- Peniswurm
-
Priapswürmer 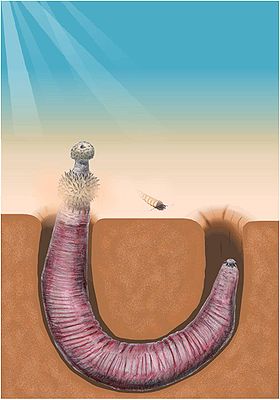
Zeichnerische Rekonstruktion des ausgestorbenen Priapuliden Ottoia aus dem Burgess-Schiefer
Systematik Unterreich: Vielzellige Tiere (Metazoa) Abteilung: Gewebetiere (Eumetazoa) Unterabteilung: Bilateria ohne Rang: Urmünder (Protostomia) Überstamm: Häutungstiere (Ecdysozoa) Stamm: Priapswürmer Wissenschaftlicher Name Priapulida Ordnungen - Priapulomorpha
- Halicryptomorpha
- Meiopriapulomorpha
- Seticoronaria
Als Priapswürmer (Priapulida) bezeichnet man einen Stamm wurmförmiger Häutungstiere (Ecdysozoa) mit verdicktem rüsselartigen Kopf, die allesamt im oder auf dem Meeresboden leben. Ihre nächsten Verwandten sind vermutlich Hakenrüssler (Kinorhyncha) und Korsetttierchen (Loricifera), mit denen sie in einem Taxon Scalidophora zusammengefasst werden. Sie sind wegen ihres an das männliche Geschlechtsorgan erinnernden Aussehens nach dem griechischen Gott der Manneskraft, Priapos, benannt, der gewöhnlich mit einem enormen Penis dargestellt wird. Sie wurden erstmals von Carl von Linné in seinem Werk Systema Naturae als Priapus humanus, übersetzt "Menschlicher Penis", erwähnt.
Inhaltsverzeichnis
Aufbau
Priapswürmer haben einen meist plumpen, zylindrisch geformten Körper, dessen Länge zwischen 0,05 Zentimetern bei Tubiluchus corallicola und 39 Zentimetern bei Halicryptus higginsi liegt. Obwohl von kreisförmigem Querschnitt, sind die inneren Organe zweiseitig symmetrisch angeordnet.
Äußere Form
Am Vorderende tragen die Tiere einen kurzen rüsselförmigen "Kopf", der auch als Proboscis oder Introvert bezeichnet wird. Dahinter befindet sich der oberflächlich in 30 bis 100 Ringe geteilte Rumpf, der jedoch intern nicht segmentiert ist. Er ist bedeckt von chitinhaltigen Dornen, den Scaliden, sowie von zahlreichen Grübchen und Papillen, die wohl allesamt als Sinnesrezeptoren dienen. Die Dornen werden darüber hinaus auch zur Fortbewegung eingesetzt. Besonders häufig treten sie, meist in Form dorniger Haken, auch auf dem Introvert auf, wo sie in mehreren Längsreihen angeordnet sind und wahrscheinlich auch zum Beutefang benutzt werden.
Am Hinterende können sich ebenfalls Haken befinden; sie dienen wohl zur Fortbewegung. Einige Arten besitzen zudem entweder einen langen, einziehbaren Schwanz, der vermutlich zur Verankerung im Sediment dient, oder ein bis zwei büschelige Schwanzanhänge. Letztere kommen besonders bei Tieren vor, die in sauerstoffarmen Sedimenten leben; sie dienen daher wahrscheinlich eher dem Gasaustausch, daneben vielleicht auch der Regulation des Salzhaushalts oder der Wahrnehmung chemischer Substanzen.
Haut, Muskulatur und Pseudocoelom
Die Körperwand ist ein Hautmuskelschlauch, der aus einer chitinhaltigen nicht-zelligen Außenhaut, der Cuticula, einer als Epidermis bezeichneten Innenhaut, sowie aus zwei Schichten quergestreifter Muskulatur besteht. Die Cuticula setzt sich zusammen aus einer außenliegenden kollagenhaltigen Epicuticula, einer aus Proteinen aufgebauten Exocuticula und einer chitinhaltigen Endocuticula und wird regelmäßig gehäutet. Die Epidermis besteht aus einer einlagigen Zellschicht; unter ihr befindet sich erst eine Schicht Ringmuskulatur, der eine weitere Schicht Längsmuskulatur folgt. Am Vorderende sitzen zusätzlich zwei Gruppen spezialisierter Längsmuskeln, die Introvert-Retraktor-Muskeln, die dem Wurm ermöglichen, das Introvert in den Rumpf einzuziehen und somit zu schützen.
