- Osmotischer Druck
-
Als Osmose (von griechisch ὠσμός, ōsmós = „Eindringen, Stoß, Schub, Antrieb“) wird in den Naturwissenschaften der gerichtete Fluss von Molekülen durch eine semipermeable (auch: selektiv permeable) Membran bezeichnet. Osmose ist für viele Abläufe in der Natur von Bedeutung, besonders für die Regulation des Wasserhaushalts von Zellen und Pflanzen, und findet als Trennverfahren Anwendung in der Medizin und Verfahrenstechnik. Die theoretische Erklärung der Osmose liefert die statistische Mechanik. Die physikalischen Abläufe bei der Membrandurchquerung sind auf mikroskopisch-molekularer Ebene auch zu Beginn des 21. Jahrhunderts noch immer Gegenstand aktiver Forschungstätigkeit.[1][2][3]
Inhaltsverzeichnis
Einleitung
Ein anschauliches Beispiel für die Wirkung der Osmose ist das Aufplatzen reifer Kirschen nach Benetzung mit Regenwasser. Das Wasser auf der Außenseite der Frucht enthält nur sehr wenig gelöste Teilchen, hat also ein hohes chemisches Potenzial. Es dringt durch die äußere Haut in die Frucht ein, in der das Wasser als Lösungsmittel durch den hohen Zuckergehalt und andere gelöste Stoffe ein niedriges chemisches Potenzial aufweist. Durch den Wassereinstrom steigt der Druck im Inneren der Frucht und führt zum Aufreißen ihrer äußeren Haut. Diese ist neben anderen Stoffen für Wasser durchlässig, nicht jedoch für Zuckermoleküle; aufgrund dieser Eigenschaften wirkt sie als semipermeable Membran. Wassermoleküle können diese Membran prinzipiell in beide Richtungen passieren, werden jedoch im Inneren der Frucht stärker „festgehalten“. Die Wassermoleküle müssen dort mit den anderen gelösten Molekülen und Teilchen um den Zugang zur Membran konkurrieren, so dass weniger Wassermoleküle pro Zeiteinheit nach außen dringen als umgekehrt.
Triebkraft der spontan ablaufenden Osmose ist der Unterschied zwischen den chemischen Potenzialen eines oder mehrerer Stoffe in den durch eine Membran getrennten Phasen. Diese können aus flüssigen und gasförmigen Lösungen oder Reinstoffen bestehen. Teilchen (Atome, Moleküle oder Ionen) der Komponenten, für die die Membran durchlässig ist, diffundieren in Richtung ihres niedrigeren chemischen Potenzials. Der aus dieser Bewegung resultierende Mischungsvorgang verringert die Gibbs-Energie (oder freie Enthalpie) des Gesamtsystems; daher läuft der Vorgang freiwillig ab (siehe auch exergone und endergone Reaktionen). Im Allgemeinen ist es ein Lösungsmittel, das aufgrund der unterschiedlichen Anzahl gelöster Teilchen in Richtung seines niedrigeren chemischen Potenzials diffundiert.
In geschlossenen Systemen erfolgt durch Osmose ein Ausgleich der Potenzialdifferenzen, die osmotische Bewegung hält solange an, bis das chemische Potential der diffundierenden Komponenten auf beiden Seiten der Membran ausgeglichen ist; zwischen beiden Phasen hat sich dann ein Gleichgewicht eingestellt. Erfolgt der Stofffluss in ein geschlossenes Volumen, muss sich zwangsläufig der Druck in diesem Volumen (der Seite mit dem anfangs niedrigerem Potenzial) erhöhen; diese Differenz wird als osmotischer Druck bezeichnet. Der osmotische Druck ist eine kolligative Eigenschaft, da er von der Anzahl der gelösten Teilchen abhängt.
Geschichte und Definitionen des Begriffs Osmose
Geschichtliches
Im Jahre 1748 wurde von Jean-Antoine Nollet ein Experiment beschrieben, bei dem ein mit „Weingeist“ gefüllter und einer entfetteten Schweinsblase verschlossener Zylinder in reines Wasser getaucht wurde. Die Blase blähte sich auf und zerriss manchmal nach einer Weile.[4] Den Begriff „Osmose“ prägte der Botaniker Henri Dutrochet. Er bezeichnete den Einstrom von Wasser in eine mit einer Schweinsblase umschlossene Messkammer als Endosmose, die Auswärtsbewegung als Exosmose[5].
1877 beschrieb der deutsche Botaniker Wilhelm Pfeffer erstmals eine Methode zur quantitativ exakten Messung des osmotischen Drucks wässriger Lösungen. Er entwickelte dazu ein Osmometer, die Pfeffersche Zelle, aus Tonzellen, die er mit den von Moritz Traube zuvor beschriebenen kolloidalen Niederschlagsmembranen, wie Kupfer(II)-hexacyanoferrat(II), beschichtete. Der niederländische Chemiker Jacobus Henricus van ’t Hoff veröffentlichte zehn Jahre später seine wegweisende Arbeit zur Analogie von Gasdruck und osmotischem Druck von Lösungen,[6], für die er 1901, neben anderen Leistungen, für seine Arbeiten zur Natur des osmotischen Drucks mit dem ersten Nobelpreis für Chemie ausgezeichnet wurde.[7] Das Pfeffer'sche Verfahren zur Messung des osmotischen Drucks wurde um die Jahrhundertwende durch Harmon Northrop Morse wesentlich verbessert, der das Membranmaterial mittels Elektrolyse auf die Wände der Tonzellen aufbrachte. 1905 veröffentlichte Albert Einstein seine grundlegende Arbeit zur Erklärung der Osmose mit Mitteln der „molekularkinetischen Theorie“.[8]
Definitionen
Während ursprünglich „unter Osmose die partielle Diffusion einzelner Komponenten von Flüssigkeitsgemischen durch poröse Wände“[9] verstanden wurde – was gelöste Stoffe mit eingeschliesst, Pfeffer spricht in diesem Zusammenhang von der Diosmose gelöster Körper[10]) – wurde später, wohl durch die Verwendung kolloidaler Membranen in Osmometern und den physikalischen Erklärungsansätzen auf mikroskopischer Ebene motiviert, das Konzept der semipermeablen Membran zum zentralen Begriff für die Beschreibung des Phänomens Osmose. In der heutigen Literatur wird Osmose unterschiedlich, teils auch widersprüchlich definiert, wie die nachfolgende Beispiele belegen:
- „Osmose, die Bewegung von Wasser durch eine semipermeable Membran aufgrund eines Unterschieds im osmotischen Druck, ...“[11]
- „Das Phänomen Osmose besteht im Bestreben eines reinen Lösungsmittels durch eine semipermeable Membran in eine Lösung hineinzuwandern.“[12]
- „Unter Osmose versteht man die Diffussion von Teilchen durch eine selektiv permeable Membran“[13]
- Verzichtet wird in einigen Lehrbüchern und wissenschaftlichen Arbeiten auf eine wörtlichde Definition der Osmose (beispielsweise in [4], [14] oder [8]), die Bedeutung muss sich dem Leser anhand der zur Darstellung benutzten Beispiele erschließen.
