- Reversible Inhibition
-
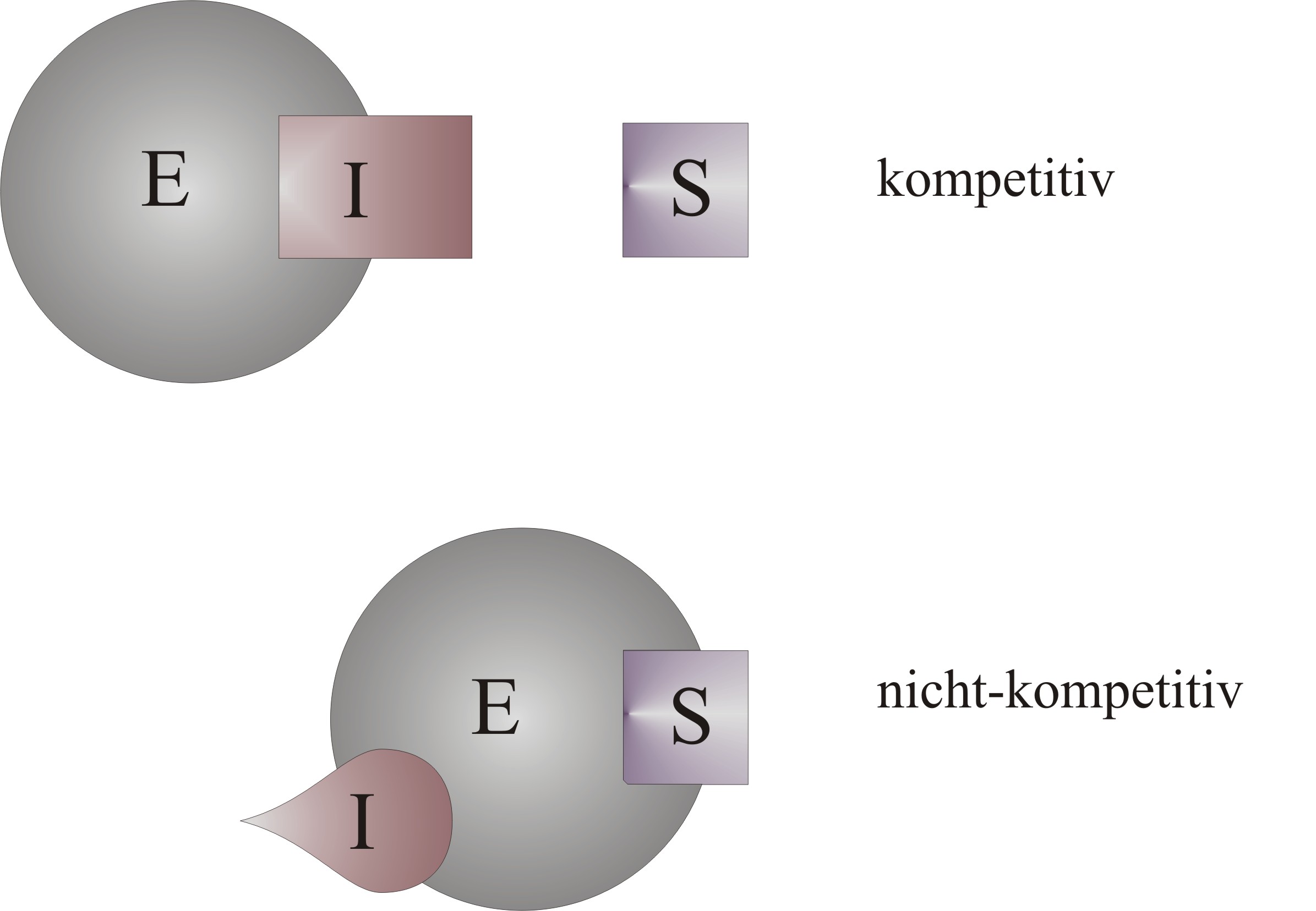
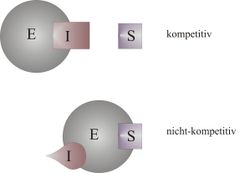 Bindungsort des Hemmstoffes bei einer kompetitiven bzw. nicht-kompetitiven Hemmung
Bindungsort des Hemmstoffes bei einer kompetitiven bzw. nicht-kompetitiven HemmungEnzymhemmung ist die negative Beeinflussung einer enzymatischen Reaktion durch einen Hemmstoff, der Inhibitor genannt wird. Dabei wird die Geschwindigkeit der Reaktion herabgesetzt. Die Hemmstoffe können an unterschiedliche an der Reaktion beteiligten Stoffe binden, wie zum Beispiel an das Enzym oder das Substrat. Auch der Bindungsort am Enzym kann vom aktiven Zentrum, an dem das Substrat bindet, bis hin zu anderen Stellen, die für die Aktivität des Enzyms wichtig sind, variieren.
Inhaltsverzeichnis
Grundlagen
Enzyme sind für jeden Organismus essentiell. Sie sind an jedem Stoffwechselprozess beteiligt und wirken als Katalysatoren für die meisten Reaktionen. Um diese Prozesse regulieren zu können, brauchen die Zellen bestimmte Mechanismen, welche die Aktivität der Enzyme beeinflussen. Manche Enzyme können durch Modifikationen angeschaltet, also aktiviert werden. Beispielsweise wird die bei der Verwertung von Glukose benötigte Pyruvatkinase durch Phosphorylierung reguliert, d. h. dem Enzym kann eine Phosphorylgruppe angehängt werden. Diese phosphorylierte Form der Pyruvatkinase ist wenig aktiv. Wurde das Enzym jedoch nicht durch eine Phosphorylgruppe modifiziert, besitzt es volle Aktivität.
Die Aktivität von Enzymen kann auch durch Bindung von bestimmten Stoffen beeinflusst werden. Diese Stoffe werden Effektoren genannt. Je nach dem wie Effektoren auf ein Enzym wirken, werden sie Aktivatoren oder Inhibitoren genannt. Aktivatoren erhöhen die Aktivität von Enzymen, d. h. sie fördern die durch das Enzym katalysierte Reaktion. Inhibitoren beeinflussen das Enzym negativ. Sie senken die Aktivität und somit hemmen sie die durch das Enzym katalysierte Reaktion. Das wird Enzymhemmung genannt.