Zwischen der innersten Muskelschicht und dem Verdauungstrakt samt assoziierter Muskeln befindet sich die Leibeshöhle. Sie wurde lange Zeit als echtes Coelom angesehen, also als flüssigkeitsgefüllter Hohlraum, der von Epithelgewebe begrenzt wird, das während der Embryonalentwicklung aus Zellen des Mesoderm hervorgeht. Es ist aber nach neueren Erkenntnissen wohl nicht von einer eigenen Zellschicht umkleidet und stellt damit ein so genanntes Pseudocoelom dar; eine Ausnahme bilden lediglich die Keimdrüsen und vielleicht auch der Schlund, die anscheinend von einem echten Coelom umgeben sind. Das Pseudocoelom ist von einer Flüssigkeit gefüllt, in der neben amöboiden Fresszellen auch rosafarbene Blutzellen zirkulieren, die den sauerstoffbindenden Blutfarbstoff Hämerythrin enthalten und deshalb als Hämerythrocyten bezeichnet werden. Sie erlauben den Würmern vermutlich den Aufenthalt in ihrem teilweise sauerstofflosen (anoxischen) Lebensraum. Neben seiner Funktion in der Weiterleitung von Nährstoffen und Gasen durch den Körper dient das Pseudocoelom auch als hydrostatisches Stützskelett.
Verdauungs- und Ausscheidungsorgane
Der Verdauungstrakt beginnt mit dem Mund, an den sich ein muskulöser und von innen mit zahlreichen "Zähnchen" besetzter Schlund anschließt. Er kann ein- und ausgestülpt werden und wird in letzterem Zustand auch als Mundkegel bezeichnet - die Zähnchen kommen dann außen zu liegen. Hinter dem Schlund schließt sich manchmal eine weitere Höhlung an, das Polythyridium, das wohl als Muskelmagen der weiteren Zerkleinerung der Nahrung dient, die dann im mit zahlreichen Einstülpungen, den Microvilli, besetzten Mitteldarm aufgenommen wird. Nicht verwertbare Reststoffe gelangen in einen kurzen Enddarm, der im endständigen After endet. Der gesamte Verdauungstrakt ist von zwei Muskelschichten umgeben, innenliegender Ringmuskulatur und außenliegender Längsmuskulatur.
Die Ausscheidung flüssiger Abfallstoffe findet durch zwei paarig angelegte büschelige Organe statt, die als Protonephridien bezeichnet werden und im hinteren Rumpf links und rechts des Darmes an speziellen Bändern, den Mesenterien, aufgehängt sind. Sie bestehen aus feinen Röhrchen, an deren Ende mindestens zwei einfach bewimperte Zellen, die Solenocyten, sitzen. Zusammen mit dem von der zylindrischen Keimdrüse ausgehenden Ei- beziehungsweise Samenleiter münden sie in einem gemeinsamen Urogenitaltrakt, der durch kleine Öffnungen, die Nephridioporen, am Rumpfende mit der Außenwelt verbunden ist.
Nervensystem und Sinnesorgane
Das Nervensystem besteht aus einem Nervenring, der am Vorderende des Kopfes um den Mund herum verläuft. Von ihm zieht ein einfacher, ungepaarter und ganglienloser Nervenstrang in der Mitte der Bauchseite nach hinten, von dem seinerseits in regelmäßigen Abständen Ringnerven abzweigen. Er selbst endet in einem Schwanzganglion.
Der Sinneswahrnehmung dienen die zahllosen Grübchen (Flosculi), Noppen (Papillen) und Dornen (Scaliden), die hohl sind und jeweils eine einfach begeißelte Sinnesnervenzelle beherbergen.
Verbreitung und Lebensraum
Die größeren Priapswurm-Arten leben nur in den kalten zirkumpolaren Gewässern der Arktis und Antarktis, an den Küsten Nordamerikas östlich bis etwa auf die Höhe des US-Bundesstaats Massachusetts, westlich bis etwa Zentralkalifornien, sowie in Nord- und Ostsee; von Süden auch um das argentinische Patagonien herum. Die kleineren Arten, besonders aus der Familie Tubiluchidae, finden sich dagegen weltweit in marinen Gewässern, auch in den Tropen, speziell in der Karibik und vor den Küsten Mittelamerikas.