Tatsächlich können bei geeigneter Membranbeschaffenheit außer Wasser auch andere Flüssigkeiten sowie Stoffe aus einer Gasphase (Gasosmose) an osmotischen Vorgängen beteiligt sein.[15] Bestimmte osmotische Phänomene wie der Gibbs-Donnan-Effekt beruhen darauf, dass neben dem Lösungsmittel auch gelöste Ionen die Membran passieren können.[4] Dialyse, bei der neben dem Lösungsmittel andere gelöste Substanzen durch eine Membran diffundieren, wird als ein auf Osmose beruhendes Phänomen beschrieben.[16][4]
Von Thermoosmose spricht man, wenn der Stofffluss aufgrund eines Temperaturunterschieds erfolgt.[17] Elektroosmose bezeichnet die Bewegung einer Flüssigkeit entlang einer elektrochemischen Doppelschicht wie sie bei der Elektrophorese auftritt.
Anmerkungen zu den Definitionsbeispielen
- Ähnliche Definition wie diese finden sich meist in Schulbüchern, Lehrbüchern der Biologie sowie der Medizin. Zwar ist die Beschränkung des Begriffs Osmose auf Wasser als einzig durchgelassene Substanz aus Sicht der physikalischen Chemie nicht gerechtfertigt, erfüllt aber im Zusammenhang mit lebenden Systemen oder didaktischen Einführungen ihren Zweck, da es ja hier in den allermeisten Fällen um die Bewegung von Wasser durch eine Membran geht.
- Hier wird zwischen Osmose als Bewegung des Lösungsmittels (dies kann flüssig oder gasförmig sein) und dem Verhalten der gelösten Substanzen unterschieden, da eine ideal semipermeable Membran als ausschließlich ist für das Lösungsmittel durchlässig gedacht wird. Reale Membranen, insbesondere Biomembranen, sind in gewissen Maße auch für gelöste Stoffe und Ionen durchlässig. Dieser Umstand kann bei der Berechnung des osmotischen Flusses durch eine Membran durch Einführung eines Reflektionskoeffizienten der Membran für jede in der Lösung vorhandene Substanz berücksichtigt werden[18]. Osmose durch eine ideal semipermeable Membran kann somit als Spezialfall eines weiter gefassten Osmosebegriffs verstanden werden, der auch sogenannte leaky membranes oder leaky pores (lecke Membranen oder Poren) umfasst[19] .
- Häufig wird der Begriff Diffusion zur Erklärung der Osmose verwendet; dies geht auf einen Vorschlag des Physiologen Adolf Fick aus dem Jahr 1855 zurück.[20] Lange Zeit war die physikalische Natur dieser Bewegung umstritten[21], bis 1957 Mauro zeigen konnte, dass ein Lösungsmittel die Membranporen mit derselben Geschwindigkeit passiert, unabhängig davon, ob die Bewegung aufgrund eines osmotischen Potenzialunterschieds oder durch Anlegen eines hydrostatischen Drucks erfolgt. Daraus wurde geschlossen, dass der osmotische Fluss ein Massenfluss („convective“ oder auch „non-diffusional flow“) sei, der nicht den Fick'schen Diffusionsgesetzen unterliegt.[22]
Grundlagen
 Osmose: links ein U-Rohr mit verschieden konzentrierten Lösungen unmittelbar nach dem Einfüllen, rechts dasselbe U-Rohr zu einem späteren Zeitpunkt. Das osmotische Gleichgewicht ist erreicht, wenn der physikalische Druck der rechten Flüssigkeitssäule den durch Konzentrationsunterschiede verursachten osmotischen Druck ausgleicht.
Osmose: links ein U-Rohr mit verschieden konzentrierten Lösungen unmittelbar nach dem Einfüllen, rechts dasselbe U-Rohr zu einem späteren Zeitpunkt. Das osmotische Gleichgewicht ist erreicht, wenn der physikalische Druck der rechten Flüssigkeitssäule den durch Konzentrationsunterschiede verursachten osmotischen Druck ausgleicht.Bei der Osmose kann ein Konzentrationsunterschied zwischen beiden Seiten nur durch den Fluss der Substanzen ausgeglichen werden, die aufgrund Ihrer Eigenschaften die Membran passieren können. In vielen Fällen ist dies das Lösungsmittel, z. B. Wasser in biologischen Systemen.
Wie im Beispiel rechts dargestellt, fließt Wasser von der Seite der geringeren Konzentration (in der Abb. die linke Hälfte des U-Rohrs) durch die Membran; seine Menge nimmt dort ab und führt zu einer entsprechenden Volumenvergrößerung auf der anderen Seite. Die Bewegung des Lösungsmittels endet, sobald sich auf beiden Membranseiten ein gleichgroßes chemisches Potential eingestellt hat. In der Abbildung ist es der hydrostatische Druck der rechten Flüssigkeitssäule, der dem weiteren Fluss von Lösungsmittel entgegenwirkt.
Wird nach Erreichen des Gleichgewichtszustands das Wasser einer Seite mit D2O oder 3H2O (schweres Wasser) markiert, so lässt sich zeigen, dass weiter ein Austausch von Wassermolekülen erfolgt.[23] Im Gleichgewicht ist jedoch die Menge der einströmenden und ausströmenden Moleküle auf beiden Seiten gleich.
Thermodynamische Betrachtung
Die spontan verlaufende Osmose beinhaltet einen Mischungsvorgang – ein Stoff tritt aus einer Phase in die andere über und vermischt sich dort mit den vorhandenen Komponenten. Freiwillig ablaufende Mischungsvorgänge führen in abgeschlossenen Systemen zu einer Verringerung der freien Enthalpie (Gibbsche Energie) und zu einer Vergrößerung der Entropie. Im Fall der Osmose verhindert die selektiv durchlässige Membran als Barriere, dass mit Erreichen des thermodynamischen Gleichgewichts sämtliche Konzentrationsunterschiede verschwinden; es sind zu jedem Zeitpunkt zwei getrennte Phasen vorhanden. (Tatsächlich drei Phasen, wenn die Membran in die Betrachtung mit einbezogen wird.)