Es gibt auch noch andere Arten zur Verringerung der Enzymaktivität, welche aber nicht zur Enzymhemmung gehören. Dazu zählen Beeinflussungen durch die Temperatur, den pH-Wert, die Ionenstärke oder Lösungsmitteleffekte. Diese Faktoren wirken unspezifisch, also zum Beispiel durch Veränderung der Konformation des Enzyms, also der räumlichen Struktur, ohne spezifisch auf das aktive Zentrum des Enzyms einzuwirken. Das aktive Zentrum ist sozusagen der Ort am Enzym, wo das Substrat bindet, also auch die Reaktion stattfindet.
Einteilung der Enzymhemmung
Die Enzymhemmung unterteilt sich abhängig von der Bindung des Hemmstoffes (Inhibitors) in reversible und irreversible Inhibition. Bei der reversiblen Enzymhemmung kann der Inhibitor wieder vom Enzym abgespaltet oder verdrängt werden. Er bindet nicht fest an das Enzym. Diese Form der Enzymhemmung wird zur Regulation verschiedener Stoffwechselprozesse genutzt, die zeitweise nicht ablaufen sollen. Zum Beispiel wird die Glykolyse zur Energiegewinnung aus Glucose genutzt. Ein Enzym daraus wurde oben schon erwähnt, die Pyruvatkinase. Ein weiteres Enzym der Glykolyse ist die Phosphofructokinase. Ist in der Zelle viel Energie vorhanden, hat sie diese in Form von Adenosintriphosphat (ATP) gespeichert. Dieses ATP hemmt als Inhibitor die Phosphofructokinase und auch die Pyruvatkinase. Somit wird keine Glukose mehr in Energie, also ATP, umgewandelt. Diese spezielle Form der Enzymhemmung, bei der das Endprodukt das Enzym hemmt, welches zu der Synthese dieses Stoffes führt, nennt man Endprodukthemmung oder Feedback-Hemmung.
Bei der irreversiblen Hemmung bindet der Inhibitor so fest, dass er nicht mehr vom Enzym zu lösen ist. Die Aktivität des Enzyms geht verloren. Die irreversible Hemmung findet man zum Beispiel bei Pilzen, die Antibiotika zu deren Schutz produzieren. Diese Antibiotika hemmen oft irreversibel bestimmte Stoffwechselwege wie die Proteinbiosynthese.
Reversible Enzymhemmung
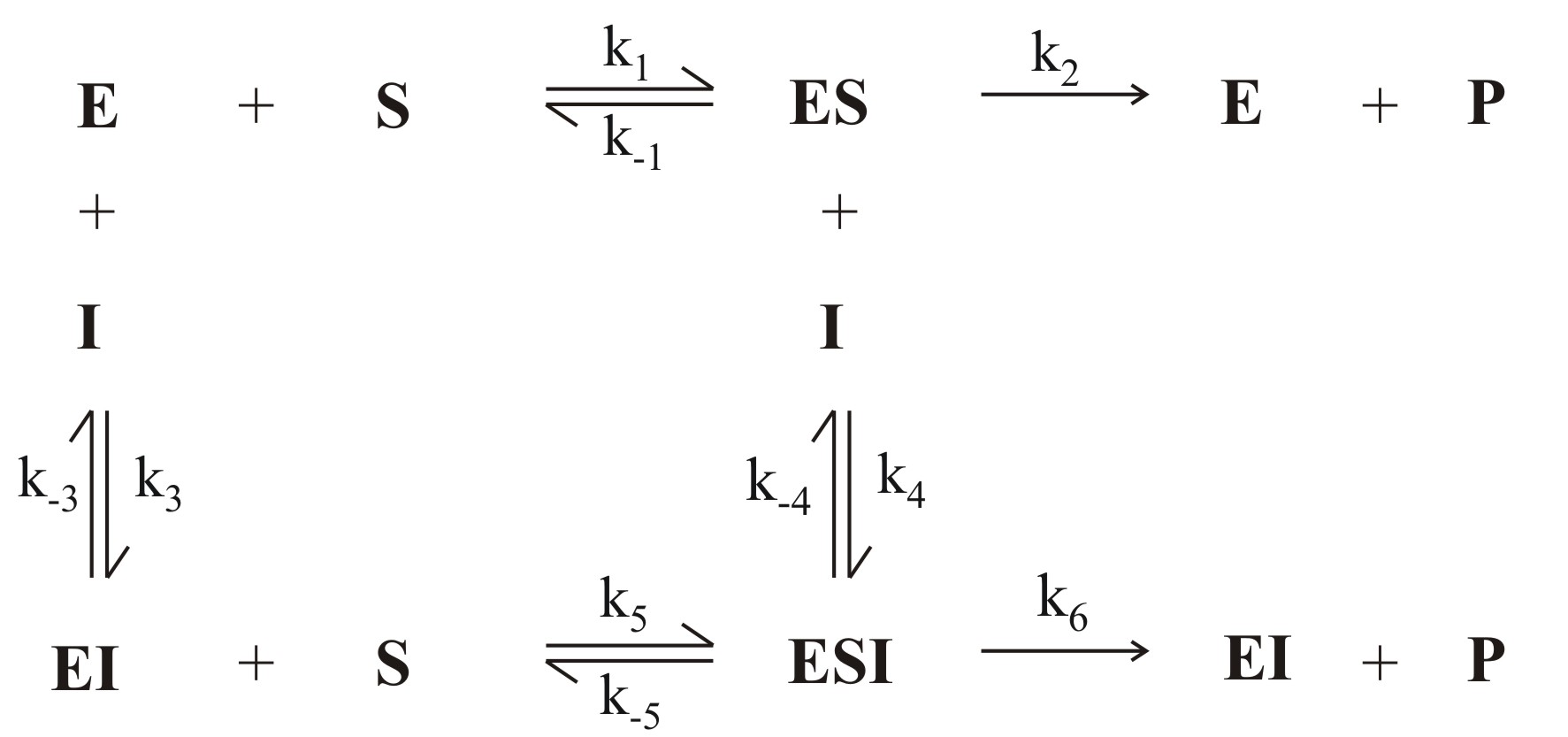
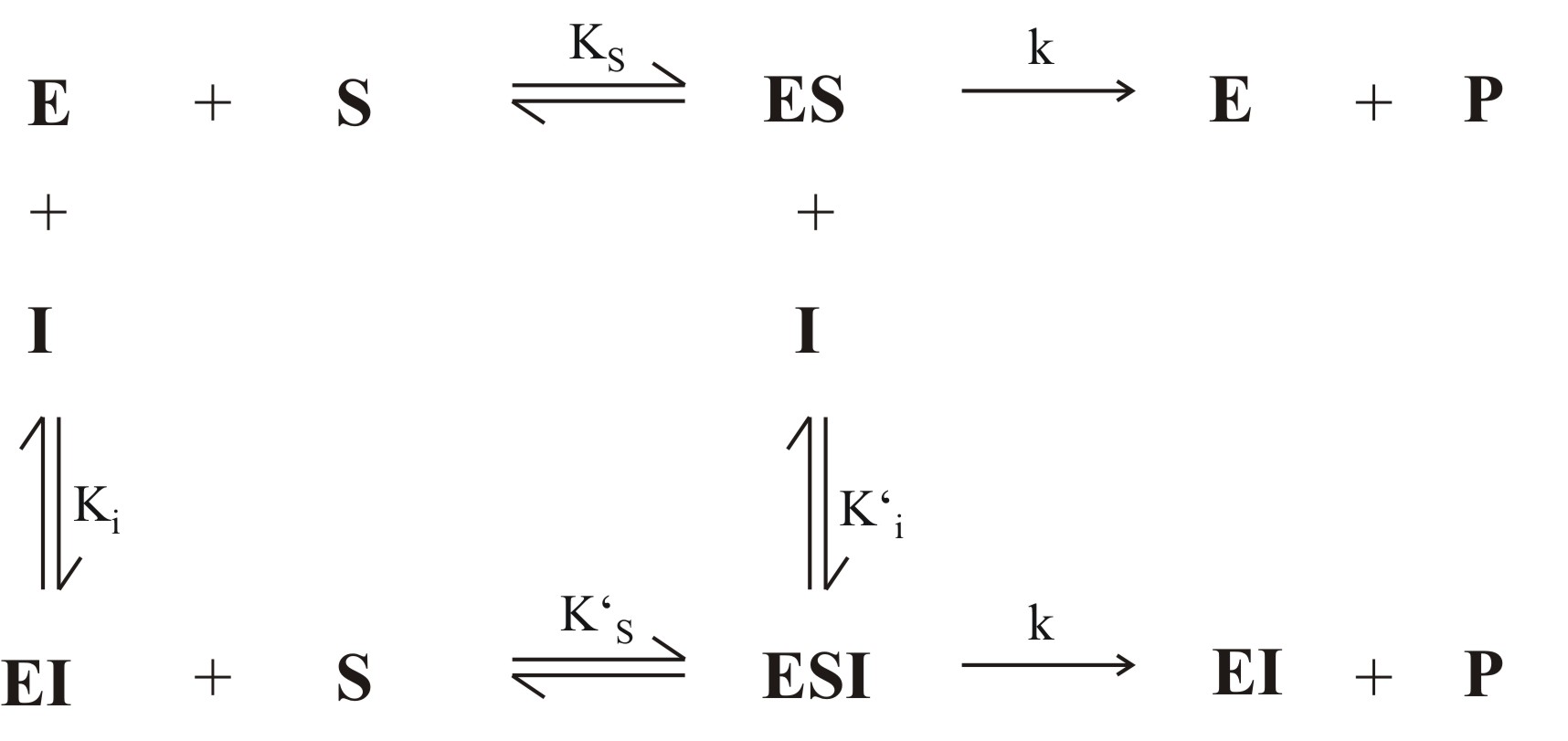
 Abb. 1: Allgemeiner Mechanismus der Enzymhemmung (E:Enzym, S: Substrat, P: Produkt, I: Inhibitor, ES: Enzym-Substrat-Komplex, EI: Enzym-Inhibitor-Komplex, ESI: Enzym-Substrat-Inhibitor-Komplex)
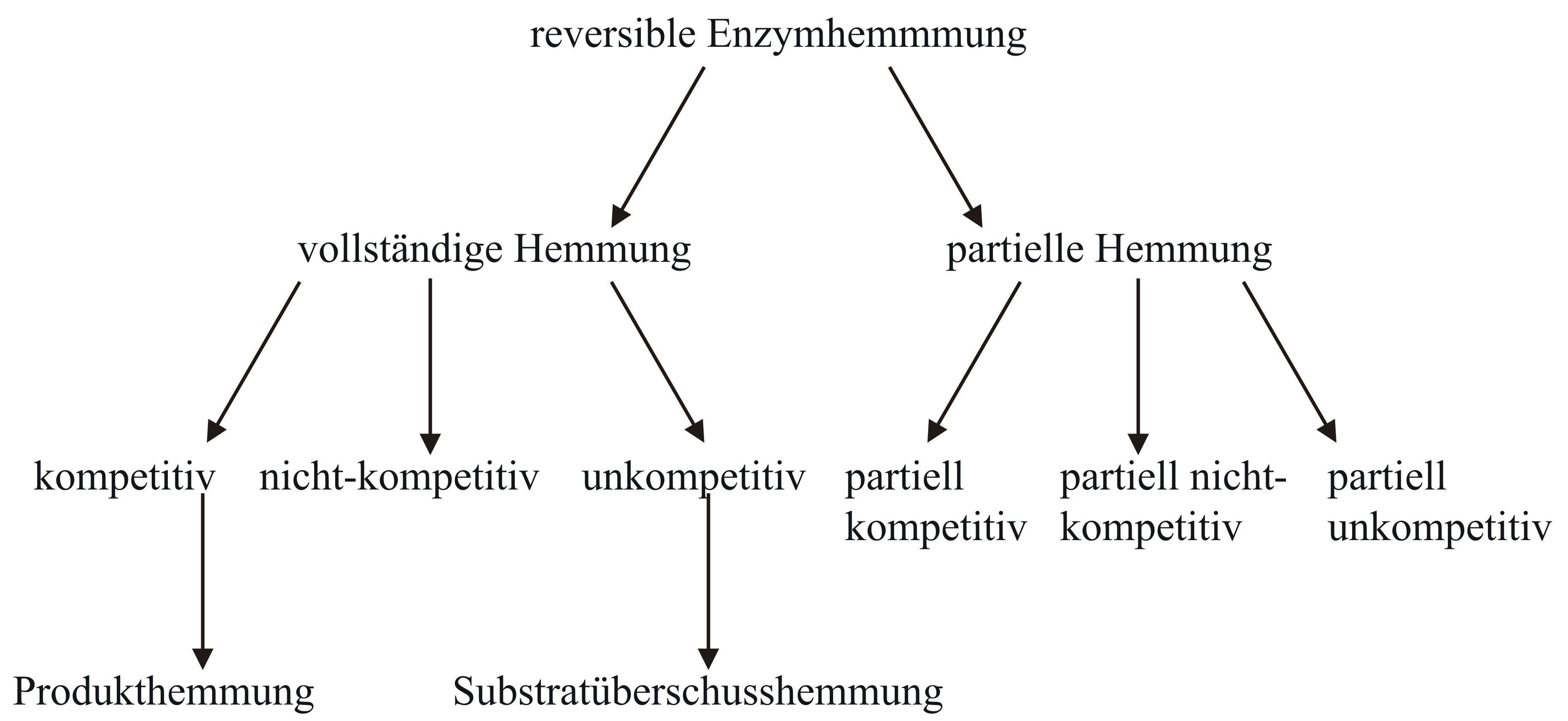
