Blätter
- Die meisten Blätter sind bifazial gebaut, d. h. es wird eine Ober- und Unterseite ausgebildet.




















{kind=link}
Wikimedia Foundation.