Die Gesamtänderung der freien Enthalpie in der osmotischen Zelle muss daher als die Summe der Änderungen in beiden Phasen geschrieben werden:
wobei das Superskript (x) den jeweiligen Abschnitt x der osmotischen Zelle bezeichnet. Wenn nur eine Stoffart L (z.B. das Lösungsmittel) von der Membran durchgelassen wird, kann der Zusammenhang zwischen freier Enthalpie und Änderung der Stoffmenge dnL durch
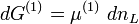 und
und 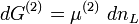
beschrieben werden, wobei μ das chemische Potenzial in Joule bezeichnet und n die Menge (in Mol) des Stoffes, der die Membran passiert. Da der Zufluss auf Seite (2) den Verlust auf Seite (1) ausgleicht, folgt für die gesamte Änderung der freien Enthalpie:
Solange das Gleichgewicht noch nicht erreicht ist (dG > 0) gilt für die chemischen Potenziale:
- μ(1) > μ(2)
und im Gleichgewicht (dG = 0):
- μ(1) = μ(2)
Nimmt mehr als eine Stoffart am osmotischen Prozess teil, müssen obenstehende Relationen um die chemischen Potenziale der zusätzlichen Komponenten erweitert werden (siehe auch Abschnitt Das osmotische Potenzial nichtidealer Lösungen).
Die Bedingung für das osmotische Gleichgewicht ist, dass die chemischen Potenziale der von der Membran durchgelassenen Stoffe (meist das Lösungsmittel) in beiden Phasen gleich sind. Solange noch eine Potenzialdifferenz besteht, werden sich Teilchen der jeweiligen Komponente in Richtung des niedrigeren Potenzials bewegen. Andererseits kann durch Erhöhen des chemischen Potenzials, etwa durch Anwendung von Druck oder eine Temperatursteigerung auf einer Seite der Membran, die Bewegungsrichtung umgekehrt werden. Osmose durch eine Membran, im Gegensatz zur Diffusion in einem einphasigen System, ist ein reversibler Vorgang.[24]
Lösungseffekt als unmittelbare Ursache
Durch den Lösungseffekt verringert sich das chemische Potenzial des Lösungsmittels in der Lösung gegenüber dem reinen Stoff, verbunden mit einer Erniedrigung des Sättigungsdampfdrucks bei Flüssigkeiten oder des Partialdrucks bei Gasen. Damit ist der Lösungseffekt eine unmittelbare Ursache für Osmose.
Dagegen wirkt ein Konzentrationsgefälle der gelösten Stoffe mittelbar, da nur, wenn die Konzentrationsunterschiede zu einer Differenz des chemischen Potenzials des Lösungsmittels in beiden Lösungen führen, Osmose stattfindet. Sind auf beiden Seiten der Membran unterschiedliche Stoffe in gleicher Teilchenzahl gelöst, so ist das chemische Potenzial des Lösungsmittels gleich und es tritt somit keine Osmose auf. Darauf beruht die Wirkung der Osmolyte, mit denen sich Zellen gegen osmotischen Druck schützen können.[25]
Molekularkinetische Betrachtung
Einen Ansatz zur Erklärung der Osmose mit den Mitteln der statistischen Mechanik lieferte Albert Einstein 1905 in seiner Arbeit Über die von der molekularkinetischen Theorie der Wärme geforderte Bewegung von in ruhenden Lösungen suspendierten Teilchen. Darin beschreibt er, dass sich suspendierte von „eingelösten“ Teilchen lediglich durch ihre Größe unterscheiden und beide daher nach der molekularen Theorie der Wärme einen osmotischen Druck hervorrufen. Nach Einstein wirkt auf suspendierte Teilchen eine Kraft, die mit den osmotischen Druckkräften in dynamischem Gleichgewicht steht; die Bewegung kann als Überlagerung zweier Prozesse aufgefasst werden: Einer Bewegung der suspendierten Teilchen aufgrund einer auf jedes einzelne Teilchen einwirkenden Kraft, sowie „eines Diffusionvorganges als Folge der ungeordneten Molekularbewegung der Wärme.“[26]
Osmotischer Druck
Analogie zum Gasdruck
 Abb. 1 Osmotischer Druck im Gleichgewicht
Abb. 1 Osmotischer Druck im Gleichgewicht Abb. 2 Ausgleich des osmotischen Drucks mittels eines Kolben
Abb. 2 Ausgleich des osmotischen Drucks mittels eines KolbenWird ein mit Zuckerlösung (A) gefülltes Gefäß in reines Wasser (B) gebracht, und ist seine Wand nur für die gelösten Zuckermoleküle undurchlässig, so wird sich durch den Einstrom von Wasser der Druck im Innenraum erhöhen. Der so erzeugte osmotische Druck wirkt dem Wassereinstrom entgegen; die Bewegung des Wassers endet mit Erreichen des Gleichgewichts (Abb. 1). Die gleiche Druckverteilung wie in Abb. 1 kann ohne Wassereinstrom erreicht werden, wenn (z.B. über einen Kolben) ein gleichgroßer Druck auf die Flüssigkeit (A) wirkt. (Abb. 2). Durch Erhöhen oder Erniedrigen des Kolbendrucks verändert sich die Konzentration der Lösung (A), da dann Wasser durch die Gefäßwände entsprechend aus- oder einströmt. Dieses Prinzip wird bei der Umkehrosmose (in älteren Schriften auch als Anti-Osmose bezeichnet) eingesetzt; dabei wird unter Druck eine Lösung weiter aufkonzentriert, um die darin gelösten Stoffe zu entfernen.
