- Raubdinosaurier
-
Theropoda 
Zeitraum Obere Trias bis Obere Kreide
(Vögel bis Jetztzeit)227 bis 65,5 (bzw. bis heute) Mio. Jahre Fossilfundorte - Weltweit
Systematik Diapsida Archosauria Ornithodira Dinosaurier (Dinosauria) Echsenbeckendinosaurier (Saurischia) Wissenschaftlicher Name Theropoda Marsh, 1881 Untertaxa Die Theropoda sind eine systematische Gruppe (Taxon) der Echsenbeckendinosaurier (Saurischia). Sie bewegten sich zweibeinig fort und waren zum größten Teil Fleischfresser. Umgekehrt gehörten nahezu alle fleischfressenden Dinosaurier, darunter die nach heutiger Kenntnis größten landbewohnenden Fleischfresser aller Zeiten mit Längen von rund 15 Metern, dieser Gruppe an. Viele Theropoda waren jedoch verglichen mit anderen Dinosauriern klein und maßen oft nur 2 bis 6 Meter.
Aus der aktuellen Sicht der Systematik (das heißt, nach Definition der Kladistik) können nur solche Gruppen Gültigkeit haben, die alle Nachfahren einer gemeinsamen Vorfahrenart einschließen. Da die Vögel im Jura nachweislich aus noch nicht flugfähigen Theropoden hervorgegangen sind, welche viele gemeinsame Merkmale mit Vögeln wie beispielsweise Federn aufwiesen, gilt die Gruppe der Vögel (Aves) als den Theropoden zugehörig.
Demnach überlebten die Theropoda (und somit die Dinosaurier) das Massenaussterben am Ende der Kreidezeit und kommen, vertreten durch die modernen Vögel, noch heute vor. Der nachfolgende Artikel beschränkt sich jedoch weitgehend auf fossile Vertreter des Mesozoikums, die nicht den Vögeln angehörten („Nicht-Vogel-Theropoden“ im Sinne des englischen Neologismus non-avian theropods).
Im Zuge der Popularisierung der Dinosaurier seit Mitte des 19. Jahrhunderts entstanden zahlreiche Lebendrekonstruktionen gut bekannter Theropodengattungen wie Megalosaurus, Tyrannosaurus, Allosaurus, Deinonychus und Velociraptor, die das öffentliche Bild der Theropoda seither prägen.
Inhaltsverzeichnis
Merkmale
Allgemeines
 Giganotosaurus war mit geschätzten 14 Metern Länge einer der größten Theropoden
Giganotosaurus war mit geschätzten 14 Metern Länge einer der größten Theropoden Bambiraptor hingegen wurde nur knapp 1 Meter lang
Bambiraptor hingegen wurde nur knapp 1 Meter langDie Theropoda haben in ihrem allgemeinen Körperbau den ursprünglichen Körperbau der Dinosaurier beziehungsweise den ihrer Vorfahren in vielen Aspekten beibehalten. Verglichen mit anderen Dinosauriergruppen wie den Sauropodomorpha oder den Ornithischia zeigten sie wenig Spezialisierungen. Theropoden waren durchgehend biped, ihre Fortbewegung fand also ausschließlich mit den Hinterbeinen statt, die darum bei allen Theropoden länger als die Vordergliedmaßen waren. Die Vorderbeine waren zur Fortbewegung unbrauchbar, was der ursprünglichen Körperhaltung der Dinosaurier entspricht.
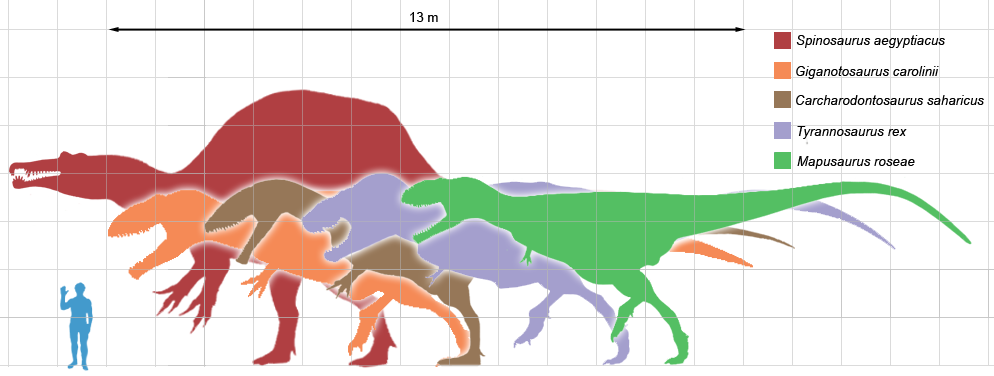
Bei den Theropoda herrschte hinsichtlich der Körpergröße die größte Diversität innerhalb der Dinosaurier. Viele waren verhältnismäßig klein, manche Vertreter wie Microraptor und Parvicursor maßen weniger als einen Meter und wogen nur wenige Kilogramm. Die größten Theropoden hingegen konnten Längen von rund 15 Metern erreichen, dazu zählen Tyrannosaurus, Carcharodontosaurus, Giganotosaurus und Spinosaurus. Größenangaben sind aber oft hypothetisch, da von vielen Vertretern nur sehr unvollständige Skelette gefunden wurden. Gewichtsschätzungen sind bei ausgestorbenen Tieren allgemein schwierig; das Gewicht der größten Theropoden wird auf 5 bis 8 Tonnen geschätzt.
Schädel
 Schädel von Dromaeosaurus mit gut erkennbaren Schädelfenstern
Schädel von Dromaeosaurus mit gut erkennbaren SchädelfensternDie Theropoda hatten meist einen verhältnismäßig großen Schädel. Der Bau des Schädels ist verglichen mit anderen Dinosauriern eher unspezialisiert und zeigt wenig Veränderungen gegenüber dem urtümlichen Schädel der Archosauria („triapsider Schädel“). Wie bei allen Archosauria ist neben den für die Diapsiden typischen zwei Schädelfenstern hinter der Augenhöhle eine zusätzliche Schädelöffnung vor der Augenhöhle (Präorbitalfenster oder Antorbitalfenster) vorhanden, wodurch sich ein leichter, spangenartiger Schädelbau ergibt. Eine evolutive Neuerung (Synapomorphie) der Theropoda ist das bis auf die Schädeloberseite reichende Tränenbein (Os lacrimale) sowie eine zusätzliche antorbitale Öffnung, das Maxiliarfenster. Die Augen der Theropoden waren vergleichsweise groß, insbesondere bei den Coelurosauria, und zum Teil nach vorn orientiert – wie bei vielen Fleischfressern. Das so verbesserte räumliche Sehen ermöglichte eine präzisere Abschätzung der Entfernung zur möglichen Beute. Auch die Mittelohrhöhle war oft stark vergrößert. Daraus lässt sich schließen, dass die Theropoda einen vergleichsweise guten Gesichts- und Gehörsinn besessen haben. Eine weitere Synapomorphie der Theropoda ist der Bau des Gelenkknorrens am Hinterhauptsbein (Condylus occipitalis), der zusammen mit dem Atlas das obere Kopfgelenk bildet. Dieser ist groß und rund und sorgt so für eine erhöhte Mobilität des Kopfes – vermutlich in Kombination mit einer kräftigen Nackenmuskulatur. Der Theropodenschädel zeigt außerdem eine gewisse Beweglichkeit der einzelnen Elemente zueinander (Schädelkinese), die das Verschlingen größerer Beutestücke erleichterte, da die Theropoden ihre Nahrung nicht durch Kauen zerkleinern konnten.
Einige Vertreter der Theropoda wiesen an der Oberseite des Kopfes Schädelkämme oder hornartige Auswüchse auf. Diese Auswüchse unterschieden sich deutlich in ihrem Bau voneinander, manchmal waren sie sehr fragil – wie die Kämme von Dilophosaurus oder Oviraptor – manchmal sehr robust gebaut – wie die Hörner von Ceratosaurus oder Carnotaurus. Diese Schädelstrukturen spielten vermutlich eine Rolle bei der Interaktion mit Artgenossen.