Alle Tiere leben benthisch, also auf dem Meeresboden, wo sie sich derart in sauerstoffarme Sedimente eingraben, dass die Mundöffnung auf der Sedimentoberfläche liegt. Sie kommen von der Gezeitenzone bis in etwa 500 Meter Wassertiefe vor; manche Arten finden sich im Wattenmeer oder im brackigen Wasser vor Flussmündungen. Die kleineren Arten leben auch interstitial in den Lücken zwischen den feinen Körnern von Muschel- oder Korallensand.
Eine besondere ökologische Nische hat die Art Halicryptus spinulosus gefunden, die in nahezu sauerstofffreien (anoxischen), sulfidgesättigten Sedimenten der Ostsee lebt und offensichtlich in der Lage ist, große Konzentrationen dieses Gifts zu tolerieren beziehungsweise innerhalb des eigenen Körpers abzubauen.
Insbesondere die größeren Wurmarten sind heute in erster Linie auf arten- und damit konkurrenzarme Lebensräume spezialisiert und spielen ökologisch im Vergleich zu anderen Bewohnern des Meeresbodens nur noch eine untergeordnete Rolle.
Ernährung und Fortbewegung
Die größeren Arten leben allesamt räuberisch von wirbellosen Tieren mit weichem Körper wie Vielborstern (Polychaeta), aber auch anderen Priapswürmern. Ihrer Beute lauern sie im Sediment eingegraben auf, packen sie mit den dornigen Haken ihres Introverts und schieben sie als Ganzes durch permanentes Ein- und Ausstülpen des Mundkegels immer weiter in den Schlund, wo sie durch die feinen Zähnchen klein gehäckselt wird. Beim Ausstülpen des Mundkegels gelangen die Zähnchen nach außen und helfen so mit, die Beute sicher zu ergreifen.
Die kleineren Arten ernähren sich dagegen eher von organischem Abfall und den darin enthaltenen Bakterien. Maccabeus filtert eventuell Nährstoffe aus dem Wasser, erzeugt dazu allerdings anscheinend keinen Atemwasserstrom wie andere Filtrierer. Ausgekleidete Wohnröhren, wie sie von anderen meereslebenden Würmern angelegt werden, sind bei den Priapswürmern unbekannt.
Priapswürmer bewegen sich mit Hilfe ihres rüsseligen Introverts vorwärts und können sich, wenn auch bei den erwachsenen Tieren mit Schwierigkeiten, in weiche Sedimente eingraben, indem sie dort abwechselnd ihre vordere und hintere Körperregion verankern.
Zu Beginn eines Zyklus wird der Körper am Hinterende durch Entspannung der dortigen Ringmuskulatur verdickt und bildet dort auf diese Weise den so genannten Penetrationsanker. Die restliche Ringmuskular kontrahiert hingegen und verringert damit den Körperquerschnitt. Da die Flüssigkeit im Pseudocoel praktisch immer dasselbe Volumen einnimmt, stülpen sich bei entspannter Längsmuskulatur Introvert und Schlund nach vorne aus. Dadurch, dass nun durch eine vom Hinterende ausgehende wellenförmige Kontraktion der Ringmuskulatur immer mehr Flüssigkeit aus dem Rumpf in das Introvert gelangt, dehnt sich dieses erheblich aus und verankert nun seinerseits den Körper an der Vorderseite. Durch Kontraktion der Längsmuskulatur und der Introvert-Retraktormuskeln wird der Rest des Körpers nach vorne nachgezogen, so dass nach Aufbau eines neuen Penetrationsankers am Hinterende der Zyklus von vorne beginnen kann.
Fortpflanzung und Entwicklung
Priapswürmer sind getrenntgeschlechtliche Tiere, obwohl sich Männchen und Weibchen meist nicht unterscheiden lassen. In der Gattung Maccabeus sind keine Männchen bekannt, sie vermehrt sich daher wahrscheinlich parthenogenetisch, also ohne Befruchtungsvorgang.
Samen und die kleinen Eier mit relativ hohem Dottergehalt werden bei den größeren Arten meist im Spätwinter freigesetzt und extern befruchtet. Meist geben zuerst die Männchen, dann die Weibchen ihre Keimzellen ins Meerwasser ab. Bei den kleineren Arten kommt eventuell auch eine interne Befruchtung im Körper des Weibchens vor.