Diese grundlegende Analogie zwischen osmotischem und Gasdruck wurde zuerst 1887 von dem niederländischen Chemiker van ’t Hoff beschrieben.[24] Als Ursache des osmotischen Drucks sah er die Stöße der gelösten Teilchen auf die (für sie undurchlässige) Membranwand (Solute bombardment theory)[2] an. Der Einfluss der Wassermoleküle sei dagegen auf beiden Membranseiten gleich und würde sich daher gegenseitig aufheben. Gegen diese Interpretation spricht, dass keine Durchbiegung der Membran beobachtet wird, wenn der hydrostatische Druck auf beiden Seiten gleich ist.[27][28]
Anwendung der Gasgesetze
In verdünnten flüssigen Lösungen gelten dieselben Gesetze wie für ideale Gase (Boyle-Mariotte-Gesetz, Gay-Lussac-Gesetz, Avogadro-Gesetz). Der osmotische Druck
- ist proportional zu der molaren Konzentration des gelösten Stoffes
- ist proportional zur absoluten Temperatur
- von Lösungen hängt nur von der Teilchenzahl des gelösten Stoffes (molaren Konzentration) ab (→ kolligative Eigenschaft)
- einer Lösung von 1 Mol in 22,4 l Lösungsmittel beträgt bei 273,15 K (0 °C) 101,325 kPa (Standarddruck)
Diese Aussagen werden durch das van ’t Hoffsche Gesetz zusammengefasst:
Hierbei ist Π der osmotische Druck in Pascal, c = n/V die Stoffmengenkonzentration (molare Konzentration) der Lösung, R die universelle Gaskonstante, T die absolute Temperatur in K. In dieser Form gilt das Gesetz nur für verdünnte Lösungen (< 0,1 M), wie die idealen Gasgesetze nur bei niedrigen Drücken gelten (die Wechselwirkungen der Teilchen miteinander können vernachlässigt werden).
Osmotischer Druck und Dampfdruck
Der Dampfdruck einer Lösung ist immer niedriger als der des reinen Lösungsmittels L (Lösungs- oder Verdünnungseffekt). Das Hinzufügen eines Stoffes zu L erniedrigt dessen chemisches Potential
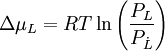 .
.
Dabei ist
 der Dampfdruck des reinen Lösungsmittels und
der Dampfdruck des reinen Lösungsmittels und 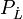 der Lösung. Der osmotische Druck wirkt entgegensetzt und führt zu einer Vergrößerung von μL
der Lösung. Der osmotische Druck wirkt entgegensetzt und führt zu einer Vergrößerung von μLFür Flüssigkeiten kann das partielle Molvolumen
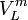 als druckunabhängig angesehen werden. Unter dieser Bedingung kann aus beiden obenstehenden Termen die Gleichung:
als druckunabhängig angesehen werden. Unter dieser Bedingung kann aus beiden obenstehenden Termen die Gleichung:abgeleitet werden. Sie besagt, dass der osmotische Druck dem äußeren Druck entspricht, der den Dampfdruck einer Lösung auf den Dampfdruck des reinen Lösungsmittels L erhöhen würde. Die treibende Kraft der Osmose ist die durch den Lösungseffekt verursachte Dampfdruckerniedrigung des Lösungsmittels. Eine osmotische Zelle befindet sich im Gleichgewicht, wenn der osmotische Druck durch eine gleichgroße Gegenkraft ausgeglichen wird.
Salzlösungen (Elektrolyte)
Der osmotische Druck einer Salzlösung ist immer höher als es dem van ’t Hoffschen Gesetz in der Form Π = c⋅R⋅T entspricht, und zwar häufig doppelt oder dreifach so hoch. Dieser Effekt beruht darauf, dass Salze beim Lösungsvorgang in negativ und positiv geladene Ionen zerfallen (→ Dissoziation) und gelöst eine höhere Teilchenzahl aufweisen, als der Stoffmenge im festen Zustand entspricht. Für vollständig gelöste Salze (starke Elektrolyte) ist dies ein ganzzahliges Vielfaches der ursprunglichen Stoffmenge. Durch Erweiterung der Gleichung um einen Faktor:
kann dieser Effekt berücksichtigt werden. Hierbei ist i der van ’t Hoff-Faktor, eine dimensionslose Zahl. Für Natriumchlorid (NaCl), Kaliumchlorid (KCl) und andere binäre Elektrolyte ist i = 2; für 1,2-wertige Elektrolyte wie Natriumsulfat (Na2SO4) ist i = 3.
Für Salze, die in Lösung nicht vollständig zerfallen (schwache Elektrolyte), kann der van ’t Hoff-Faktor aus dem Dissoziationsgrad α berechnet werden:
wobei ν die Anzahl der Ionen pro Salzmolekül ist.
Das osmotische Potenzial nichtidealer Lösungen
Das van ’t Hoffsche Gesetz gilt nicht für Lösungen, bei denen die Wechselwirkungen der Moleküle nicht mehr vernachlässigt werden kann. Hier muss das chemische Potential aus der Gibbsschen Fundamentalgleichung verwendet werden. Im thermodynamischen Gleichgewicht ist die freie Enthalpie (Gibbs-Energie) einer osmotischen Zelle minimal:
Bei konstanter Temperatur vereinfacht sich die Gleichung zu:
Bei konstantem Umgebungsdruck folgt für die Änderung des osmotischen Drucks:
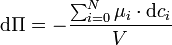 .
.
Der osmotische Druck Π ergibt sich somit mit den molaren Dichten ci aus der Veränderung aller chemischen Potentiale μ. In dieser Gleichung sind Mischungseffekte der beteiligten Substanzen berücksichtigt. Meist jedoch werden die Mischungseffekte der gelösten Stoffe untereinander und die Konzentration des Lösungsmittels vernachlässigt:
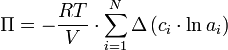 .
.
Eine weitere Näherung wäre die Vernachlässigung des Mischungseffektes des gelösten Stoffes mit dem Lösungsmittel. Hierbei wird die Aktivität ai dieser Stoffe als eins angenommen, so dass sich als grobe Näherung ergibt:
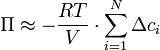 .
.
Diese überschlägige Berechnung kann für verdünnte Lösungen verwendet werden, führt jedoch bei höheren Konzentrationen zu Fehlern von mehr als 50%, insbesondere, da hier der Lösungseffekt nicht berücksichtigt wird.
Der negative Wert des osmotischen Drucks wird als osmotisches Potential ψo bezeichnet.
Osmometrie – Messung des osmotischen Drucks
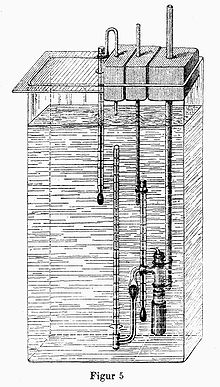 Pfeffersche Zelle - Prototyp des Membranosmometers.[10]
Pfeffersche Zelle - Prototyp des Membranosmometers.[10]Der osmotische Druck einer Lösung wird mit Membranosmometern ähnlich der Pfefferschen Zelle bestimmt. Dabei kann der Druck entweder statisch, nach Einstellung des Gleichgewichts, gemessen werden oder dynamisch, indem an das Steigrohrmanometer ein äußerer Druck angelegt wird, der den osmotischen Fluss gerade unterbricht.