Zähne
 Schädel von Allosaurus mit nach hinten gebogenen Zähnen
Schädel von Allosaurus mit nach hinten gebogenen ZähnenDie Zähne der Theropoden waren seitlich abgeflacht und leicht rückwärts gekrümmt. Sie waren zudem an der Vorderkante mit einer Serration versehen, das heißt, mit kleinen Zacken, die etwa im rechten Winkel zur Schneidekante standen, und darum sehr scharf. Die Form der Serration entlang der Schneidekante unterschied sich bei einzelnen Gruppen: So hatten viele kleinere Vertreter kleine, spitze Zacken, die für reißende Tätigkeiten geeignet waren, während etwa bei den Tyrannosauridae die Zähnelung rundlicher und stumpfer war. Der Bau der Zähne ist jedoch urtümlich, das gleiche Muster findet sich beispielsweise auch bei Vertretern der Crurotarsi, basalen Krokodilvorfahren. Auch die Zähne der nahe verwandten Prosauropoden zeigen eine Serration der Zähne, doch stehen die Kerben zu den Schneidekanten in einem Winkel von etwa 45° nach oben oder verlaufen in verschiedene Richtungen.
Aus dem Bau der Zähne ist ersichtlich, dass die meisten Theropoden Fleischfresser waren. Sie setzten ihre Zähne zum Packen der Beute, zum Töten und auch zum Zerreißen der Fleischstücke ein; die stumpferen Zähne mancher Vertreter könnten auch zum Zermalmen von Knochen gedient haben. Die pflanzenfressenden Therizinosauria hingegen hatten kleine, spatelförmige Zähne.
Mehrmals in der Entwicklungsgeschichte der Theropoda ist es zum Verlust der Zähne gekommen. Das betrifft zum einen die Ornithomimosauria (deren ursprünglichste Vertreter allerdings noch die vordersten Zähne des Unterkiefers aufwiesen), und zum anderen die Oviraptorosauria. Was genau diese zahnlosen Theropoden fraßen, ist umstritten, siehe dazu den Abschnitt Ernährung. Auch die modernen Vögel sind – im Gegensatz zu vielen ihrer mesozoischen Vorfahren – zahnlos.
Rumpfskelett
Die Hintergliedmaßen waren, wie bei allen Dinosauriern, senkrecht unter dem Körper angebracht, der Körper war direkt über dem Becken ausbalanciert. Die Wirbelsäule wurde nahezu waagrecht gehalten; um dennoch eine gute Sicht nach vorne zu ermöglichen, war der Hals vogelartig S-förmig gebogen – ältere Rekonstruktion stellen die Theropoda mit abwärts geneigter Wirbelsäule und am Boden schleifendem Schwanz dar, das dürfte nach jüngeren Erkenntnissen falsch sein. Eine Synapomorphie (ein gemeinsam abgeleitetes Merkmal) der Theropoden war, dass fünf oder mehr Sakralwirbel vorhanden waren. Der Schwanz war meist lang und wies eine hohe Anzahl von Schwanzwirbeln auf. Bei den Tetanurae („Steifschwänzen“), der artenreicheren der beiden Untergruppen der Theropoden, war der Schwanz versteift und wurde mehr oder weniger waagerecht über den Boden gehalten.
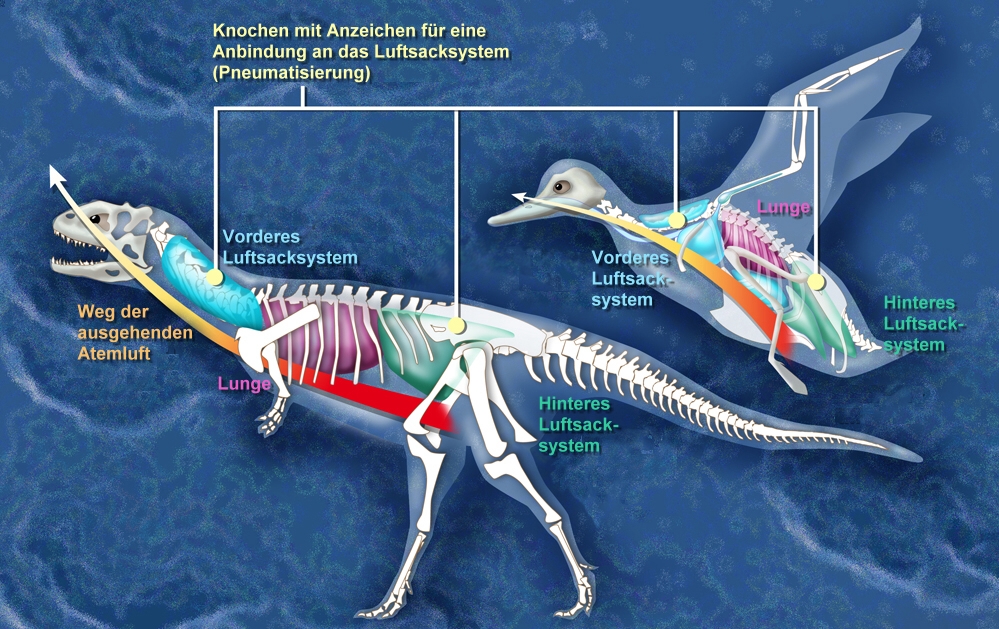
 Vergleich des Luftsacksystems von Dinosauriern (Theropoden) und Vögeln
Vergleich des Luftsacksystems von Dinosauriern (Theropoden) und VögelnDiese Versteifung wurde einerseits dadurch erreicht, dass die Schwanzwirbel durch lange, nach vorne und hinten reichende Knochenstäbe (Zygapophysen) und andererseits durch Chevron-Knochen (V-förmige Fortsätze an der Unterseite der Schwanzwirbel) verbunden waren. Bei den modernen Vögeln sind die letzten Schwanzwirbel zum Pygostyl verwachsen, eine ähnliche Struktur findet sich auch bei manchen Nicht-Vogel-Theropoden wie Nomingia und Beipiaosaurus. Dabei dürfte es sich allerdings um konvergente Entwicklungen handeln.
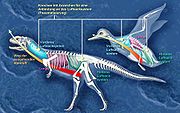
Das Skelett der Theropoda ist dadurch charakterisiert, dass die langen Gliedmaßenknochen dünnwandig und hohl sind, auch die Wirbel sind ausgehöhlt. Bereits bei basalen Vertretern wie Majungasaurus waren die Wirbel pneumatisiert, das heißt teils gefüllt mit Ausstülpungen (Divertikeln) eines Luftsacksystems. Auch bei Mirischia wurden vergleichbare Funde gemacht. Das ist ein Anzeichen, dass zumindest die Theropoda (möglicherweise aber auch andere Dinosaurier) eine Durchströmungsatmung wie die heutigen Vögel aufwiesen (siehe auch Evolutionärer Ursprung des Luftsacksystems).
Gliedmaßen
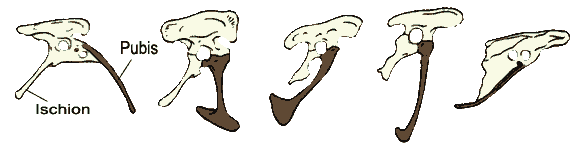 Relative Lage des Pubis' im Beckengürtel vom primitiven Zustand bis zu den Vögeln
Relative Lage des Pubis' im Beckengürtel vom primitiven Zustand bis zu den Vögeln
1 Coelophysis, 2 Allosaurus, 3 Velociraptor, 4 Archaeopteryx, 5 Columba (Feldtauben)Der Schulterbereich der Theropoden war charakterisiert durch ein riemenartiges Schulterblatt. Zusätzlich war meist ein bumerangförmiges Gabelbein (Furcula) – das einst als reines Vogelmerkmal gesehen wurde – vorhanden. Ein Gabelbein ist schon bei basalen Vertretern wie den Coelophysoidea bekannt.
Im Bau des Beckens ragte wie bei allen Echsenbeckensauriern (Saurischia) das Schambein (Os pubis) ursprünglich nach vorne. Im Lauf der Entwicklung der Theropoda ist es zu einer Rotation gekommen, bei den Deinonychosauria weist es nach unten und bei den Vögeln schließlich nach hinten. Damit ähnelt das Becken der heutigen Vögel verwirrenderweise dem der Vogelbeckensaurier, der zweiten Untergruppe der Dinosaurier. Trotzdem haben sich Vögel aus den Echsenbecken- und nicht aus den Vogelbeckensauriern entwickelt, die Ähnlichkeit ist nur oberflächlich.
 Skelett von Deinonychus: Deutlich sind die langen Arme mit den großen Händen und die spezialisierte zweite Zehe mit der großen Kralle zu erkennen.