Abb. 1: Allgemeiner Mechanismus der Enzymhemmung (E:Enzym, S: Substrat, P: Produkt, I: Inhibitor, ES: Enzym-Substrat-Komplex, EI: Enzym-Inhibitor-Komplex, ESI: Enzym-Substrat-Inhibitor-Komplex)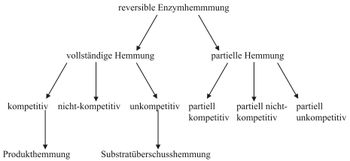 Abb. 2: Überblick über verschiedene Mechanismen der reversiblen Hemmung
Abb. 2: Überblick über verschiedene Mechanismen der reversiblen HemmungBei der reversiblen Enzymhemmung bindet der Inhibitor I reversibel an das Enzym E und senkt somit dessen Aktivität bzw. die Geschwindigkeit der Reaktion des Substrates S zum Produkt P. Der Inhibitor kann aber zum Beispiel vom Substrat wieder verdrängt werden. Mathematisch gesehen gibt die Geschwindigkeitskonstante k2 die „Schnelligkeit“ der ungehemmten Reaktion an. Bindet nun der Inhibitor an den Enzym-Substrat-Komplex ES so wird die Reaktion durch die Geschwindigkeitskonstante k6 definiert. k6 ist kleiner als k2. Da es sich um reversible Enzymhemmung handelt, stellt sich bei den Reaktionen zu ES, dem Enzym-Inhibitor-Komplex EI und dem Enzym-Substrat-Inhibitor-Komplex ESI ein Gleichgewicht ein. Alle Geschwindigkeitskonstanten auch für die Gleichgewichte sind in Abb. 1 an den entsprechenden Reaktionspfeilen abgebildet.
Im Fließgleichgewicht kann die Reaktionsgeschwindigkeit sogar rechnerisch bestimmt werden:
Durch Umformulieren erhält man folgende für die reversible Enzymhemmung allgemein gültige Gleichung für die Reaktionsgeschwindigkeit:
Die eingeführte Konstante V1 entspricht k2[E0] und die Konstante V2 = k6[E0]. Die Gleichgewichtskonstanten Kic und Kiu lassen sich aus Abb. 1 ableiten: Kic = k − 3 / k3 und Kiu = k − 4 / k4.
In Abb. 2 sind die wichtigsten Hemm-Mechanismen der reversiblen Enzymhemmung aufgeführt. Die Einteilung erfolgte in vollständige und partielle Hemmung, die sich in V2 unterscheiden. Bei der vollständigen Enzymhemmung liegt V2 bei 0, bei der partiellen ist dieser Wert ungleich 0. Das heißt das Enzym behält bei einer partiellen Hemmung seine katalytische Aktivität bei, die allerdings durch den Hemmstoff beeinflusst wird. Bei der vollständigen Enzymhemmung hingegen kann der ESI-Komplex nicht mehr an der Reaktion teilnehmen und ist somit inaktiv.
Die Produkthemmung ist ein Sonderfall der kompetitiven Hemmung, denn der Inhibitor entspricht dem Produkt der Reaktion. Auch die Substratüberschusshemmung ist ein Sonderfall der unkompetitiven Hemmung. Das Substrat ist in diesem Fall der Hemmstoff, wenn dieses in hoher Konzentration vorliegt.