Eine 1 molale Lösung von Rohrzucker (molare Masse 342,30 g·mol−1) in Wasser bewirkt bei Raumtemperatur bereits einen osmotischen Druck von 2,70 MPa (27 bar). Für erheblich höhere Drücke (mehrere 100 bar) lassen sich Messprinzipien wie die Änderung des Brechungsindex von Wasser oder der piezoelektrische Effekt einsetzen.[4]
Durch Messung des osmotischen Drucks beziehungsweise Potenzials ist es möglich, die mittlere Molekülmasse von Makromolekülen zu bestimmen; dieses Verfahren wird als Osmometrie bezeichnet. Da zwischen dem osmotischen Druck und den anderen kolligativen Eigenschaften einer Lösung, dies sind Siedepunktserhöhung und Gefrierpunktserniedrigung, ein unmittelbarer Zusammenhang besteht, kann durch deren Messung indirekt der osmotische Druck als osmotischer Wert ermittelt werden.
Während die direkte Messung des osmotischen Drucks die Anwesenheit zweier Phasen und einer spezifisch permeablen Membran erfordert, benötigen die indirekten Verfahren der Osmometrie lediglich die zu messende Lösung. Sie eignen sich daher besonders gut für die Charakterisierung unterschiedlicher Lösungen hinsichtlich ihrer osmotischen Eigenschaften. Osmolarität und Osmolalität geben die Konzentration gelöster Teilchen bezogen auf das Volumen oder die Stoffmenge an. Isoosmotisch sind Lösungen, deren osmotischer Wert gleich ist. Da der osmotische Wert keine Aussage über die Komponenten in den miteinander zu vergleichenden Lösungen beinhaltet, kann isoosmotisch nicht mit isotonisch gleichgesetzt werden.
Die selektiv permeable Membran
Wesentliches Element der Osmose ist die selektiv durchlässige (oder semipermeable) Membran; sie bestimmt, welche Stoffe passieren können. Durch diese Eigenschaft wird der erreichbare Gleichgewichtszustand des Systems festgelegt. Zugleich beeinflusst sie über die Diffusionsgeschwindigkeit der durchgelassenen Stoffe das dynamische Verhalten des Systems.[3]
Traubesche und Pfeffersche Zelle
- → Hauptartikel: Pfeffersche Zelle
Künstliche Membranen erzeugte zuerst der Privatgelehrte Moritz Traube aus Kaliumhexacyanidoferrat(II) (gelbes Blutlaugensalz), das in verdünnter Kupfersulfatlösung eine Haut aus Kupfercyanoferrat-II (siehe auch Kupfer-Nachweis) bildet. Diese ist nur für Wasser durchlässig. Das durch Osmose einströmende Wasser zerreißt die Haut, wodurch das eingeschlossene Kaliumhexacyanoferrat(II) wieder freigesetzt wird und sich erneut osmotische Zellen bilden, die nach einiger Zeit wiederum zerreißen. Ähnlich verhalten sich Salze von Erdalkali- und Schwermetallen in Alkalisilikatlösungen (Wasserglas). Die dabei entstehenden Gebilde sind unter der Bezeichnung osmotischer oder chemischer Garten bekannt.[29]
1877 gelang es Wilhelm Pfeffer diese mechanisch instabilen Niederschlagsmembranen auf das poröse Wandmaterial von Tonzellen aufzubringen und sie so zu stabilisieren. Er benutzte die Pfeffersche Zelle als Osmometer und konnte so den osmotischen Druck erstmals quantitativ bestimmen.
Mechanismen der Selektivität
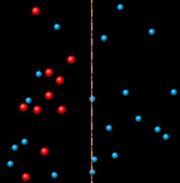 Osmose durch eine semipermeable Membran. Die größeren roten Teilchen können die Poren der Membran nicht durchdringen.
Osmose durch eine semipermeable Membran. Die größeren roten Teilchen können die Poren der Membran nicht durchdringen.Bei dem in der Abb. rechts dargestellten Membranmodell resultiert die selektive Durchlässigkeit aus der maximalen Porenweite. Die größeren roten Teilchen können die Membran nicht passieren, während die kleineren blauen Teilchen ungehindert von einer Seite zur anderen wechseln. Die für die Osmose entscheidende selektive Permeabilität kann aber auch auf anderen Mechanismen beruhen. In vielen Fällen spielen Lösungsvorgänge eine Rolle, so bei der von Nollet verwendeten Schweinsblase.
Auch katalytische Eigenschaften des Membranmaterials können für die selektive Permeabilität verantwortlich sein. Wenn ein Gemisch von Stickstoff (N2) und Wasserstoff (H2) durch eine dünne Folie aus Palladium von reinem Stickstoff getrennt wird, tritt Osmose auf; der Wasserstoff bewegt sich zu der wasserstoffärmeren Seite. Die Wasserstoff-Moleküle werden an der Palladiumoberfläche katalytisch in atomaren Wasserstoff gespalten, der anschließend durch die Palladiumfolie diffundiert.[4]
Letztlich kann selektive Permeabilität an Grenzflächen auftreten, die keine Membranen im engeren Sinne sind. Ein Beispiel dafür ist die Elektroosmose.
Osmose in der Biologie
Die Osmose hat eine besondere Bedeutung für biologische Systeme. Jede Zelle ist von einer Membran umgeben, die eine Barriere für den ungehinderten Stofftransport darstellt, für das Lösungsmittel Wasser aber durchlässig ist. Zugleich besitzt sie mehrere Zellorganellen mit selektiv permeablen Membranen. Viele Zellen stehen im Austausch mit Wasser, insbesondere Pflanzenzellen, die für die Aufnahme, den Transport und die Abgabe von Wasser zuständig sind. Zellen von Wirbeltieren sind von Blut und Lymphe umgeben.