Skelett von Deinonychus: Deutlich sind die langen Arme mit den großen Händen und die spezialisierte zweite Zehe mit der großen Kralle zu erkennen.Synapomorphien der Theropoda im Bau der Vordergliedmaßen waren zum einen die vergrößerte Hand, die bis zu 50 % der Länge des gesamten Arms ausmacht, und zum anderen eine Tendenz zur Rückbildung des vierten und fünften Fingers bis zu ihrem völligen Fehlen. Die Hand bestand also typischerweise aus den Strahlen (Finger, Digiti) I, II und III (vom Daumen her gezählt), wobei der erste Strahl teilbeweglich, also halb-opponierbar war. Bei den Tyrannosauridae war auch der dritte Finger rückgebildet und Alvarezsauridae wie Mononykus („eine Klaue“) besaßen nur noch einen gut entwickelten Fingerstrahl. Die verbleibenden Finger waren stark verlängert und dank großer Vertiefungen an der Oberseite des vorderen Endes der Mittelhandknochen äußerst beweglich. Viele Theropoda, oftmals die kleineren Vertreter, waren durch lange Arme mit großen Händen und schlanken Fingern gekennzeichnet. Bei diesen Tieren spielte die Greiffunktion der Hand – unter anderem zum Beutefang – eine große Rolle. (Bei den Vögeln sind die Arme in Anpassung an das Fliegen besonders stark abgewandelt, so haben sie einen Carpometacarpus aus verschmolzenen Handwurzel- und Mittelhandknochen.)
Im Gegensatz dazu zeigten einige große Theropoden wie die Abelisauroidea und die Tyrannosauridae stark verkleinerte Vordergliedmaßen. Bei diesen Tieren waren die Arme so kurz, dass sie nicht einmal den Mund erreichen konnten – sie haben bei der Jagd sicher keine Bedeutung gehabt. Die Frage nach der Funktion dieser verkleinerten Arme ist nicht restlos geklärt. Nach einer Theorie ist dieses Merkmal biomechanisch bedingt: So mussten, um die Balance aufgrund des stark vergrößerten und damit schwerer gewordenen Kopfes aufrecht zu erhalten, die Arme verkleinert werden. Auffällig ist, dass diese kurzen Arme sehr kräftig und muskulös waren. Möglicherweise dienten sie dazu, um sich aus der vermuteten Schlafposition (auf dem Bauch liegend) wieder aufrichten zu können. Andere Theorien sehen ihre Funktion hingegen im Festhalten des Partners bei der Paarung.
 Typischer dreizehiger Theropodenfuß
Typischer dreizehiger TheropodenfußDie Hintergliedmaßen waren stets länger als die Vordergliedmaßen. Meist war der Unterschenkel länger als der Oberschenkel (insbesondere bei den Troodontidae und den Ornithomimosauria), was vermuten lässt, dass diese Tiere hohe Geschwindigkeiten erzielen konnten. Der Fuß war durch einen kompakten, schmalen und üblicherweise verlängerten Mittelfuß (Metatarsus) gekennzeichnet – mit dem die Ornithodira (Dinosaurier und Flugsaurier) kennzeichnenden mesotarsalen Sprunggelenk. Die Zehen II, III und IV waren symmetrisch nach vorne gerichtet und besaßen eine charakteristische Anzahl von Zehengliedern (Phalangenformel 3, 4, 5). Diese Zehen waren die Laufzehen, denn die innere, erste Zehe war nur kurz und berührte den Boden nicht und die äußere, fünfte Zehe war auf ein kleines Knöchelchen reduziert. Der typische Theropodenfuß war demnach funktional tridactyl. Bei den modernen Vögeln weist die erste Zehe meist nach hinten (Anisodactylie), in dieser Gruppe haben sich aber auch andere Zehenanordnungen entwickelt.
Die Zehen endeten in scharfen Klauen oder Krallen aus Horn, die auf spitz zulaufenden, gebogenen Knochenzapfen saßen. Eine Besonderheit entwickelte sich innerhalb der Deinonychosauria. Hier war die zweite Zehe mit einer besonders großen und scharfen Kralle ausgestattet und wies zusätzlich ein spezielles Gelenk auf. Dieses ermöglichte das Hochheben der Zehe bei der Fortbewegung (funktionale Didactylie), um Abnutzung zu vermeiden, und eine hohe Beweglichkeit bei der Jagd. Den ebenfalls bipeden und dreizehigen Ornithopoda aus der Gruppe der Vogelbeckendinosaurier fehlen die langen Krallen, was für die Zuordnung fossiler Fußabdrücke bedeutsam ist.
Integument und Thermoregulation
 Fossil von Sinosauropteryx mit Spuren von Federn
Fossil von Sinosauropteryx mit Spuren von FedernDie wohl drastischste Änderung in der menschlichen Vorstellung über das Aussehen der Theropoda betraf das Integument (die Haut). Bei Sinosauropteryx wurden 1996 erstmals bei einem Nicht-Vogel-Theropoden Anzeichen eines Federkleides entdeckt, in den nachfolgenden Jahren wurden weitere gefiederte Dinosaurier beschrieben, sodass heute bei rund 15 Vertretern Anzeichen eines Gefieders bekannt sind. Dabei sind unterschiedliche Federarten – von einfachen, haarähnlichen Strukturen bis zu Konturfedern – bekannt. (Federn fossilieren als weiche Struktur nur selten, diese Funde sind bislang auf wenige Fundstätten beschränkt.)
Die meisten gefiederten Theropoden lassen in ihrem Körperbau keinerlei Anzeichen für eine Anpassung an den Flug erkennen, sodass das Gefieder ursprünglich wahrscheinlich der Thermoregulation diente. Denkbar ist aber auch, dass das Federkleid später auch andere Funktionen übernahm, beispielsweise Sichtschutz- oder Signalwirkung bei der Kommunikation mit Artgenossen.
Gemäß den Theorien von Richard Prum und Alan Brush haben die Federn fünf Entwicklungsstufen durchschritten[1] (siehe auch Evolution der Feder):
- Im ersten Schritt bildeten sich einfache, zylinderförmige, haarähnliche Hohlstäbe.
- In einem zweiten Schritt entstanden Büschel von verlängerten Fäden, die auf einer gemeinsamen Federspule (Calamus) saßen. Diese Büschelfedern ähnelten den Daunen der heutigen Vögel, wiesen aber noch keine Nebenäste auf.
- Beim dritten Schritt kam es zur Entwicklung zweier Federtypen. Zum einen sind dies die frühesten Konturfedern mit auf dem Federschaft aufsitzenden Federästen – allerdings noch ohne die Verzahnung. Zum anderen entstanden auch die mit Nebenstrahlen versehenen Büschelfedern, die sich heute noch in Form der Daunen finden.
- Der vierte Schritt bestand in der Entwicklung einer Deckfeder mit ineinander verzahnten Bogen- und Hakenstrahlen. Diese ähnelte der Konturfeder der heutigen Vögel, war aber im Gegensatz zu dieser noch symmetrisch.
- In einem fünften Schritt entwickelte sich schließlich die bei den heutigen Vögeln vorkommende asymmetrische Feder mit einer Außen- und einer Innenfahne.
Weitere Untersuchungen haben zu vermuteten Übereinstimmungen zwischen der Entwicklung der Feder und der allgemeinen Entwicklung der Theropoden (siehe Kladogramm im Abschnitt Systematik) geführt. Die Coelurosauria dürften dementsprechend mit einem Federkleid ausgestattet gewesen sein, das deckt sich damit, dass bei Sinosauropteryx (der als Compsognathidae und damit als basaler Coelurosaurier klassifiziert wird) einfache Federn vom Typ 1 oder 2 vorhanden waren. Auch bei Dilong, einem urtümlichen Vertreter der Tyrannosauroidea, fand man Anzeichen dieser einfachen Federn. Bei dem Therizinosaurier Beipiaosaurus wurden Federn des Typs 2 oder 3, bei dem Oviraptorosaurier Caudipteryx jene der Stufe 4 entdeckt. Bei den Deinonychosauria wurden bei Microraptor bereits Anzeichen der asymmetrischen Federn des Typs 5 entdeckt, die auch beim frühesten Vogel Archaeopteryx entdeckt wurden.Die Frage, ob die Dinosaurier generell gleichwarm („warmblütig“) oder wechselwarm („kaltblütig“) waren, ist noch nicht endgültig geklärt (siehe dazu Warmblütigkeit der Dinosaurier). Seit der Entdeckung der gefiederten Dinosaurier geht man davon aus, dass zumindest bei den gefiederten Coelurosauria eine Art der inneren Temperaturregelung vorhanden war und dass bei den Theropoden mit höher entwickeltem Gefieder die Thermoregulation annähernd ebenso effektiv wie die der Vögel war. Andere Theropoden könnten nach Meinung mancher Paläontologen eine andere Methode zur Thermoregulation entwickelt haben: die verlängerten Wirbelfortsätze mancher Theropoden wie Spinosaurus und Acrocanthosaurus könnten ein Rückensegel gestützt haben, das diesem Zweck diente. Diese Sichtweisen sind jedoch umstritten.