Die Entwicklung der Tiere verläuft manchmal direkt, meist aber über ein ebenfalls bodenlebendes Larvenstadium. Der Rumpf der Larven ist von einem Korsett, der Lorica, aus zehn Cuticulaplatten bedeckt, von denen je eine bauch- und rückenseitig, drei auf jeder Seite und zwei kleinere am Vorderende liegen. Wie bei den erwachsenen Tieren ist auch bei den Larven das Vorderende als Introvert ausgebildet und in den Rumpf einstülpbar. Am Hinterende befinden sich spezielle "Zehen", die mit Klebedrüsen versehen sind und wahrscheinlich der Anheftung am Sediment dienen. Vor der Metamorphose, also der Umwandlung zum erwachsenen Tier, durchläuft die Larve zahlreiche Häutungen, bei denen auch die Lorica jeweils erneuert wird. Während der komplexen Entwicklungsphase, die womöglich bis zu zwei Jahre andauern kann, ernährt sie sich vermutlich detrivor, also von organischen Abfallstoffen. Am Ende dieser Zeit steht die Metamorphose selbst, bei der die Lorica verloren geht; Häutungen finden jedoch auch bei den erwachsenen Tieren weiterhin statt.
Stammesgeschichte
Vergleiche mit modernen Taxa sehen die nächsten Verwandten der Priapswürmer recht eindeutig in den Hakenrüsslern (Kinorhyncha) und Korsetttierchen (Loricifera), mit denen sie das Taxon Scalidophora bilden. Die drei Gruppen teilen zahlreiche Merkmale, so die mit Chitin verstärkte Außenhaut, die darauf befindlichen chitinhaltigen Borsten oder Stacheln, der Sinneswahrnehmung dienende Grübchen (Flosculi) sowie zwei Gruppen von Introvert-Retraktor-Muskeln, die vorne am Gehirn ansetzen.
Welche der beiden Tierstämme die evolutionäre Schwestergruppe darstellt, ist hingegen weitaus stärker umstritten; alle drei kombinatorisch möglichen Varianten sind von Zoologen vorgeschlagen und begründet worden. Für eine engere Verwandtschaft von Priapswürmern und Korsetttierchen spricht das Vorhandensein eines von der Cuticula gebildeten Korsetts, das bei den ersteren im Larvenstadium vorhanden ist, für eine enge Verwandtschaft von Priapswürmern und Hakenrüsslern dagegen die Tatsache, dass das Schlundgewebe nicht aus Epithelmuskelzellen besteht, sondern sich von embryonalem Mesoderm ableitet. Die dritte Alternative, ein Schwestertaxon-Verhältnis zwischen Hakenrüsslern und Korsetttierchen mit den Priapswürmern als Außengruppe, wird durch den vorstreck-, aber nicht ausstülpbaren Mundkegel der beiden ersteren Taxa begründet.
Seit dem Jahr 2004 sind aus Hunan in Südchina Embryo-Fossilien der Art Markuelia hunanensis bekannt. Sie entstammen der erdgeschichtlichen Epoche des mittleren bis späten Kambriums vor etwa 500 Millionen Jahren und werden durch eine kladistische Analyse als Vertreter der Stammlinie der Scalidophora angesehen, lassen sich also keiner der modernen drei Gruppen zuordnen, aus denen dieses Taxon besteht. Markuelia hunanensis war möglicherweise segmentiert - falls sich dieser Befund und zugleich die kladistische Analyse bestätigen sollte, wäre der Verlust der Segmentierung ein gemeinsames abgeleitetes Merkmal (Synapomorphie) sowohl der Priapswürmer als auch der Korsetttierchen und würde damit deren Schwestergruppenverhältnis unterstreichen.
In die weitere Verwandtschaft der Priapswürmer gehören Faden- (Nematoda) und Saitenwürmer (Nematomorpha), mit denen die Scalidophora das Taxon Cycloneuralia bilden. Allesamt werden sie in die Häutungstiere (Ecdysozoa) eingeordnet, zu denen auch die Panarthropoda mit den Gliederfüßern (Arthropoda) als wichtigster Gruppe gerechnet werden.