Kompetitive Hemmung
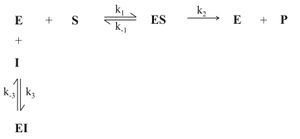 Abb. 3: Mechanismus der kompetitiven Hemmung (E: Enzym, S: Substrat, P: Produkt, ES: Enzym-Substrat-Komplex, I: Inhibitor, EI: Enzym-Inhibitor-Komplex)
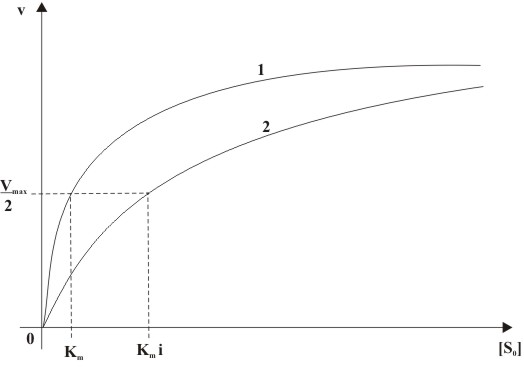
Abb. 3: Mechanismus der kompetitiven Hemmung (E: Enzym, S: Substrat, P: Produkt, ES: Enzym-Substrat-Komplex, I: Inhibitor, EI: Enzym-Inhibitor-Komplex) Abb. 4: Abhängigkeit der Anfangsgeschwindigkeit (v) von der Ausgangssubstratkonzentration([S0]) einer kompetitiven Hemmung mit (2) und ohne (1) Inhibitorzugabe
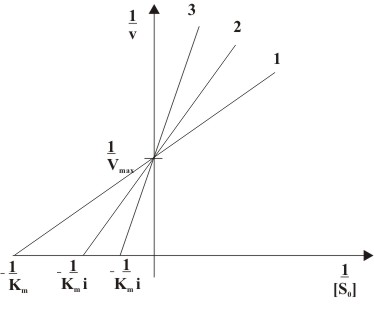
Abb. 4: Abhängigkeit der Anfangsgeschwindigkeit (v) von der Ausgangssubstratkonzentration([S0]) einer kompetitiven Hemmung mit (2) und ohne (1) Inhibitorzugabe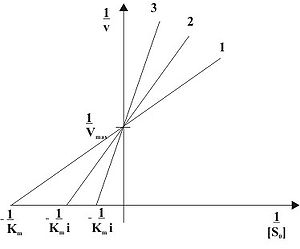 Abb. 5: Doppelt reziproke Auftragung einer kompetitiven Hemmung in Anwesenheit unterschiedlicher Inhibitorkonzentrationen ( 1: [I]=0, 2: [I]>0, 3: [I]>[I]von2)
Abb. 5: Doppelt reziproke Auftragung einer kompetitiven Hemmung in Anwesenheit unterschiedlicher Inhibitorkonzentrationen ( 1: [I]=0, 2: [I]>0, 3: [I]>[I]von2)Kompetitive Inhibitoren sind Substanzen, die mit dem Substrat um die Bindungsstelle im aktiven Zentrum des Enzyms konkurrieren. Sie werden nicht umgesetzt und können dadurch vom Substrat wieder verdrängt werden. Kompetitive Inhibitoren haben oft hohe Ähnlichkeit mit dem Substrat. Der Mechanismus der kompetitiven Hemmung ist in Abb. 3 dargestellt. Deutlich zu sehen ist, dass das Enzym E das Substrat S und den Inhibitor I nicht gleichzeitig binden kann. Durch die reversible Bindung von S bzw. I an E entsteht ein Gleichgewicht zwischen freiem Enzym E, dem Enzym-Substrat-Komplex ES und dem Enzym-Inhibitor-Komplex EI. Unter der Voraussetzung, dass sowohl das Substrat S als auch der Inhibitor I in sehr viel höheren Konzentrationen als das Enzym E vorliegen, kann man für den steady-state-Zustand folgende Reaktionsgeschwindigkeitsgleichung formulieren:
Die enthaltenen Konstanten sind so definiert:
![V_{\max} = k_2 \, [E_0]](/pictures/dewiki/51/3ee4ec016c901ba835f9cdeb46c64590.png) ,
, 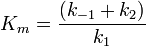
und
![i = 1 + \left(\frac{k_3[I_0]}{k_{-3}}\right)](/pictures/dewiki/48/0b710fd1d2362cc45b191217aca4cf2a.png) .