Nach Meinung einiger Forscher könnten aber die größten Coelurosauria wie Tyrannosaurus keine Federn gehabt haben. [2] Bei großen Tieren ist das Verhältnis der Oberfläche zum Volumen kleiner als bei kleineren. Da über die Hautoberfläche die Wärme entweicht, droht größeren Tieren eher die Gefahr der Überhitzung, da ein Gefieder die Wärmeabgabe behindert. (Aus den gleichen Gründen sind auch die größten Säugetiere wie Elefanten und Nashörner nahezu haarlos.) In dieses Schema passt auch, dass es innerhalb der (gefiederten) Coelurosauria mit Ausnahme der Tyrannosauridae keine Riesenformen mehr gab.
Wie die Haut der Nicht-Coelurosauria innerhalb der Theropoden aussah, ist nicht genau bekannt. Bei einem Vertreter, Carnotaurus aus der Gruppe der Abelisauroidea, wurden Anzeichen einer schuppigen Haut entdeckt.
Paläobiologie
Verbreitung und Lebensraum
Die Theropoden waren weltweit verbreitet, Fossilfunde sind aus allen Kontinenten bekannt – mit Cryolophosaurus auch aus der Antarktis. Die frühe weltweite Verbreitung hängt damit zusammen, dass zur Zeit der Entwicklung der Theropoden alle Erdteile noch im Großkontinent Pangaea verbunden waren. Seit dem Auseinanderbrechen dieses Kontinents entwickelten sich auf den einzelnen Landmassen teilweise unterschiedliche Theropodenfaunen. So sind die Abelisauroidea fast ausschließlich aus dem ehemaligen Südkontinent Gondwana bekannt, andere Gruppen wie die Tyrannosauroidea hingegen nur aus dem Nordkontinent Laurasia. Da Fossilfunde auf der Südhalbkugel generell seltener sind, lassen sich für viele Theropodentaxa keine genauen Angaben zum Verbreitungsgebiet machen.
Auch bei den Ablagerungsorten der Theropoden herrscht eine enorme Bandbreite. Neben Flussbetten, Überschwemmungsgebieten und der Umgebung von Seen wurden Funde auch an Orten gemacht, die auf einen trockenen, wüstenhaften Lebensraum schließen lassen. Eine Habitatspräferenz der Theropoden als Gesamtgruppe ist – sofern überhaupt vorhanden – nicht bekannt.
Sozialverhalten
Spekulationen über das Sozialverhalten der Theropoda sind, wie bei allen nur durch Fossilfunde bekannten Tieren, schwierig. Selbst Funde von Überresten mehrerer Tiere an einem Ort müssen nicht auf ein Gruppenleben hindeuten, sondern können auch ablagerungstechnisch bedingt sein. Die meisten Funde stammen von Einzeltieren. Es gibt jedoch zwei Vertreter, bei denen Massenansammlungen von hunderten Tieren entdeckt wurden: Coelophysis in Ghost Ranch (New Mexico) und Allosaurus im Cleveland-Lloyd-Steinbruch (Utah). Die Interpretation dieser Massenfunde ist umstritten, neben einer Deutung als Hinweis auf Rudeljagden werden auch die Möglichkeiten erwogen, dass es sich um versiegende Wasserquellen handelte, an denen sich viele Tiere versammelt haben und schließlich verdurstet sind; oder dass es sich um eine Falle in irgendeiner Form handelte, die die Tiere anlockte und aus der sie sich nicht mehr befreien konnten. Auch Funde von mehreren Deinonychus, unter anderem in Zusammenhang mit dem möglichen Beutetier Tenontosaurus, wurden als Hinweis auf Rudeljagden betrachtet. [3]
 Rekonstruktion von Dilophosaurus mit deutlich sichtbarem Schädelkamm
Rekonstruktion von Dilophosaurus mit deutlich sichtbarem SchädelkammDie oben erwähnten Kämme und Auswüchse an der Oberseite des Schädels spielten vermutlich eine Rolle bei der Interaktion mit Artgenossen. Während die fragilen Kämme vieler Tiere wohl keinem anderem Zweck als der Zurschaustellung dienten, ist es denkbar, dass die robusteren Hörner für direkte kämpferische Auseinandersetzungen verwendet wurden. Ob es dabei um das Paarungsvorrecht, die Etablierung einer Gruppenhierarchie oder andere Zwecke ging, muss Spekulation bleiben.
Bei einigen Theropoda sind Anzeichen von Geschlechtsdimorphismus erkennbar. So gibt es etwa bei manchen Coelophysoidea zwei Formen, eine mit langem Kopf und Hals und kräftigeren Gliedmaßen und eine andere Form mit kurzem Kopf und schlankeren Gliedmaßen. Dabei könnte es sich um einen leicht unterschiedlichen Körperbau bei Männchen und Weibchen handeln. Auch bei Tyrannosaurus könnte es ähnliche Unterschiede zwischen den Geschlechtern gegeben haben. Die robustere Form wird aufgrund des breiteren Beckens als Weibchen identifiziert. [4]
Fortbewegung
 Fossilierter Fußabdruck (Ichnofossil) eines großen Theropoden
Fossilierter Fußabdruck (Ichnofossil) eines großen TheropodenWie eingangs erwähnt, bewegten sich die Theropoda ausschließlich mit den Hintergliedmaßen fort. Dabei berührten nur die Zehen den Boden, sie waren also Zehengänger (digitigrad). Aus Ichnofossilien (fossilen Fußspuren) ergibt sich, dass die Hinterbeine unter dem Körper gehalten wurden. Die Schrittbreite war sehr schmal, oft wurde ein Fuß vor den anderen gesetzt. Die Beine waren nur in einer Ebene (vorwärts-rückwärts) bewegbar. Aufgrund des Baus der Gelenke war es den Theropoda nicht möglich, die Gliedmaßen nach außen zu drehen, wie es etwa Säugetiere können.
Es gibt verschiedene Versuche, anhand des Skelettbaus und Ichnofossilien die Geschwindigkeiten von Dinosauriern zu berechnen, etwa von Robert McNeill Alexander [5] und R. A. Thulborn [6]. Nach diesen Berechnungen könnten die kleineren Theropoden bis zu 40 km/h erreicht haben, die Ornithomimosauria sogar zu bis 60 km/h. Für größere Theropoda werden geringere Geschwindigkeiten vermutet, doch sind diese Berechnungen umstritten.
Bei manchen Deinonychosauria wie Microraptor ist es denkbar, dass sie auch auf Bäume klettern und Gleitflüge unternehmen konnten.
Gehirngröße und Intelligenz
Die Theropoda sind die Dinosaurier mit den vergleichsweise größten Gehirnen. Zur Berechnung der Gehirngröße wurde von J. A. Hopson der Enzephalisationsquotient (EQ) entwickelt [7]. Dies ist eine Berechnungsmethode, die mit der Größe der Schädelhöhle und allometrischen Faktoren – größere Lebewesen haben vergleichsweise kleinere Gehirne als kleinere – arbeitet. Als Vergleichswert dienen dazu die Krokodile, deren EQ = 1 festgelegt wird. Alle Theropoden haben einen EQ > 1 (Zum Vergleich: Unter den Dinosauriern weisen sonst nur manche Vertreter der Ornithopoda einen EQ > 1 auf, alle anderen Dinosaurier liegen darunter. Den kleinsten EQ haben Sauropodomorpha, gefolgt von Ankylosauria und Stegosauria). Innerhalb der Theropoda haben die Dromaeosauridae den höchsten EQ mit 5,8, ein Wert, der mit Straußen vergleichbar ist. [8]
Ernährung
Als einzige Gruppe der Dinosaurier (mit der möglichen Ausnahme einiger urtümlicher Vertreter, siehe unten) ernährten sich die Theropoda mehrheitlich von Fleisch. Aufgrund der großen Bandbreite in Körperbau und Größe haben sie zweifelsohne unterschiedliche Ernährungsstrategien entwickelt. Vereinzelte Fossilfunde wie Bissspuren oder Koprolithen lassen manchmal Rückschlüsse auf die Ernährungsweise zu, in vielen Fällen bleibt die Art des Beuteerwerbs und die Frage nach möglichen Beutetieren jedoch spekulativ. D. Fastovsky und J. B. Smith haben fünf Morphotypen fleischfressender Theropoden herausgearbeitet. [9] Dabei handelt sich um keine verwandtschaftliche Klassifikation, weswegen die Typen auch nicht mit Namen versehen werden.