Biomembranen
→ Hauptartikel Biomembran
Biomembranen sind für zahlreiche Stoffe selektiv durchlässig. Die Trägersubstanz einer Biomembran ist die Lipiddoppelschicht; sie ist nahezu undurchlässig für Wasser und darin gelöste Substanzen. Eingebettet in die Lipidschicht sind zahlreiche Transmembranproteine, die auf unterschiedliche Weise den Transport von Wasser und gelösten Teilchen durch die Membran hindurch ermöglichen. Zu den passiven gehören, neben porenbildenden Proteinen wie Ionenkanälen und Aquaporinen (Wasserkanäle), sogenannte Cotransporter. Aktiv arbeitende Transportproteine wie Protonenpumpen befördern Substanzen unter Verbrauch von Energie (meist aus der Hydrolyse von Adenosintriphosphat gewonnen) gegen ein vorhandenes Konzentrationsgefälle. Die Tätigkeit dieser Proteine kann das chemische Potenzial des Lösungsmittels Wasser inner- und außerhalb der Zelle oder seiner Organellen aktiv beeinflussen.
Die Durchlässigkeit oder Transportrate zahlreicher passiver Membranproteine kann durch zelluläre Mechanismen reguliert werden (zum Beispiel dem Gating von Ionenkanälen); dadurch ist es der Zelle möglich, auf Änderungen des umgebenden Milieus zu reagieren und so den osmotischen Stofftransport zu beeinflussen.
Wasserpotenzial
→ Hauptartikel: Wasserpotenzial
Zur Beschreibung des Wasserhaushalts von biologischen Systemen hat sich das Konzept des Wasserpotenzials als nützlich erwiesen. Es beschreibt den Saugwert einer Lösung für Wasser. Es werden lediglich Potenzialdifferenzen betrachtet, das Wasserpotenzial von reinem Wasser hat bei Standardbedingungen den Wert 0. Für das Wasserpotenzial einer Lösung gilt:
Ψ hat die Dimension Energie · Volumen-1 oder Kraft · Fläche-1 und wird in der Einheit Pascal angegeben. In obenstehender Gleichung ist p der hydrostatische Druck, − Π das osmotische Potenzial sowie der Ausdruck
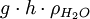 das Gravitationspotenzial, das bei der Betrachtung einzelner Zellen vernachlässigt werden kann.[30]
das Gravitationspotenzial, das bei der Betrachtung einzelner Zellen vernachlässigt werden kann.[30]Das Wasserpotenzial einer Lösung ist somit die Summe mehrerer Teilpotenziale. Dabei beschreibt das osmotische Potenzial ψo = − Π den Anteil des osmotischen Drucks am Saugwert der Lösung.
Wassertransport in Pflanzen
Pflanzen befördern Flüssigkeiten aus dem Wurzelbereich bis in die Spitzen. Durch die Osmose wird der sogenannte Wurzeldruck aufgebaut, der zusammen mit dem Transpirationssog und den Kapillarkräften die benötigte Druckdifferenz zum Wassertransport gegen die Schwerkraft bereit stellt. Bei diesem Transportprozess dominiert meist jedoch der Transpirationssog, da dieser deutlich höhere Drücke als der Wurzeldruck erreicht.[31] Nach den Erkenntnissen eines Teams der Universität Nordarizona können Bäume maximal 130 Meter hoch werden, da bei dem dort auftretenden Unterdruck die Kohäsion (Chemie) der Wassermoleküle nicht mehr ausreicht Kavitation in dem transportierenden Gewebe (Xylem) zu verhindern.[32]
Osmoregulation
 Wirkung des osmotischen Drucks auf Erythrozyten
Wirkung des osmotischen Drucks auf Erythrozyten Osmotische Zustände einer Pflanzenzelle (Plasmolyse)
Osmotische Zustände einer Pflanzenzelle (Plasmolyse)→ Hauptartikel Osmoregulation
Wird die osmotische Resistenz roter Blutkörperchen durch Einbringen in destilliertes Wasser (stark hypotones Medium) überschritten, nehmen sie unkontrolliert Wasser auf, bis sie schließlich platzen. Ihre Zellmembranen können nur geringen Drücken standhalten. Pflanzenzellen dagegen sind von einer stützenden Zellwand umgeben, wodurch sie erheblich höhere Innendrücke (→ Turgor) aushalten.
Zahlreiche Organismen wie Salzpflanzen, Halophile sowie Süßwasserbewohner leben in Umgebungen, deren osmotischer Wert stark von dem im Körper- oder Zellinneren abweicht. Ohne wirksame Osmoregulation würde der Organismus entweder austrocknen oder unkontrolliert Wasser aufnehmen. In Süßwasser lebende Einzeller verfügen über eine kontraktile Vakuole, die zunächst Wasser aus dem Cytoplasma aufnimmt und es danach aus der Zelle hinausschleust. Halophile (Salzbewohner) haben eine Reihe von Strategien entwickelt, um mit dem Salzüberschuss umzugehen. Dazu gehört die Ausbildung spezieller Organe wie Salzdrüsen oder Nieren zur Salzausscheidung, Mechanismen zur Salzeinlagerung oder die Anreicherung osmotisch aktiver Substanzen (Osmolyte) in den Zellen.
Nieren finden sich auch bei allen Wirbeltieren. Sie dienen der Ausscheidung sogenannter harnpflichtiger Substanzen, darunter überschüssige Elektrolyte und Glucose, die sonst zu einem Ansteigen des osmotischen Werts im Körper führen würden.
In Säugetierzellen wurde das Protein NFAT5, (ein Transkriptionsfaktor, auch bekannt als Tonicity-Responsive Enhancer Binding Protein, TonEBP) gefunden, das verstärkt exprimiert (synthetisiert) wird, wenn der osmotische Druck ansteigt. Es setzt eine Reihe von Gegenregulationsmechanismen in Gang, um die Zelle vor hypertonem Stress zu schützen. Dazu gehört die Anreicherung von Osmolyten in der Zelle. Beispiele für solche Substanzen sind myo-Inositol, Betain und Taurin, für die jeweils eigene Transportproteine existieren.[25]
Osmotische Arbeit
Im Experiment mit dem U-Rohr (siehe Abschnitt Grundlagen) wurde die rechte Flüssigkeitssäule gegen die Schwerkraft angehoben. Das zeigt anschaulich, dass in einer osmotischen Zelle Arbeit verrichtet werden kann.
Das Konzept der Energiegewinnung durch osmotische Arbeit wird als Osmosekraftwerk bezeichnet, siehe auch Abschnitt Anwendungen und Beispiele.
In den Biowissenschaften und der Medizin wird unter osmotischer Arbeit[33] (engl. osmotic Work) oft die Energiedifferenz zwischen den osmotischen Potenzialen eines Systems (zum Beispiel einer Zelle) verstanden. In diesem Sinne verrichtet die Zelle osmotische Arbeit, wenn Stoffe aktiv, unter Energieverbrauch gegen ein Konzentrationsgefälle transportiert werden. Andererseits kann die, aus der Differenz osmotischer Potenziale resultierende, osmotische Energie von der Zelle für energieverbrauchende Prozesse genutzt werden, beispielsweise bei der chemiosmotischen Kopplung.