.
Die Reaktionsgeschwindigkeit ist in Abhängigkeit von der Ausgangssubstratkonzentration S0 in Ab- und Anwesenheit des kompetitiven Inhibitors in Abb. 4 dargestellt. Die Michaelis-Menten-Konstante wird in Gegenwart des Inhibitors um den Faktor i erhöht. Die Maximalgeschwindigkeit Vmax bleibt jedoch unverändert.
Die Linearisierung der Michaelis-Menten-Auftragung wird durch den Reziprokwert der Gleichung erreicht.
Die doppelt reziproke Auftragung, also die Auftragung von 1 / v gegen 1 / [S0] in Gegenwart unterschiedlicher Konzentrationen des Hemmstoffes ist in Abb. 5 dargestellt und wird Lineweaver-Burk-Auftragung genannt. Die Steigung des Graphen erhöht sich in Anwesenheit des Inhibitors um i. Da Vmax unverändert bleibt, schneiden sich alle Geraden auf der Ordinate im Punkt 1 / Vmax. Die Schnittpunkte mit der Abszisse spiegeln den Wert − 1 / Kmi wider.
Die oben beschriebenen Gleichungen wurden für die kompetitive Hemmung abgeleitet, bei der die gleichzeitige Bindung von Substrat und Hemmstoff ausgeschlossen wird. Die Bedingungen für eine kompetitive Hemmung können aber auch erfüllt werden, wenn der Inhibitor nicht die gleiche Bindungsstelle am Enzym einnimmt wie das Substrat. Eine Bindung im aktiven Zentrum, wodurch die Substratbindung sterisch eingeschränkt ist, führt auch zum kompetitiven Hemmtyp.
Inhibierung durch ein konkurrierendes Substrat
In diesem speziellen Fall von reversibler Enzymhemmung ist das Enzym in der Lage zwei Reaktionen zu katalysieren, also zwei verschiedene Substrate zu binden. Das Enzym E bindet das Substrat A und setzt dieses zum Produkt P um. Die Bindung des Substrates B an das Enzym E führt zur Bildung von Produkt Q. Somit wird die Reaktionsgeschwindigkeit vA durch Zugabe des Substrates B verringert, da auch das zweite Substrat B gebunden und umgesetzt werden kann. Das eine Substrat wirkt bei dieser Konkurrenzreaktion auf die Reaktionsgeschwindigkeit der Reaktion des jeweils
anderen Substrates als kompetitiver Inhibitor. Dadurch ergeben sich folgende
Geschwindigkeitsgleichungen:Nicht-kompetitive Hemmung
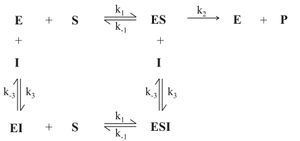 Abb. 6: Mechanismus der nicht-kompetitiven Hemmung (E: Enzym, S: Substrat, P: Produkt, I: Inhibitor, ES: Enzym-Substrat-Komplex, EI: Enzym-Inhibitor-Komplex, ESI: Enzym-Substrat-Inhibitor-Komplex)
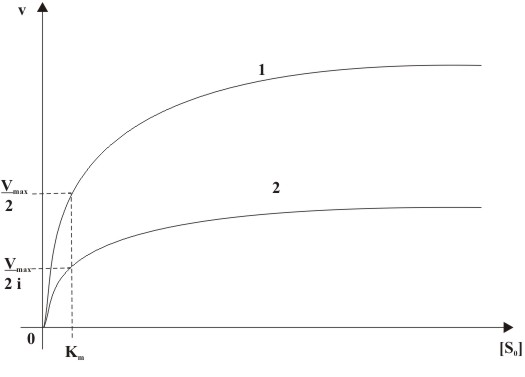
Abb. 6: Mechanismus der nicht-kompetitiven Hemmung (E: Enzym, S: Substrat, P: Produkt, I: Inhibitor, ES: Enzym-Substrat-Komplex, EI: Enzym-Inhibitor-Komplex, ESI: Enzym-Substrat-Inhibitor-Komplex) Abb. 7: Abhängigkeit der Anfangsgeschwindigkeit (v) von der Ausgangssubstratkonzentration([S0]) einer nicht-kompetitiven Hemmung mit (2) und ohne (1) Inhibitorzugabe
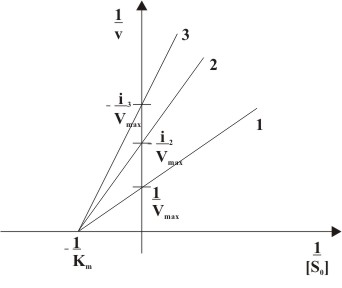
Abb. 7: Abhängigkeit der Anfangsgeschwindigkeit (v) von der Ausgangssubstratkonzentration([S0]) einer nicht-kompetitiven Hemmung mit (2) und ohne (1) Inhibitorzugabe Abb. 8: Doppelt reziproke Auftragung einer nicht-kompetitiven Hemmung in Anwesenheit unterschiedlicher Inhibitorkonzentrationen ( 1: [I]=0, 2: [I]>0, 3: [I]>[I]von2)
Abb. 8: Doppelt reziproke Auftragung einer nicht-kompetitiven Hemmung in Anwesenheit unterschiedlicher Inhibitorkonzentrationen ( 1: [I]=0, 2: [I]>0, 3: [I]>[I]von2)Bei der nicht-kompetitiven Hemmung wird durch die Bindung des Inhibitors I an das Enzym E die Substratbindung nicht beeinflusst. Der Inhibitor I ist somit in der Lage sowohl an das freie Enzym E als auch an den Enzym-Substrat-Komplex ES zu binden, d. h. der Inhibitor bindet nicht im Substrat bindenenden Teil des Enzyms, dem aktiven Zentrum. Das Substrat kann mit dem Enzym-Inhibitor-Komplex EI ebenfalls eine Reaktion eingehen, jedoch der gebildete Enzym-Inhibitor-Substrat-Komplex EIS ist nicht in der Lage das Produkt P abzuspalten. Der Reaktionsmechanismus ist genauer in Abb. 6 dargestellt.