 Die größten Theropoden im Vergleich
Die größten Theropoden im Vergleich- Der Morphotyp 1 umfasst die extrem großen Vertreter mit über 10 Metern Länge. Bei diesem Typ herrscht eine Tendenz zu einem großen Schädel, kurzen Vordergliedmaßen und verringerter Fingeranzahl, bekannte Beispiele aus dieser Gruppe sind Tyrannosaurus, Tarbosaurus, Carcharodontosaurus und Giganotosaurus. Dieser Typ umfasst die nach heutigem Kenntnisstand größten landbewohnenden Fleischfresser aller Zeiten, es gibt keine modernen Analogien zu diesen Tieren. Ihre Ernährungsweise ist vielfach umstritten, bei manchen Vertretern, insbesondere den Tyrannosauridae, wird diskutiert, ob sie Aasfresser waren oder ihre Beute selbst erlegten. Anhaltspunkte, die für eine aasfressende Ernährung sprechen könnten, sind unter anderem die breiten Zähne, die zum Zermalmen von Knochen geeignet waren, und der Bau der Gliedmaßen. Im Gegensatz zu den meisten anderen Theropoda waren Ober- und Unterschenkel annähernd gleich lang, was eher für eine langsame Fortbewegung spricht.
- Der Morphotyp 2 umfasst die mittelgroßen, rund 3 bis 8 Meter langen Theropoden. Sie hatten ebenfalls einen eher großer Schädel, verglichen mit dem Typ 1 waren die Hintergliedmaßen schlanker und die Unterschenkel deutlich länger als die Oberschenkel. Auch waren die Hände oft gut entwickelt. Als Beispiele für diesen Typ werden Allosaurus, Albertosaurus, Dilophosaurus, Ceratosaurus und Carnotaurus genannt. Die Vertreter dieses Typs waren ziemlich sicher aktive Jäger, Mutmaßungen gehen in Richtung großer Beutetiere, zum Beispiel Sauropoda.
 Der berühmte Fund der „kämpfenden Dinosaurier“: Ein Protoceratops (links) wird von einem Velociraptor angegriffen, beide Tiere starben möglicherweise durch einen Sandsturm
Der berühmte Fund der „kämpfenden Dinosaurier“: Ein Protoceratops (links) wird von einem Velociraptor angegriffen, beide Tiere starben möglicherweise durch einen Sandsturm- Der Morphotyp 3 umfasst eher kleine Theropoden (mit 2 bis 6 Metern Länge). Der Schädel war groß, die Augen nach vorn gerichtet. Sie hatten lange Arme und große Hände mit drei Greiffingern, der Körperbau war schlank und der Schwanz auffallend versteift. An der spezialisierten zweiten Zehe trugen sie eine scharfe, bewegliche Kralle. Beispiele sind Deinonychus, Troodon, Velociraptor oder Utahraptor. Laut John H. Ostrom handelt es sich dabei um „ultimate killing machines“ [10]. Bei diesen Tieren dürfte es sich um schnelle Läufer und agile, geschickte Jäger gehandelt haben, möglicherweise sind sie auch im Rudel auf Beutefang gegangen. Für diesen Typ gibt es konkrete Hinweise auf die Beutetiere, darunter die berühmten „kämpfenden Dinosaurier“ (Ein Protoceratops wird von einem Velociraptor angegriffen, beide Tiere starben im Kampf, möglicherweise durch einen Sandsturm.)
- Der Morphotyp 4 umfasst ebenfalls eher kleine, 2 bis 6 Meter lange Tiere. Sie hatten wie der Typ 3 einen schlanken Körperbau und lange Arme mit großen Händen, unterschieden sich aber durch einen kleineren Schädel und vergleichsweise kurze Hintergliedmaßen. Vertreter dieses Typs sind unter anderem Coelophysis, Compsognathus oder Syntarsus. Die Tiere vom Typ 4 waren ebenfalls aktive Jäger, die sich vermutlich von Kleintieren wie Echsen, Therapsiden (inkl. Säugetieren) oder Insekten ernährt haben. Konkrete Hinweise liefern die Überreste einer Echse, die in der Bauchhöhle eines Compsognathus gefunden wurde, sowie die Fossilien von Kleintieren im Bauch von Coelophysis, die lange Zeit fälschlicherweise für dessen Jungtiere gehalten wurden.
 Rekonstruktion von Chirostenotes, einem Vertreter der Oviraptorosauria
Rekonstruktion von Chirostenotes, einem Vertreter der Oviraptorosauria- Die Tiere des Morphotyps 5 sind ebenfalls eher klein (2 bis 6 Meter). Ihr auffälligstes Merkmal ist der zahnlose, leicht gebaute Schädel. Ihr Körper war schlank gebaut, die Arme waren lang und die großen Hände trugen drei Greiffinger. In diese Gruppe gehören die Oviraptorosauria. Der Körperbau deutet eindeutig auf aktive Jäger hin (sie ähneln bis auf die Zahnlosigkeit und die fehlende Zehenkralle dem Typ 3), aber wovon sich diese Tiere ernährt haben, ist ungewiss. Für die ursprüngliche Hypothese, sie hätten sich von Eiern ernährt, ist der Schnabel wenig geeignet. Andere Theorien interpretieren den Schnabel als Werkzeug zum Knacken von Muscheln, dies ist aber umstritten, da bei vielen Fossilfundorten kein Hinweis auf Wasser gefunden wurde. Letztlich bleibt die Ernährung dieser Tiere unklar, Vermutungen gehen in Richtung Eidechsen, Säuger und andere Kleintiere.
Einen Sonderfall innerhalb der fleischfressenden Theropoda stellen die Spinosauridae dar. Ihre Zähne sind kegelförmig, wobei die vordersten Zähne stark vergrößert sind. Aufgrund von Ähnlichkeiten mit Krokodilen wird angenommen, dass diese Tiere teils große Fische fraßen. Tatsächlich wurden bei Baryonyx in der Bauchhöhle Fischschuppen und Reste eines jungen Iguanodon gefunden, was auf eine gemischte Ernährung aus Fischen und Landtieren schließen lässt.
Zwar war die große Mehrzahl der Theropoden Fleischfresser, es haben sich jedoch auch zwei Gruppen entwickelt, bei denen im Gegensatz zu den übrigen eine pflanzliche (herbivore) Ernährung vermutet wird:
 Skelettrekonstruktion von Gallimimus, einem Vertreter der Ornithomimosauria
Skelettrekonstruktion von Gallimimus, einem Vertreter der Ornithomimosauria- Die Ornithomimosauria waren bis auf einige basale Vertreter zahnlos, ihr Maul endete in einem vogelähnlichen Schnabel. Ihr Körperbau war schlank und auf hohe Geschwindigkeiten ausgerichtet, die Vordergliedmaßen schwächlich, aber möglicherweise zum Herbeiziehen von Ästen geeignet. Bei ihnen wurden Gastrolithen (Magensteine) gefunden. Vermutungen über die Ernährung dieser Tiere gehen in Richtung Pflanzenfresser oder Allesfresser, ähnlich den heutigen Straußen, mit denen sie gerne verglichen werden. Manche Forscher haben allerdings auch Hinweise auf eine kammförmige Struktur im Schnabel entdeckt, was ein Anzeichen für eine filtrierende Nahrungsaufnahme sein könnte. Das würde damit übereinstimmen, dass die Mehrzahl der Ornithomimosauria nahe bei Süßwasserhabitaten gefunden wurden. Allerdings sind diese Funde umstritten.
- Die zweite pflanzenfressende Theropodengruppe waren die Therizinosauria. Sie wiesen kleine, spatelförmige Zähne auf, die Hände waren auffallend groß und trugen lange Krallen. Das Becken war nach kaudal (hinten) gedreht, wodurch der Rumpf vergrößert wurde, was mehr Platz für einen langen Verdauungstrakt schuf. Ihre mutmaßliche Lebensweise wird manchmal mit Riesenfaultieren verglichen. Vermutlich haben sie mit den langen Krallen Äste herbeigezogen, um an die Nahrung zu gelangen.