Anwendungen und Beispiele
Osmolaritätsmessung, isoosmolare Puffer und Lösungen
Die Bestimmung der Osmolarität von Lösungen mittels osmometrischer Methoden gehört in vielen Bereichen der Biowissenschaften zum Laboralltag. Bei der Arbeit mit lebenden Zellen ist eine isotone Pufferlösung (wie Ringer) oft unabdingbar, um unerwünschte Reaktionen der Zellen durch osmotischen Stress zu vermeiden. Insbesondere bei der Isolation von Protoplasten würde ein hypoosmolarer Puffer die von keiner Zellwand mehr geschützten Zellen platzen lassen. Bei der Herstellung solcher Lösungen im Labor kann zur Kontrolle die tatsächliche Osmolarität gemessen und mit dem erwarteten Wert verglichen werden.
In der Medizin wird für Infusionen eine isotonische Kochsalzlösung eingesetzt, um eine Schädigung der Körperzellen durch osmotischen Druck zu vermeiden. Es handelt sich dabei um ein Gemisch aus Wasser mit 0,9 % (Massenprozent) Kochsalz, die Osmolarität dieser Lösung entspricht mit 308 mosmol/l annähernd der des Blutplasmas, dies entspricht einem osmotischen Druck von 0,7 MPascal. Würde bei Infusionen reines Wasser anstatt einer isotonen Lösung benutzt werden, könnte ein solcher Druckunterschied die Blutzellen platzen lassen.
Dialyse
- → Hauptartikel: Dialyse und Dialyse (Chemie)
Bei der Dialyse werden Membranen eingesetzt, die Moleküle und Ionen unterhalb einer bestimmten Größe oder Molekularmasse durchlassen und Makromoleküle wie Proteine oder Nukleinsäuren zurückhalten. Mit diesem Verfahren können niedermolekulare Substanzen und Ionen gezielt aus einer Lösung entfernt oder deren Konzentration auf den Wert einer vorgegebenen Lösung angeglichen werden. Dazu wird je nach Anwendung die zu dialysierende Lösung in ein geeignetes Gefäß (den Dialyseschlauch) gefüllt und in die äußere Dialyselösung getaucht, worin sie über längere Zeit verbleibt. Oder die zu reinigende Flüssigkeit steht, wie bei der Hämodialyse, über eine semipermeable Membran in Kontakt mit einer Spüllösung. Dialyseverfahren werden in der Medizin unter anderem für die Blutreinigung eingesetzt, sowie in der Chemie und Verfahrenstechnik (zum Beispiel bei der Herstellung von alkoholfreiem Bier).
Dichtegradientenzentrifugation
Bei der Dichtegradientenzentrifugation von lebenden Zellen oder deren Bestandteilen können Stoffe wie Saccharose als Träger des Dichtegradienten oft durch hochmolekulare Substanzen mit nur geringer osmotischer Aktivität (niedriger osmotischer Wert) ersetzt werden, um die Zellen während der Zentrifugation nicht osmotischem Stress auszusetzen.
Umkehrosmose
- → Hauptartikel: Umkehrosmose
Bei der Umkehrosmose (auch reverse Osmose oder Antiosmose genannt) wird durch Anwendung von Druck eine Substanz gegen ein Konzentrationsgefälle aufkonzentriert. Dieses Verfahren wird insbesondere für die Aufbereitung (zum Beispiel Entsalzung) von Trinkwasser eingesetzt.
Osmosekraftwerk
- → Hauptartikel: Osmosekraftwerk
Bei dem Konzept des Osmosekraftwerks wird die osmotische Arbeit zur Energiegewinnung eingesetzt. Das Kraftwerk nutzt dazu die Unterschiede im chemischen Potential zwischen salzhaltigem Meer- und Süßwasser aus, um damit Turbinen für die Stromgewinnung zu betreiben. Über eine Membran strömt vorgereinigtes Süßwasser in eine Leitung mit Salzwasser und erhöht so den osmotischen Druck in dieser Leitung. Mit einem Teil des so entstehenden Brackwassers werden Turbinen angetrieben während der größere Teil (2/3) über einen Druckaustauscher den Druck des frisch zuströmenden Salzwassers erhöht. [34] Osmosekraftwerke sind bislang noch nicht im kommerziellen Einsatz; Prototypen mit einer Leistung von bis zu drei Megawatt werden bereits seit einigen Jahren entwickelt. [35]
Osmose im Alltag
- Bei der Konservierung von Lebensmitteln durch Einzuckern oder Pökeln wird das enthaltene Wasser durch Osmose entzogen, da die Konzentration von Zucker oder Salz außen sehr viel höher als im Inneren des Lebensmittel ist. Vorhandene Mikroorganismen können sich nicht mehr vermehren und daher auch nicht mehr zersetzend wirken. Auf diese Weise haltbar gemachte Lebensmittel verändern sich dabei durch den Wasserentzug oft drastisch.
- Beim Kochen von Gemüse wird dem Wasser Salz zugefügt, um den Einstrom von Wasser in das (leicht salzhaltige) Gemüse und den damit verbundenen Geschmacksverlust zu verhindern.
- Ein mit Salatsoße angemachter Blattsalat verliert nach relativ kurzer Zeit seine Festigkeit (Turgor). Diese erhält er normalerweise durch das in den Zellen vorhandene Wasser, welches durch Osmose an die Salatsoße abgegeben wird.
- Das Aufplatzen reifer Früchte nach einem Regen wird durch den osmotischen Einstrom des Regenwassers und den daraus resultierenden osmotischen Druck im Innern der Frucht bewirkt.