Fossile Überlieferung
Anders als die meisten weichkörperigen Tiere sind die Priapswürmer auch fossil bekannt; bisher wurden mindestens elf Arten beschrieben, die sich formell auf sieben Gattungen aufteilen. Sie finden sich schon im kanadischen Burgess-Schiefer, der vor 530 Millionen Jahren in der erdgeschichtlichen Epoche des mittleren Kambriums entstand, sind also schon von Anbeginn des modernen Äons, des Phanerozoikums, erhalten.
Priapswürmer waren neben den Gliederfüßern (Arthropoda) die bedeutendste Wirbellosen-Gruppe des Kambriums und stellten bis zum Ordovizium die dominanten Raubwürmer des Meeresbodens. Erst mit dem Auftreten kieferbewehrter Vielborster (Polychaeta) aus den Reihen der Ringelwürmer (Annelida) verloren sie an ökologischer Bedeutung und verschwanden bis zum Silur weitgehend aus der fossilen Überlieferung, weswegen Priapswürmer auch schon als "basic failure", also als (evolutionärer) Fehlschlag gesehen wurden. Lediglich eine weitere fossile Gattung, Priapulites, konnte aus der späteren erdgeschichtlichen Epoche des Karbon beschrieben werden; sie lässt sich bereits in die moderne Familie Priapulidae einordnen.
Die fossil erhaltenen Arten lebten wie ihre modernen Verwandten räuberisch als Bestandteil des Benthos, also des weichen schlammigen Meeresbodens. Die acht Zentimeter lange Art Ottoia prolifica, deren Häufigkeit schon im wissenschaftlichen Namen zum Ausdruck kommt, lauerte Hyolithiden (Hyolitha) auf, seltsamen, wurmartigen, heute ausgestorbenen Tieren des Paläozoikums, die manchmal zu den Weichtieren (Mollusca) gestellt werden, deren tatsächliche Verwandtschaftsverhältnisse und Lebensweise aber bis heute praktisch unbekannt sind. Reste von Artgenossen im Darm von Ottoia prolifica deuten darüber hinaus auf Kannibalismus hin. Neben Ottoia verfügten auch Arten der Gattungen Corynetis und Anningvermis aus dem Maotianshan-Schiefer Chinas bereits über Schlundzähnchen, sie wiesen zudem schon Schwanzanhänge auf.
Stammesgeschichtlich bilden die fossilen Arten (mit Ausnahme von Priapulites) vermutlich die Schwestergruppe der modernen Arten, wie eine systematische Analyse ergab. Die Arten Corynetis brevis, Anningvermis multispinosus, Acosmia matiania, Paraselkirkia sinica und Xiaoheiquingella peculiaris wurden allerdings bisher nicht in diese mit einbezogen:
Priapswürmer (Priapulida)
│ ?────Anningvermis, Corynetis, Acosmia, Paraselkirkia, Xiaoheiquingella ├─┬────(Fossile Formen) │ ├────†Selkirkia │ ├────N. N. │ ├─┬──N. N. │ │ ├──†Ottoia │ │ ├──N. N. │ │ ├──† Scolecofurca │ │ └──† Louisella │ │ │ ├────†Paleoscolidea │ ├────†Cricocosmia │ ├────N. N. │ ├────†Palaeoscolex │ └────†Maotianshania │ └──────(Moderne Formen + † Priapulites)
Die als Paleoscolidea bezeichneten Gattungen wurden lange Zeit nicht zu den Priapswürmern gerechnet und mit verschiedenen anderen Tiergruppen alliiert; nach moderner Auffassung gibt es aber keinen Grund, sie von den anderen fossilen Formen systematisch besonders zu unterscheiden. Zwei traditionell zu den Priapswürmern gerechnete Gattungen, Ancalagon und Fieldia, gehören auf der anderen Seite wahrscheinlich nicht zu dieser Gruppe, sondern sind möglicherweise Vertreter der Stammlinie aller Scalidophora.