Fortpflanzung und Entwicklung
 Skelett eines Oviraptoriden samt Gelege
Skelett eines Oviraptoriden samt GelegeWie vermutlich alle Dinosaurier und auch die heutigen Vögel waren die Theropoden eierlegend. Mehrere Skelette von Vertretern der Oviraptorosauria wurden über Gelegen entdeckt. Früher hielt man die Tiere für Eierfresser, daher auch der Name, der „Eierdieb“ bedeutet, heute interpretiert man die Funde dahingehend, dass die Eltern die Eier bebrütet haben. Die Nester bestanden aus bis zu 22 Eiern, das adulte Tier war über der Nestmitte positioniert und hielt die Arme um das Nest herum. Es wurden auch andere fossile Nester gefunden, die allerdings keiner bestimmten Gruppe zugeordnet werden können.
Die Entwicklung der Theropoda ist durch ein vermutlich relativ schnelles Wachstum charakterisiert, wobei die Größe vom Schlüpfling zum adulten Tier um das 10- bis 15-fache zunahm. Auch in ihrem Körperbau unterschieden sich die Jungtiere, so hatten sie andere Schädelproportionen mit kleineren Kiefern, einen kürzeren Nacken und längere Hinterbeine. Funde von Jungtieren sind aber nur von wenigen Vertretern bekannt, etwa von den Coelophysoidea Syntarsus und Coelophysis oder von Tyrannosaurus, sodass viele Angaben über die Entwicklung sehr vage bleiben müssen.
Systematik
Äußere Systematik
Die Theropoda werden mit einer stammlinienbasierten Definition (stem-based definition) beschrieben als „alle Taxa, die näher mit Passer domesticus [=Haussperling] als mit Cetiosaurus oxoniensis [ein Sauropode] verwandt sind“ [11]. Sie werden innerhalb der Dinosaurier in die Gruppe der Echsenbeckensaurier (Saurischia) eingeordnet, die daneben noch die Sauropodomorpha umfasst. Diese Abstammung lässt sich in folgendem Kladogramm wiedergeben:
Dinosaurier (Dinosauria) ├─Vogelbeckensaurier (Ornithischia) └─Echsenbeckensaurier (Saurischia) ├─Sauropodomorpha └─Theropoda
 Rekonstruktion von Eoraptor, einem systematisch umstrittenen basalen Dinosaurier
Rekonstruktion von Eoraptor, einem systematisch umstrittenen basalen DinosaurierEs gibt einige urtümliche Dinosaurier wie beispielsweise Eoraptor und die Herrerasauridae (mit Herrerasaurus und Staurikosaurus), deren systematische Einordnung umstritten ist. Für diese frühen Vertreter aus der Oberen Trias gibt es Untersuchungen mit unterschiedlichen Ergebnissen, so gelten sie manchmal als außerhalb der Dinosaurier stehend, manchmal als basale Vertreter der Theropoden und manchmal als basale Vertreter der Echsenbeckensaurier. Eine phylogenetische Untersuchung von Max C. Langer [12] ordnet diese umstrittenen Arten in die Echsenbeckensaurier ein, sieht die Theropoda aber als näher mit den Sauropodomorpha als mit ihnen verwandt, weswegen sie als basale Echsenbeckensaurier (und nicht als Theropoden) klassifiziert werden. Aufgrund des unvollständigen Materials herrscht über diese systematische Frage aber keine Einigkeit. [13]
Innere Systematik und Entwicklungsgeschichte
Wenn die oben erwähnten umstrittenen Vertreter zu den Theropoden gerechnet werden, wie von Sereno et al. [14] vorgeschlagen, dann gelten die übrigen Theropoden als Schwestergruppe dieser Vertreter und werden als Neotheropoda zusammengefasst.
 Ceratosaurus, ein Vertreter der Ceratosauria
Ceratosaurus, ein Vertreter der CeratosauriaAbgesehen davon lassen sich die Theropoda gut in zwei Gruppen einteilen, die Ceratosauria und die Tetanurae. Die Ceratosauria stellen dabei eine frühe Abspaltung dar, hier werden wiederum zwei Taxa unterschieden. Das sind zum einen die Coelophysoidea, eine Gruppe kleinerer Dinosaurier, die in der Oberen Trias und im Unteren Jura verbreitet war und abgesehen von den umstrittenen Gruppen die ältesten Theropoden sind. Das zweite Taxon der Ceratosauria sind die Neoceratosauria. Diese umfassen neben frühen Vertretern wie Ceratosaurus auch die Abelisauroidea, die in der Kreidezeit zu den dominanten Räubern auf den südlichen, aus dem Großkontinent Gondwana hervorgegangen Kontinenten zählten. Die Systematik innerhalb der Ceratosauria ist aber nicht unumstritten, ebenso ist unklar, ob sie monophyletisch sind.
Die Tetanurae („Steifschwänze“, aufgrund der durch Knochenstäbe verbunden und damit versteiften Schwanzwirbel) sind eine gut belegte, wahrscheinlich monophyletische Gruppe, die erstmals im frühen Jura erschien. Abgesehen von einigen basalen Vertretern sind innerhalb der Tetanurae zwei Entwicklungslinien zu erkennen, die Spinosauroidea und die Avetheropoda. Die Spinosauroidea (oder Megalosauroidea), deren Zusammengehörigkeit erst 1998 erkannt wurde, teilen sich in die Megalosauridae – ein Taxon mittelgroßer Theropoda aus dem Jura, zu denen unter anderem Megalosaurus und Eustreptospondylus gerechnet werden – und in die Spinosauridae, eine aus der Kreidezeit belegte Gruppe mit krokodilartig verlängertem Kopf, deren Vertreter sich möglicherweise zum Teil von Fischen ernährten.
Die Avetheropoda oder Neotetanurae sind durch ein zusätzliches Fenster im Oberkiefer und durch andere Merkmale im Schädel, dem Becken und den Schwanzwirbeln gekennzeichnet. Sie werden in zwei Untertaxa eingeteilt, die Carnosauria und die Coelurosauria. Diese beiden Bezeichnungen wurden früher für die Grobeinteilung der Theropda in größere und kleinere Vertreter verwendet, in modernen Systematiken bezeichnen sie eingeschränkte, genauer definierte Taxa.
Die Carnosauria sind relativ große Theropoda und unter anderem durch vergrößerte Nasenlöcher charakterisiert, sie umfassen drei größere Gruppen. Dies sind die Sinraptoridae, die vorwiegend aus dem späten Jura aus Asien bezeugt sind, die Allosauridae, unter anderem mit dem bekannten Allosaurus, und die Carcharodontosauridae, zu denen mit Carcharodontosaurus und Giganotosaurus einige der größen bekannten Theropoden gerechnet werden.
Die Coelurosauria sind das Taxon, bei dem laut Fossilbefund die Entwicklung der Federn einsetzte. Innerhalb dieser Gruppe stellen möglicherweise die Compsognathidae, eine Gruppe sehr kleiner Tiere aus dem Oberjura und der Unterkreide, die Schwestergruppe der übrigen Vertreter dar. Neben einigen basalen Vertretern wie Ornitholestes umfassen die Coelurosauria noch die Tyrannosauroidea, die Ornithomimosauria und die Maniraptora. Die Tyrannosauroidea, deren bekanntester Vertreter Tyrannosaurus ist, waren zum Teil sehr große Theropoden, bei denen die späteren Vertreter durch einen massiven Kopf und kurze Vordergliedmaßen gekennzeichnet waren. Die Ornithomimosauria bildeten ein Taxon laufvogelähnlicher, vermutlich pflanzen- oder allesfressender Dinosaurier.