Unter steady-state-Bedingungen kann eine einfache Geschwindigkeitsgleichung hergeleitet werden:
Die Konstanten Km und i sind folgender Maßen definiert: Km = k − 1 / k1 und i = 1 + (k3[I0] / k − 3). Aus der Geschwindigkeitsgleichung kann man ableiten, dass der nicht-kompetitive Inhibitor in Anwesenheit die Maximalgeschwindigkeit Vmax um den Faktor 1 / i verringert. Der Km-Wert für das Substrat bleibt unverändert.
Die Auftragung nach Lineweaver-Burk erfolgt unter Bildung der reziproken Reaktionsgeschwindigkeit nach dieser Formel:
Die Lineweaver-Burk-Auftragung ist in Abb. 8 dargestellt. Der Anstieg nach Zugabe des nicht-kompetitiven Inhibtiors ist i-fach höher als ohne Inhibition. Der Ordinatenschnittpunkt jeder Gerade liegt bei i / Vmax. Die Geraden für Reaktionen unterschiedlicher Inhibitorkonzentrationen schneiden sich in genau einem Punkt auf der Abszisse, im Wert − 1 / Km.
In manchen Fällen der nicht-kompetitiven Hemmung weicht das Verhalten des Inhibitors etwas vom „Normalfall“ ab. Die Umsetzung des Inhibitors mit dem Enzym erfolgt dann sehr viel schneller als die des Substrates. Bei niedrigen Substratkonzentrationen ist die Verminderung der Maximalgeschwindigkeit nicht so stark. Daher ergibt sich für die Bedingungen
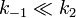 und Km = k2 / k1 folgende Geschwindigkeitsgleichung:
und Km = k2 / k1 folgende Geschwindigkeitsgleichung:![v = \frac{V_{\max}\;[S_0]}{[S_0] \cdot i + K_m}](/pictures/dewiki/53/52d6906e834b20ec95fac55658cf11e7.png) .
.
Bei Ermittlungen des Hemmtypes eines Inhibitors sollte man mit einem solchen Verhalten rechnen.
Unkompetitive Hemmung
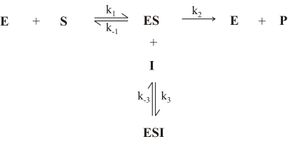 Abb. 9: Allgemeiner Mechanismus der unkompetitiven Enzymhemmung (E: Enzym, S: Substrat, P: Produkt, I: Inhibitor, ES: Enzym-Substrat-Komplex, ESI: Enzym-Substrat-Inhibitor-Komplex)
Abb. 9: Allgemeiner Mechanismus der unkompetitiven Enzymhemmung (E: Enzym, S: Substrat, P: Produkt, I: Inhibitor, ES: Enzym-Substrat-Komplex, ESI: Enzym-Substrat-Inhibitor-Komplex)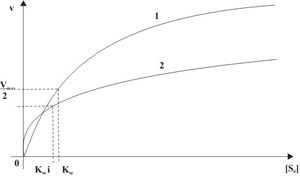 Abb. 10: Abhängigkeit der Anfangsgeschwindigkeit (v) von der Ausgangssubstratkonzentration([S0]) einer unkompetitiven Hemmung mit (2) und ohne (1) Inhibitorzugabe
Abb. 10: Abhängigkeit der Anfangsgeschwindigkeit (v) von der Ausgangssubstratkonzentration([S0]) einer unkompetitiven Hemmung mit (2) und ohne (1) InhibitorzugabeGelegentlich kommt neben der kompetitiven und der nicht-kompetitiven Hemmung auch der unkompetitive Hemmtyp vor. Dabei geht der Inhibitor nur mit dem Enzym-Substrat-Komplex ES eine Reaktion ein, wie in Abb. 9 zu sehen ist. Die für diesem Mechanismus unter steady-state-Bedingungen abgeleitete Gleichung für die Reaktionsgeschwindigkeit lautet wie folgt:
Die Konstanten sind so definiert: i = 1 + k3[I0] / k − 3 und Km = (k − 1 + k2) / k1. Aus der in Abb. 10 dargestellten Michaelis-Menten-Auftragung einer unkompetitiven Inhibierung kann man sehen, dass durch den Inhibitor sowohl die maximale Reaktionsgeschwindigkeit als auch der Km-Wert verändert wird.
Durch Umformung der Geschwindigkeitsgleichung in die reziproke Form kann die Abhängigkeit von Reaktionsgeschwindigkeit zu Ausgangskonzentration des Substrates linearisiert dargestellt werden:
Aus dieser Formel ist zu erkennen, dass der Anstieg unabhängig vom unkompetitiven Inhibitor ist, das heißt die Graphen liegen bei unterschiedlichen Inhibitorkonzentrationen parallel zueinander. Die Nullstelle gibt den Wert i / Km wieder. Die Geraden schneiden die Ordinate im Punkt i / Vmax.