Priapswürmer als paläontologische Modellorganismen
Aufgrund des vergleichsweise häufigen Vorkommens fossiler Priapswürmer in kambrischen Sedimenten eignen sich diese gut für die Untersuchung makroevolutionärer Fragestellungen. Ein wichtiges Problem in Bezug auf die Evolutionsgeschichte der vielzelligen Tiere (Metazoa) betrifft zum Beispiel die Natur der so genannten Kambrischen Radiation. Dabei handelt es sich um eine nur wenige Millionen Jahre lange Zeitspanne im Kambrium, innerhalb derer erstmals zahlreiche moderne Taxa mit ihren morphologisch zum Teil erheblich verschiedenen Grundplänen auftraten. Neben der Frage nach der Ursache dieser plötzlichen "Explosion" ist man heute auch daran interessiert, ob die morphologische Vielfalt innerhalb eines gegebenen Taxons damals größer war als in der Gegenwart. Hintergrund dieser Fragestellung ist die von einigen Paläontologen geäußerte Vermutung, dass das Kambrium in der Tierwelt eine Zeit des "Experimentierens" mit verschiedenen Körperbauplänen war, von denen sich in der Folge nur einige wenige durchsetzen konnten. Untersuchungen an fossilen Gliederfüßern stützen diese These bedingt, liefern allein jedoch nicht notwendigerweise ein repräsentatives Bild.
Zu diesem Zweck wurden die erhaltenen Priapswurmfossilien phänetisch analysiert: Im Gegensatz zur kladistischen Analyse, die der Aufdeckung der stammesgeschichtlichen Verwandtschaftsverhältnisse dient und bei der Bildung höherer Taxa nur gemeinsame abgeleitete Merkmale heranziehen darf, werden dabei auch die "primitiven", also von einem gemeinsamen Vorfahren stammenden Charakteristika in die Untersuchung mit einbezogen; sie eignet sich daher besonders für quantitative Vergleiche der morphologischen Typenvielfalt zweier Gruppen. Das Ergebnis einer phänetischen Untersuchung ist die Positionierung aller Arten in einem mehrdimensionalen Raum, der fachsprachlich als "morphospace" bezeichnet wird. Jeder Art entspricht genau ein Punkt dieses Raumes; der Abstand zweier Punkte spiegelt den Grad der morphologischen Unterschiede zwischen den beiden zugehörigen Arten wider. Durch Verbindung jedes Punktes mit seinem nächsten Nachbarn entsteht schließlich ein Netz, das das Ausmaß struktureller Ähnlichkeit (nicht notwendigerweise aber stammesgeschichtliche Verwandtschaft) anzeigt.
Die Untersuchungen ergaben, dass alle kambrischen Formen im "morphospace" eine nah beieinander liegende Gruppe, einen so genannten "Cluster" bilden. Die einzige aus dem Cluster heraus laufende Linie verbindet diesen mit der aus dem Karbon stammenden Gattung Priapulites, an welche die modernen Formen angebunden sind. Diese bilden jedoch keinen Cluster, sondern zwei sehr weit auseinander liegende Gruppen, die von den Arten der Familie Tubiluchidae einerseits und allen anderen modernen Formen andererseits gebildet werden. Letztere nehmen selbst ein relativ diffuses Gebiet ein, sind also ebenfalls nicht eng im "morphospace" konzentriert.
Dieses Ergebnis wird heute wie folgt gedeutet:
- Die morphologische Vielfalt kambrischer Formen ist bedeutend kleiner als die heutige. Zumindest bei den Priapswürmern kann von einem postkambrischen Zusammenbruch des Formenreichtums keine Rede sein.
- Anders als vielfach angenommen, haben sich moderne Priapswürmer in ihrem Aufbau von ihren ausgestorbenen Vorfahren deutlich abgesetzt. Dies steht im Gegensatz zu Ergebnissen bei den Gliederfüßern (Arthropoda), bei denen sich die von ausgestorbenen und modernen Formen eingenommenen "morphospace"-Gebiete weitgehend überlappen. Priapswürmer haben daher noch nach dem Kambrium wesentliche morphologische Neuerungen hervorgebracht - was zumindest für dieses Taxon die These widerlegt, dass das Kambrium eine besondere Zeit der evolutionären "Experimente" war, nach der wesentliche Fortentwicklungen des "Bauplans" durch eine straff integrierte genetische Steuerung der embryologischen Entwicklung stark eingeschränkt waren.
- Die morphologischen Veränderungen hängen vielleicht mit der Verdrängung der Priapswürmer aus ihrem ursprünglichen Lebensraum in die heutigen, ökologisch marginalen Gebiete zusammen. Diese These wird dadurch gestützt, dass die Arten der Familie Tubiluchidae, die im morphologischen Raum deutlich getrennt von den anderen modernen Formen zu liegen kommen, auch einen anderen, wesentlich lebensfreundlicheren Lebensraum besiedeln.