 Troodon, ein Vertreter der Deinonychosauria
Troodon, ein Vertreter der DeinonychosauriaDie Maniraptora sind durch die meist stark verlängerten Vordergliedmaßen und Hände gekennzeichnet. Zu ihnen werden unter anderem die Oviraptorosauria, die Therizinosauria, die Deinonychosauria und die Vögel (Aves oder Avialae) gerechnet. Die Oviraptorosauria bilden ein Taxon meist zahnloser Dinosaurier, die Therizinosauria oder Segnosauria wiesen stark verlängerte Krallen an den Vordergliedmaßen auf und fraßen vermutlich Pflanzen. Die Deinonychosauria waren eher kleinere, räuberisch lebende Tiere, die durch eine große Kralle an der zweiten Zehe charakterisiert waren und zu denen unter anderem Velociraptor und Deinonychus gezählt werden. Auch die Vögel gehören nach phylogenetischen Gesichtspunkten zu den Maniraptora. Umstritten innerhalb der Maniraptora ist die Position der Alvarezsauridae, die vogelähnliche mit urtümlichen Merkmalen vereinen, aber aufgrund ihrer spezialisierten, stark verkürzten Arme nicht leicht zu klassifizieren sind.
Die Klassifikation der Theropoda ist weder abgeschlossen noch unumstritten. Viele Gruppen sind nur durch schwache Übereinstimmungen belegt, und neue Funde können neue Erkenntnisse liefern, die eine Neuordnung erfordern. Verschiedene Forscher vertreten teils stark voneinander abweichende Meinungen und Klassifikationssysteme. Ein mögliches Kladogramm der Theropoden sieht folgendermaßen aus: [15]
Theropoda ├─Ceratosauria │ ├─Coelophysoidea │ └─Neoceratosauria └─Tetanurae ├─Spinosauroidea │ ├─Spinosauridae │ └─Megalosauridae └─Avetheropoda ├─Carnosauria └─Coelurosauria ├─Compsognathidae └─Tyrannoraptora ├─Tyrannosauroidea └─Maniraptoriformes ├─Ornithomimosauria └─Maniraptora ├─N.N. │ ├─Oviraptorosauria │ └─Therizinosauria └─Paraves ├─Alvarezsauridae (Position sehr umstritten) └─Eumaniraptora ├─Deinonychosauria │ ├─Troodontidae │ └─Dromaeosauridae └─Vögel (Aves oder Avialae)
Die ältesten unzweifelhaften Theropoda – die Coelophysoidea – erschienen in der Obertrias im späten Carnium (vor rund 220 Millionen Jahren). Ab diesem Zeitpunkt sind die Theropoden nahezu aus dem gesamten Mesozoikum bekannt, die Nicht-Vogel-Theropoden sind beim Massenaussterben der Dinosaurier am Ende der Kreidezeit ausgestorben. (Für die Diskussionen der Gründe für dieses Aussterben siehe Kreide-Tertiär-Grenze und Das Aussterben der Dinosaurier.)
Die Entwicklung der Vögel
Die große Mehrzahl der Forscher geht davon aus, dass Vögel im kladistischen Sinn Theropoda und damit Dinosaurier sind. Das bedeutet, dass einige Theropoden wie die Deinonychosauria näher mit den Vögeln als mit den übrigen Vertretern dieser Gruppe verwandt sind und dass die Theropoda ohne die Vögel eine paraphyletische Gruppe sind, das heißt nicht alle Nachkommen eines gemeinsamen Vorfahren umfassen. Viele Merkmale, die innerhalb der rezenten Tiere nur bei Vögeln vorkommen (beispielsweise Federn, das Gabelbein oder hohle, pneumatisierte Knochen), finden sich auch bei einigen oder sogar allen Theropoda. Im Lauf ihrer Entwicklung haben sich die Vögel in einigen Merkmalen – vor allem in Verbindung mit der Flugfähigkeit – deutlich weiterentwickelt, beispielsweise in dem mit einem Brustbeinkiel versehenen Brustbein oder in der Verschmelzung der Mittelhandknochen zum Carpometacarpus. Es sind – vor allem aus der Kreidezeit – zahlreiche Mosaikformen bekannt, die diese graduelle Entwicklung deutlich werden lassen, beispielsweise der „Urvogel“ Archaeopteryx, Confuciusornis, die Enantiornithes und einige andere. Auch einige Taxa wie die Oviraptorosauria oder die Alvarezsauridae, die üblicherweise zu den Nicht-Vogel-Theropoden gezählt werden, werden in manchen Systematiken in die Vögel eingegliedert.
Eine Minderheit von Forschern, darunter der Paläoornithologe Alan Feduccia, vertritt die Meinung, die Vögel hätten sich nicht aus den Theropoden, sondern aus einer basalen Gruppe der Archosauria entwickelt und manche vogelähnlichen Theropoden seien keine Theropoden, sondern flugunfähige, an eine laufende Fortbewegung angepasste Vögel. Aufgrund der zahlreichen anatomischen Übereinstimmungen geht jedoch die Mehrheit der Wissenschaftler von einer Theropoden-Abstammung aus.
Theropoda und Menschen
Entdeckungs- und Forschungsgeschichte
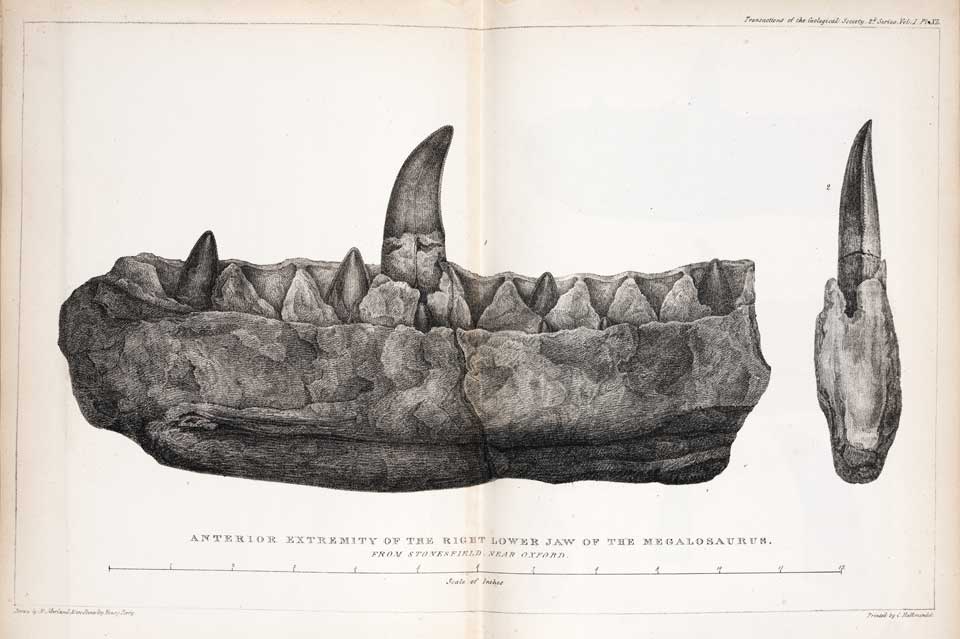
 Zeichnung des Unterkiefers von Megalosaurus von William Buckland
Zeichnung des Unterkiefers von Megalosaurus von William BucklandEin Teil eines Oberschenkelknochens – vermutlich von Megalosaurus – wurde bereits im Jahr 1677 gefunden, von Robert Plot untersucht und für den Knochen eines Riesen gehalten. Anfang des 19. Jahrhunderts wurden weitere Überreste dieser Gattung, darunter ein Kieferfragment mit Zähnen, gefunden, und 1824 von William Buckland als Megalosaurus wissenschaftlich erstbeschrieben. 1842 prägte Richard Owen den Begriff Dinosauria für diese und zwei weitere Gattungen.
Mitte des 19. Jahrhunderts wurden mit Troodon in Nordamerika und Compsognathus (und Archaeopteryx) in Solnhofen (Deutschland) die nächsten Theropoden entdeckt. Die „Bone Wars“, eine Auseinandersetzung zwischen den Paläontologen Edward Drinker Cope und Othniel Charles Marsh Ende des 19. Jahrhunderts, brachten weitere Vertreter dieser Gruppe ans Tageslicht, so beschrieb Cope Coelophysis und Marsh Allosaurus, Coelurus und Ceratosaurus. 1881 prägte Marsh den Begriff Theropoda („Tierfüßler“), zunächst für alle bekannten Dinosaurier aus der Trias und die fleischfressenden Dinosaurier aus dem Jura und der Kreide. Anfang des 20. Jahrhunderts wurden weitere Funde gemacht, darunter Tyrannosaurus und Albertosaurus in Nordamerika, Elaphrosaurus in Tendaguru (Tansania) sowie Spinosaurus und Carcharodontosaurus von Ernst Stromer von Reichenbach in Ägypten.