Die unkompetitive Hemmung kommt zum Beispiel bei Oxidasen vor, wenn der Inhibitor nur mit einer bestimmten Oxidationsstufe des Enzyms reagieren kann. Eine weitere Möglichkeit für einen unkompetitiven Inhibitor bietet sich bei einem Ordered-Mechanismus, einer Zwei-Substratreaktion, bei der der Inhibitor konkurrierend zu einem der Substrate auftritt.
Partiell kompetitive Hemmung
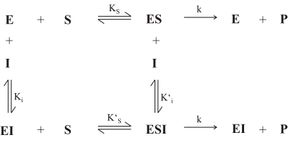 Abb. 11: Allgemeiner Mechanismus der partiell kompetitiven Enzymhemmung (E: Enzym, S: Substrat, P: Produkt, I: Inhibitor, ES: Enzym-Substrat-Komplex, EI: Enzym-Inhibitor-Komplex, ESI: Enzym-Substrat-Inhibitor-Komplex)
Abb. 11: Allgemeiner Mechanismus der partiell kompetitiven Enzymhemmung (E: Enzym, S: Substrat, P: Produkt, I: Inhibitor, ES: Enzym-Substrat-Komplex, EI: Enzym-Inhibitor-Komplex, ESI: Enzym-Substrat-Inhibitor-Komplex)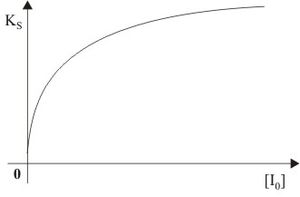 Abb. 12: Abhängigkeit des Ks-Wertes von der Ausgangsinhibitorkonzentration([I0]) einer partiell kompetitiven Hemmung
Abb. 12: Abhängigkeit des Ks-Wertes von der Ausgangsinhibitorkonzentration([I0]) einer partiell kompetitiven HemmungMan spricht von einer partiell kompetitiven Hemmung, wenn durch Inhibitorbindung (I) lediglich die Affinität des Enzyms E zum Substrat S herabgesetzt wird, ohne dass die Geschwindigkeitskonstante zur Produktbildung (P) beeinflusst wird. Das bedeutet, dass auch nach Bindung des Inhibitors Produkt gebildet werden kann. In Abb. 11 ist der Mechanismus der partiell kompetitiven Hemmung dargestellt.
Für einen unter diesen Bedingungen:
- rapid equilibrium (Bindung des Substrats bzw. Inhibitors sehr viel schneller als Bildung des Produkts)
- hohe Inhibitorkonzentrationen
erfolgten partiell kompetitiven Hemmtyp kann die Reaktionsgeschwindigkeit so berechnet werden:
Aus dieser Geschwindigkeitsgleichung ist zu sehen, dass der Ks-Wert sich entgegen der kompetitiven Hemmung dem Wert K's annähert. Im Lineweaver-Burk-Diagramm ist die partiell kompetitive Hemmung nicht von der kompetitiven zu unterscheiden. Um diese aber dennoch zu differenzieren bedient man sich der Abhängigkeit des Ks-Wertes von der Inhibitorkonzentration, die in Abb. 12 dargestellt ist. Bei der kompetitiven Inhibierung stellt diese Abhängigkeit eine Gerade im Diagramm dar. Wie in der Abbildung zu erkennen ist, trifft dieser Sachverhalt nicht für die partiell kompetitive Hemmung zu.
Substratüberschusshemmung
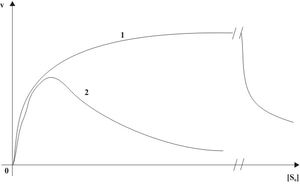 Abb. 13: Michaelis-Menten-Auftragung einer Substratüberschusshemmung (1: Ki=Km, 2: Ki>>Km)
Abb. 13: Michaelis-Menten-Auftragung einer Substratüberschusshemmung (1: Ki=Km, 2: Ki>>Km)Durch sehr hohe Substratkonzentrationen kann es bei manchen Enzymen zur Bindung eines zweiten Substratmoleküls an das Enzym kommen. Der so entstandene ESS-Komplex ist nicht in der Lage in Produkt und Enzym zu zerfallen. Für diesen Reaktionsmechanismus lautet die Geschwindigkeitsgleichung so:
Die Dissoziationskonstante des ESS-Komplexes wurde als Ki bezeichnet. Wenn der Km-Wert sehr viel niedriger als der Ki-Wert ist, erhält man eine hyperbole Abhängigkeit in der Michaelis-Menten-Auftragung (Abb. 13, Kurve 1). Sind diese beiden Werte annähernd gleich oder der Km-Wert höher, so entsteht sozusagen eine Optimumkurve (Abb. 13, Kurve 2).
Hemmung durch Reaktion eines Inhibitors mit dem Substrat
Bei diesem Hemmtyp reagiert der Inhibitor mit dem Substrat, welches dann von dem Enzym nicht mehr umgesetzt wird. Die Bindung des Inhibitors wird bei dieser Hemmung als reversibel angesehen. Dadurch wird die dem Enzym frei zugängliche Substratkonzentration [Seff] herabgesetzt:
- [Seff] = [S0] − [SI]
In Gegenwart des Inhibitors wird die Maximalgeschwindigkeit bei hohen Substratkonzentrationen erreicht. Im Lineweaver-Burk-Diagramm lässt sich dieser Mechanismus von einem kompetitiven Inhibitor differenzieren, da eine Abweichung von der Linearität auftritt.
Irreversible Enzymhemmung
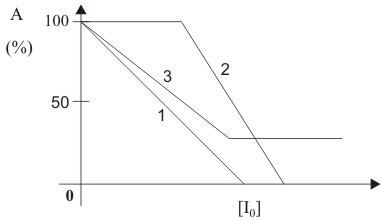
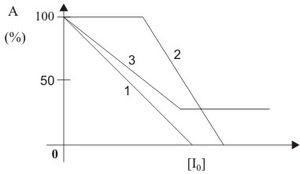 Abb. 14: Abhängigkeit der Aktivität von der Inhibitorkonzentration einer irreversiblen Enzymhemmung (im Text erläutert)
Abb. 14: Abhängigkeit der Aktivität von der Inhibitorkonzentration einer irreversiblen Enzymhemmung (im Text erläutert)Durch die irreversible Bindung des Inhibitors an das Enzym wird die katalytische Aktivität vermindert. Eine Dissoziation des Enzym-Inhibitor-Komplexes in freies Enzym und Inhibitor ist nicht möglich, d. h. das Enzym bleibt für immer inaktiv. Die Aktivität hängt linear von der Inhibitorkonzentration ab. Diese Abhängigkeit ist in Abb. 14 (Kurve 1) zu erkennen. Abweichend von diesem Verlauf entsteht die Kurve 2 in der Abbildung. Dabei reagiert der Inhibitor irreversibel mit mehreren Gruppen unterschiedlicher Spezifität, was zum Austitrieren der spezifischeren, aber nicht aktiv am Katalysemechanismus beteiligten Gruppen, führt. Anschließend reagieren die für die Katalyse bedeutsamen Gruppen, wodurch die Aktivität gesenkt wird. Andererseits kann der Inhibitor mit dem Enzym einen Komplex eingehen, der noch geringe Aktivität aufweist. Dies ist bei der Kurve 3 der Abbildung der Fall.