Siehe auch
Einzelnachweise
- ↑ David C. Guell, Howard Brenner. Physical Mechanism of Membrane Osmotic Phenomena. (1996) doi:10.1021/ie950787f
- ↑ a b Frank G. Borg. What is Osmosis arXiv.org, e-Print (2003)
- ↑ a b A. V. Raghunanthan and N. R. Aluru, Molecular Understanding of Osmosis in Semipermeable Membranes. (2006) doi:10.1103/PhysRevLett.97.024501, University of Illinois (PDF, 616 kB)
- ↑ a b c d e f Moore, Hummel: Physikalische Chemie. Verlag de Gruyter 1986, ISBN 3-11-010979-4
- ↑ H. Dutrochet. Nouvelles Recherches sur l'Endosmose et l'Exosmose, suivies de l'application expérimentales de ces actions physiques à la solution du problême de l'irritabilité végétale. Paris, 1828
- ↑ J.H. van `t Hoff (1887), "The role of osmotic pressure in the analogy between solutions and gases" Website Uri Lachish (engl., PDF). Zeitschrift für physikalische Chemie 1, 481-508
- ↑ J.H. van ’t Hoff. Osmotical pressure and chemical equilibrium. 1901. Nobel Lecture (PDF)
- ↑ a b Albert Einstein: Über die von der molekularkinetischen Theorie der Wärme geforderte Bewegung von in ruhenden Flüssigkeiten suspendierten Teilchen, Annalen der Physik, 17, 1905 Universität Wien (PDF)
- ↑ Kortüm, Lachmann. Einführung in die chemische Thermodynamik. Vandenhoeck & Ruprecht 1981, S. 245,“ Fußnote 3.
- ↑ a b Wilhelm Pfeffer. Osmotische Untersuchungen – Studien zur Zellmechanik. Engelmann Leipzig . 1877. bei [www.archive.org]
- ↑ David L. Nelson, Michael M. Cox. Lehninger - Principles of Biochemistry. 4.th Editon, S. 57. W.H.Freemann, New York 2005. ISBN 0-7167-4339-6
- ↑ Peter W. Atkins, Julio de Paula: Physikalische Chemie, S. 166. Wiley-VCH, 2005, ISBN 3-527-31546-2.
- ↑ Eduard Strasburger (Begründer), Peter Sitte (Bearbeiter), Elmar W. Weiler, Joachim W. Kadereit, Andreas Bresinsky, Christian Körner (Autoren): Lehrbuch der Botanik für Hochschulen, S. 254. 35. Auflage. Spektrum, Heidelberg, Berlin 2002, ISBN 3-8274-1010-X.
- ↑ Gerd Wedler. Lehrbuch der physikalischen Chemie, S. 318ff. Wiley VCH, 1989. ISBN 3-527-26702-6
- ↑ Ash R., Barrer R.M., Vernon A., Edge J., Foley T. Thermo-osmosis of sorbable gases in porous media. Part IV. J. Membr. Sc. 1997, 125:41–59
- ↑ Peter W. Atkins, Julio de Paula: Physikalische Chemie, S. 169. Wiley-VCH, 2005, ISBN 3-527-31546-2.
- ↑ Grosse W., Bücher H.B., Tiebel H. Pressurized ventilation in wetland plants. Acquatic Botany 1991, 39:89–98
- ↑ A.J.Staverman. The theory of measurement of osmotic pressure. Rec. Trav. Chim. 70:344, 1951.
- ↑ A. E. Hill. Osmosis in Leakey Pores: The Role of Pressure. Proc R. Soc. Lond. B 237, S. 363-367. 1989
- ↑ Adolf Fick. On liquid diffusion Philos. Mag., 10:30, 1855
- ↑ David Charles Guell. The Physical Mechanism of Osmosis and Osmotic Pressure; A Hydrodynamical Theroy for Calculating the Osmotic Reflection Coefficient. Doctoral thesis, Massachusetts Institute of Technology, 1991.
- ↑ A. Mauro. Nature of solvent transfer in osmosis. Science, 126:252, 1957
- ↑ Czihak G., Langer H., Ziegler H. Biologie, Ein Lehrbuch. 2. Auflage. Springer-Verlag Berlin 1978, ISBN 3-540-08273-5
- ↑ a b J.H. van ’t Hoff, The role of osmotic pressure in the analogy between solution and gases, Zeitschrift fur Physikalische Chemie 1 (1887), 481-508. Website Uri Lachish (engl., PDF)
- ↑ a b Stefanie Brookmann. Osmolyte und Osmolystrategien von humanen und murinen hämotopoetischen Stamm- und Progenitorzellen. Inauguraldissertation 2007 Deutsche Nationalbibliothek (PDF)
- ↑ Albert Einstein: Über die von der molekularkinetischen Theorie der Wärme geforderte Bewegung von in ruhenden Flüssigkeiten suspendierten Teilchen, Annalen der Physik, 17, 1905 Universität Wien (PDF)
- ↑ Chinard FP and Enns T. Osmotic pressure. Science 124: 472–474, 1956.
- ↑ F. Kiil. Kinetic model of osmosis through semipermeable and solute-permeable membranes (2002) doi:10.1046/j.1365-201X.2003.01062.x
- ↑ Sehon H. Physikalische Chemie. Verlag Herder Freiburg. i. Brsg. 1976, ISBN 3-451-16411-6
- ↑ Sitte P., Weiler E.W., Kadereit J.W., Bresinsky A., Körner C. Strasburger – Lehrbuch der Botanik, Kapitel Wasserpotential. Spektrum Gustav Fischer 2002, ISBN 3-8274-1010-X
- ↑ Erläuterung des Wurzeldrucks
- ↑ George W. Koch, Stephen C. Sillett, Gregory M. Jennings, Stephen D. Davis: The limits to tree height. Nature 428, 851–854, 2004, doi:10.1038/nature02417
- ↑ Lehninger, Nelson, Cox: Lehninger Biochemie. 3. Auflage. Springer-Lehrbuch, Berlin 2001, ISBN 3-540-41813-X
- ↑ Statkraft: Osmotic Power (PDF, engl.)
- ↑ Bericht der Financial Times Deutschland vom 23. Jan. 2008
Literatur
- Peter W. Atkins, Julio de Paula: Physikalische Chemie. Wiley-VCH, 2005, ISBN 3-527-31546-2.
- Moore Walter J., Hummel Dieter O.: Physikalische Chemie. Walter de Gruyter, Berlin 1986, ISBN 3-11-010979-4.
- Wagner R.H., Moore, H.D. Determination of Osmotic Pressure, in Physical Methods of Organic Chemistry, Hrsg. Weissengerber A. Part 1, 3rd ed, Interscience, New York 1959.
Weblinks
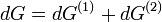
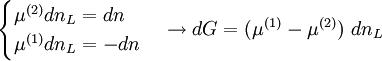


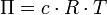
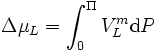
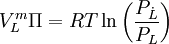
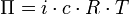
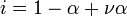
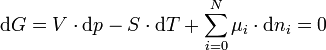
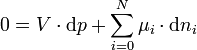

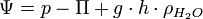


Wikimedia Foundation.