Interessanterweise ergab eine Einbeziehung (moderner) Formen der Hakenrüssler und Korsetttierchen in die phänetische Analyse eine wesentlich engere morphologische Beziehung zu den ausgestorbenen Priapswürmern - ob dies auf eine wesentlich konservativere evolutionäre Entwicklung bei diesen beiden Tiergruppen hinweist, ist allerdings unklar.
Systematik
Man unterscheidet insgesamt achtzehn moderne Arten, die in vier Ordnungen innerhalb einer Klasse aufgeteilt werden.
- Die Priapulomorpha haben als einzige Ordnung Schwanzanhänge und teilen sich ihrerseits in zwei Familien auf, die großen Priapulidae, deren Männchen und Weibchen sich nicht voneinander unterscheiden lassen und die mittelgroßen Tubiluchidae, die Sexualdimorphismus, also ein unterschiedliches Aussehen der Geschlechter, zeigen. Anders als die anderen Priapswürmer leben letztere in sauerstoffreichen, warmen Flachgewässern gemäßigter und tropischer Zonen mit hoher Artenvielfalt. Die Priapulomorpha sind wahrscheinlich polyphyletisch, also eine künstliche Gruppe, die beiden Familien also vermutlich keine Schwestertaxa.
- Die Halicryptomorpha sind große räuberische Tiere ohne Schwanzanhänge. Es gibt nur eine Familie, Halicryptidae mit zwei Arten in der Gattung Halicryptus; Halicryptus higginsi ist der längste Priapswurm überhaupt.
- Die Meiopriapulomorpha sind ein monotypisches Taxon, enthalten also nur eine Art, Meiopriapulus fijensis in der Familie Meiopriapulidae. Die Tiere sind kleiner als zwei Millimeter und besitzen keine Zähnchen im Schlund; sie ernähren sich vermutlich detrivor, also von organischen Reststoffen, und sind lebendgebärend mit direkter Entwicklung der Larven. Sie besitzen als einzige Art dieses Taxons ein echtes histologisches Coelom.
- Die Seticoronaria schließlich, weniger als drei Millimeter groß, sind vermutlich Filtrierer, die Nährstoffe aus dem Wasser aufnehmen. Sie besitzen eine charakteristische Hakenreihe am Rumpfende und am Vorderende eine Tentakelkrone, die vermutlich auch der Sinneswahrnehmung dient. Es existiert eine Gattung Maccabeus mit zwei Arten in einer Familie Chaetostephanidae.
Ein vorgeschlagenes Schema der stammesgeschichtlichen Verwandtschaftsverhältnisse der Ordnungen zueinander wird durch das folgende Diagramm wiedergegeben:
Priapswürmer |- -(Fossile Formen, † Ottoia unter anderem ) |- -(Moderne Formen) |- -N. N. | |--Priapulomorpha, Familie Tubiluchidae | |--Meiopriapulomorpha | |- -N. N. |- -Seticoronaria |- -N. N. |- -Halicryptomorpha |- -Priapulomorpha, Familie Priapulidae (inkl. † Priapulites)
Literatur
- R. C. Brusca, G. J. Brusca: Invertebrates. Zweite Auflage. Sinauer Associates, 2003, S. 365, ISBN 0878930973.
- E. E. Ruppert, R. S. Fox, R. D. Barnes: Invertebrate Zoology, A Functional Evolutionary Approach. Siebte Auflage. Brooks/Cole, 2004, S. 772, ISBN 0-03-025982-7.
Wissenschaftliche Literatur
- R. D. Adrianov, V. V. Malakhov: Priapilida (Priapulida): Structure, development, phylogeny, and classification. KMR Scientific Press, 1996, S. 266
- X.-P. Dong, P. C. J. Donoguhe, H. Cheng, J. B. Liu: Fossil embryos from the Middle and Late Cambrian period of Hunan, south China. In: Nature 427. 2004, S. 237
- D.-Y. Huang, J. Vannier, J.-Y. Chen: Anatomy and lifestyles of Early Cambrian priapulid worms exemplified by Corynetis and Anningvermis from the Maotianshan Shale (SW China). In: Lethaia 37. 2004, S. 21
- V. Storch: Priapulida. In: F. W. Harrison, E. E. Ruppert (Hrsg.): Microscopic Anatomy of Invertebrates. Wiley-Liss, 1991, S. 333
- M. A. Wills: Cambrian and recent disparity: The picture from priapulids. In: Paleobiology 24. 1998, S. 177
Weblinks
Wikimedia Foundation.