Ist die Inhibitorkonzentration erheblich höher als die des Enzyms, so kann man die Geschwindigkeitskonstante für die Reaktion des Enzyms mit dem Inhibitor als Reaktion pseudo-erster Ordnung formulieren:
A entspricht in dieser Gleichung der Aktivität. Die Geschwindigkeitskonstante k' kann aus dem Anstieg bestimmt werden, indem man den Logarithmus der Aktivität gegen die Reaktionszeit aufträgt.
Die Geschwindigkeitskonstante wird durch die Anwesenheit des Substrates beeinflusst. Denn dieses schützt das Enzym vor der Inaktivierung durch den Inhibitor, wodurch die Geschwindigkeitskonstante kleiner ist als ohne Substratzugabe. Bei einem solchen Verhalten kann es sein, dass der Inhibitor an eine spezifische Gruppe im aktiven Zentrum bindet, also eine gleiche Bindungsstelle wie das Substrat aufweist.
Ein Beispiel für eine irreversible Hemmung sind die sogenannten „Suizid-Substrate“, die mit der funktionellen Gruppe des Enzyms eine kovalente Bindung eingehen und dieses somit blockieren.
Allosterische Hemmung
Bei der allosterischen Hemmung (griech.: allos: anders; steros: Ort) lagern sich die Hemmstoffe, auch allosterische Effektoren genannt, nicht (wie bei der kompetitiven Hemmung) an das aktive Zentrum, sondern an einer anderen Stelle des Enzyms (das allosterische Zentrum) an. Dabei wird die Konformation des Enzyms so verändert, dass die Bindung des Substrats am aktiven Zentrum erschwert bzw. ganz unmöglich gemacht wird.
Die allosterische Hemmung lässt sich nur durch die Entfernung des Inhibitors rückgängig machen. Ein Enzym, welches die erste Reaktion einer Reaktionskette katalysiert, wird oft durch die am Ende gebildete Substanz gehemmt (dieser Fall wird dann Endprodukthemmung/Feedback-Hemmung genannt.)
Siehe auch
Enzymkinetik, Michaelis-Menten-Theorie, Katalytische Effizienz
Weblinks
- Programme (Composit, MMenten) zur Analyse von Inhibitionstypen
- Seite zur Enzymhemmung
- Sächsischer Bildungsserver - Enzyme
- Uni Düsseldorf - Allosterische Hemmung
- Doccheck medical services GmbH - Enzym
Literatur
- Hans Bisswanger: Enzymkinetik. Theorie und Methoden Wiley-VCH, 2000, ISBN 3-52-730096-1
- Alfred Schellenberger (Hrsg.): Enzymkatalyse: Einführung in die Chemie, Biochemie und Technologie der Enzyme Gustav Fischer Verlag, Jena 1989
![\frac{d[P]}{dt} = k_2[ES] + k_6[ESI] = v](/pictures/dewiki/53/5e3aafb2910dea3559caea2f3a1bc7be.png)
![v = \frac{\left(V_1 + \frac{V_2[I]}{K_{iu}}\right)[S]}{K_m\;\left(1 + \frac{[I]}{K_{ic}}\right) + \left(1 + \frac{[I]}{K_{iu}}\right)\;[S]}](/pictures/dewiki/52/40dcad615372dfbe3798bb7f4c33a2ac.png)

![v = \frac{V_{\max}\;[S_0]}{[S_0] + K_m \cdot i}](/pictures/dewiki/54/608c46c648cf06f17c6bd218732c653b.png)
![\frac{1}{v} = \frac{1}{V_{\max}} + \frac{k_m \cdot i}{V_{\max}}\;\frac{1}{[S_0]}](/pictures/dewiki/98/bf05df05d8c775bc55e02033c359a3c6.png)
![v_A = \frac{V_{\max A}\;[A_0]}{[A_0] + K_{mA} \cdot i_B} \qquad v_B = \frac{V_{\max B} [B_0]}{[B_0] + K_{mB} \cdot i_A}](/pictures/dewiki/56/8a18adfaba35080aab13eeb66fd75e02.png)
![i_B = \left(1+\frac{[B_0]}{K_{mB}}\right) \qquad\qquad i_A=\left(1+\frac{[A_0]}{K_{mA}}\right)](/pictures/dewiki/99/cf27ebd01646b18d0e53b5f2647cb17d.png)

![v = \frac{V_{\max}\;[S_0]}{i \cdot ([S_0] + K_m)}](/pictures/dewiki/99/c667a4e8afa515786a00492dc1452029.png)
![\frac{1}{v} = \frac{i}{V_{\max}} + \frac{K_m \cdot i}{V_{\max}}\;\frac{1}{[S_0]}](/pictures/dewiki/99/ceec8c6ebcf3b96a23582aa997b2f7fe.png)


![v = \frac{V_{\max}\;[S_0]}{i \cdot \left(\frac{K_m}{i} + [S_0]\right)}](/pictures/dewiki/102/f6487aa150ef8b30c37c39e542605bb0.png)
![\frac{1}{v} = \frac{i}{V_{\max}} + \frac{K_m}{V_{\max}}\;\frac{1}{[S_0]}](/pictures/dewiki/50/25a36910b004d18ee2a1a93932b3e321.png)

![v = \frac{V_{\max}\;[S_0]}{[S_0] + K'_s}](/pictures/dewiki/100/de484c72377f4346b633c03082be7219.png)

![v = \frac{V_{\max}}{1 + \frac{K_m}{[S_0]} + \frac{[S_0]}{K_i}}](/pictures/dewiki/50/2162f9de87c8ae053f27522b38c42d39.png)
![-\frac{d[E]}{dt} = -\frac{dA}{dt} = k\;[I_0]\;[E] = k'\;[E]](/pictures/dewiki/50/23d7ac4211f1267f400b620e3cb5cdc9.png)
Wikimedia Foundation.