In den Jahren 1914 und 1920 erschienen systematische Studien von Friedrich von Huene. Dieser teilt die Theropoda in zwei Gruppen, die kleinen „Coelurosauria“ und die großen „Carnosauria“. Diese Systematik war bis in die 1980er-Jahre gültig, gilt jedoch heute als veraltet, die Begriffe werden aber für anders definierte Taxa immer noch verwendet. Ebenfalls 1920 veröffentlichte Charles W. Gilmore eine erste umfassende Studie zu nordamerikanischen Theropoda. [16] Die Zwischenkriegszeit sah weitere Entdeckungen von Theropoden, darunter in Nordamerika Dromaeosaurus und in Asien beispielsweise Oviraptor und Velociraptor. 1932 veröffentlichte Friedrich von Huene seine Monographie über die Saurischia [17], in der er die Taxonomie, die Verteilung und die Entwicklung der Theropoda zusammenfasst. Dieses Werk blieb bis in die 1960er-Jahre maßgeblich.
Seit den 1960er-Jahren wurde die Vielfalt und Komplexität der Theropoda zunehmend erkannt, viele alte Gattungen wurden neu untersucht. Die spektakulärste Entdeckung in dieser Zeit war wohl Deinonychus durch John H. Ostrom. Dessen Arbeiten sollten das bisherige Bild der Theropoden auf den Kopf stellen. Er sah in ihnen agile, flinke Tiere statt der bisher vorherrschenden Meinung, sie seien plump und langsam gewesen. Ostrom hat auch die Thesen zur Warmblütigkeit zumindest mancher Dinosaurier und zur Abstammung der Vögel neu aufgeworfen.
Auch in den 1970er- und 1980er-Jahren wurden viele neue Theropoden entdeckt, zum Beispiel Tarbosaurus, die Abelisauroidea auf den südlichen Kontinenten oder Baryonyx. 1986 führte Jacques Gauthier erstmals die Kladistik in die Dinosauriersystematik ein [18], er war es auch, der die Theropoda erstmals definierte als „Vögel und alle Saurischia, die näher mit Vögeln als mit Sauropodomorpha verwandt sind“. Im Jahr 1996 wurde mit Sinosauropteryx erstmals ein gefiederter Dinosaurier beschrieben, später wurden auch andere wie Protarchaeopteryx, Caudipteryx, Shuvuuia, Sinornithosaurus und Microraptor entdeckt. Neben zahlreichen neuen Funden und verbesserten Untersuchungen alter Theropoden erschienen auch weitere kladistische systematische Studien, unter anderem von Thomas R. Holtz und Kevin Padian. In jüngerer Zeit sind neben anatomischen und systematischen Fragestellungen auch paläoökologische Untersuchungen bedeutend geworden, welche die Lebensweise dieser Tiere und ihre Interaktion mit anderen Lebewesen erforschen. Es steht zu erwarten, dass in Zukunft durch neue Funde und Analysen weitere Informationen über Aussehen, Lebensweise und Systematik dieser Tiere gewonnen werden können.
Theropoda in der Kultur
Seit die Dinosaurier im 19. Jahrhundert erstmals ins Licht der Öffentlichkeit rückten, erfreuen sie sich weltweit großen Interesses und hoher Beliebtheit. Zu den bekanntesten und populärsten Dinosauriern zählen dabei auch viele Theropoda. Das spiegelt sich auch in ihrer Rolle in fiktiven Werken (von Arthur Conan Doyles Buch „Die vergessene Welt“ („The Lost World“, 1912) bis zu den modernen Jurassic-Park-Verfilmungen) oder in populärwissenschaftlichen Werken, zum Beispiel der BBC-Serie „Dinosaurier – Im Reich der Giganten“ (englischer Titel: „Walking with dinosaurs“), wider. Zu den Gründen für die Popularität dieser Tiere siehe auch Dinosaurier in den Medien.
Literatur
- David Weishampel, Peter Dodson und Halszka Osmólska (Hrsg.): The Dinosauria. University of California Press, 2004, ISBN 0520242092.
- David Fastovsky und David Weishampel: The Evolution and Extinction of the Dinosaurs. Cambridge University Press, 2005, ISBN 0521811724.
- Wilfried Westheide, Reinhard Rieger (Hrsg.): Spezielle Zoologie. Teil 2: Wirbel- oder Schädeltiere. Spektrum Akademischer Verlag, Heidelberg/Berlin 2004, ISBN 3-8274-0307-3.
Weblinks
- Theropoda auf palaeos.com
- James O. Farlow, Stephen M. Gatesy, Thomas R. Holtz, Jr., John R. Hutchinson and John M. Robinson (2000): Theropod Locomotion. „American Zoologist“, 2000 40(4):640-663 - Volltext
Einzelnachweise
- ↑ R. O. Prum (1999): Development and evolutionary origin of feathers. In: Journal of Experimental Zoology. Nr. 285, S. 291–306
R. O. Prum und A. H. Brush: Which came first, the feather or the bird? In: Scientific American. Nr. 288 (2003), S. 84–93 - ↑ X. Xu, M. A. Norell, X. Kuang, X. Wang, Q. Zhao& C. Jia: Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids. In: Nature. Nr. 431 (2004), S. 680–684
- ↑ W.D. Maxwell und J. H. Ostrom: Taphonomy and paleobiological implications of Tenontosaurus-Deinonychus associations. In: Journal of Vertebrate Paleontology. Nr. 15 (1995), S. 707–712 (Abstract)
- ↑ K. Carpenter: Variation in Tyrannosaurus rex . In: K. Carpenter und P. J. Currie: Dinosaur Systematics: Approaches and Perspectives. Cambridge University Press, New York 1990, S. 141–145
- ↑ R. M. Alexander: Estimates of speeds of dinosaurs. In: Nature. Nr. 261 (1976), S. 129–130
- ↑ R. A. Thulborn: Speeds and gaits of dinosaurs. In: Palaeogeography Palaeoclimatology Palaeoecology. Nr. 38 (1982), S. 227–256
- ↑ J. A. Hopson: Relative brain size in dinosaurs; implications for dinosaurian endothermy. In: R. D. K. Thomas und E. C. Olson (Hrsg.): A Cold Look at Warm-Blooded-Dinosaurs. In: American Association for the Advancement of Science Symposium. Nr. 27 (1980), S. 287–310
- ↑ Angaben zum Enzephalsationsquotient der Theropoda
- ↑ David Fastovsky und Joahua B. Smith: Dinosaur Paleoecology. In: 'The Dinosauria. (2004), S. 620–624
- ↑ J. H. Ostrom: Deinonychus, The Ultimate Killing Machine. In G. D. Rosenberg u. D. L. Wolberg (Hrsg.): Dino Fest. Proceedings of a Conference for the General Public: Paleontological Society Special Publications, 7 (1994), S. 127-138.
- ↑ “all taxa closer to Passer domesticus than to Cetiosaurus oxoniensis”. T. R. Holtz & H. Osmólska: Saurischis. In: The Dinosauria. (2004), S. 22, bezugnehmend auf Arbeiten von K. Padian und C. L. May bzw. T. Maryańska
- ↑ Max C. Langer: Basal Saurischia. In: Weishampel, Dodson und Osmólska (Hrsg.): The Dinosauria. (2004), S. 40–44
- ↑ Das Problem der Diagnose urtümlicher Theropoden ausführlich und auf letztem Stand diskutiert in: P. C. Sereno: The phylogenetic relationships of early dinosaurs: A comparative report. In: Historical Biology, 19 (1), 2007, S. 145-155.
- ↑ P. C. Sereno, C. A. Forster, R. R. Rogers und A. M. Monetta: Primitive dinosaur skeleton from Argentina and the early evolution of Dinosauria. In: Nature. Nr. 361 (1993), S. 64–66
- ↑ vereinfacht nach Weishampel, Dodson und Osmólska: The Dinosauria. (2004)
- ↑ Charles W. Gilmore: Osteology of the Carnivorous Dinosauria in the United States National Museum. In: United States National Museum Bulletin. Nr. 110 (1920), S. 1–154
- ↑ Friedrich von Huene: Die fossile Reptil-Ordnung Saurischia, ihre Entwicklung und Geschichte. In: Monogr. Geol. Palaeontol. (ser. 1) 4; S. 1–361 (1932)
- ↑ J. Gauthier: Saurischian Monophyly and the origin of birds. In: Memoirs of the California Academy of Sciences. Nr. 8 (1986), S. 1–55















Wikimedia Foundation.