- B-Zelle
-
B-Lymphozyten oder kurz B-Zellen gehören zu den Leukozyten (weißen Blutkörperchen). Sie sind als einzige Zellen in der Lage Antikörper zu bilden und machen zusammen mit den T-Lymphozyten den entscheidenden Bestandteil des adaptiven Immunsystems aus. Während T-Zellen an der zellvermittelten Immunantwort beteiligt sind, sind die B-Zellen die Träger der humoralen Immunantwort (Bildung von Antikörpern). Wenn sie durch körperfremde Antigene aktiviert werden, können sie sich zu Antikörper-produzierenden Plasmazellen oder zu Gedächtniszellen differenzieren. Die Bezeichnung „B-Zellen“ stammt ursprünglich von ihrem Bildungsort in der Bursa Fabricii bei Vögeln. Beim Menschen und einigen anderen Säugetieren entstehen die B-Zellen im Knochenmark, daher erhielt der Buchstabe B hier nachträglich die Bedeutung bone marrow (engl. für Knochenmark), für viele Säugetiere ist die Herkunft noch nicht eindeutig geklärt, hier verwendet man den Begriff „bursaäquivalentes Organ“.
Inhaltsverzeichnis
Entwicklung
Die Entwicklung der B-Zellen findet beim Menschen und auch bei einigen anderen Säugern im Knochenmark beziehungsweise der fetalen Leber statt. Die Signale, die für das Entwicklungsprogramm notwendig sind, erhalten die sich entwickelnden Lymphozyten von so genannten Stromazellen. Bei der B-Zellentwicklung ist die Bildung eines funktionierenden B-Zell-Rezeptors (die membrangebundene Form des Antikörpers) von entscheidender Bedeutung. Nur mit diesem Antigenrezeptor sind reife B-Zellen später in der Lage, fremde Antigene zu erkennen und durch die Bildung von entsprechenden Antikörpern feindliche Strukturen zu bekämpfen. Die Antigenspezifität des Rezeptors wird durch die Verknüpfung bestimmter Gensegmente bestimmt. Die Segmente heißen V-, D- und J-Segmente, weshalb der Prozess als V(D)J-Rekombination bezeichnet wird. Dabei werden diese Segmente, die den Antigenbindenden Teil des B-Zell-Rezeptors bilden, umgeordnet. Der gesamte Rezeptor besteht aus zwei identischen leichten Proteinketten und zwei identischen schweren Proteinketten, die über Disulfidbindungen verknüpft sind. Bei der VDJ-Rekombination werden zuerst die V-, D- und J-Segmente der schweren Kette des B-Zell-Rezeptors verknüpft, danach die V- und J-Segmente der leichten Rezeptorkette. Nur wenn die Gene dabei erfolgreich umgeordnet werden, was als produktive Genumordnung bezeichnet wird, kann die Zelle in den jeweils nächsten Entwicklungsschritt übergehen.
Stadien der B-Zell-Entwicklung
Das erste Stadium der B-Zellentwicklung stellen die Pro-B-Zellen dar, die sich von pluripotenten Stammzellen herleiten. In Pro-B-Zellen erfolgt die Umordnung der schweren Kette. Im Falle einer produktiven Genumlagerung wird dabei eine so genannte schwere μ-Kette gebildet, was zum Eintritt in das Stadium der Prä-B-Zelle führt. Im ersten Prä-B-Zell-Stadium, den so genannten großen Prä-B-Zellen, wird die schwere μ-Kette zusammen mit einem Ersatz für die leichte Kette in Form eines Prä-B-Zell-Rezeptors auf der Zelloberfläche exprimiert. Die großen Prä-B-Zellen teilen sich mehrmals und entwickeln sich schließlich zu den kleinen Prä-B-Zellen, die keinen Prä-B-Zell-Rezeptor mehr bilden und nur noch über intrazelluläre schwere μ-Ketten verfügen. In den kleinen Prä-B-Zellen wird mit der V-J-Umordnung der leichten Kette begonnen. Nachdem die Gene für die leichte Kette erfolgreich umgelagert wurden, geht die Zelle ins Stadium einer unreifen B-Zelle über. Dabei wird ein komplettes B-Zell-Rezeptormolekül IgM auf der Oberfläche exprimiert. Reife B-Zellen zeichnen sich schließlich dadurch aus, dass bei ihnen durch alternatives Spleißen auch IgD-Rezeptoren gebildet werden.
- frühe Pro-B-Zelle: D-J-Umordnung der schweren Kette
- späte Pro-B-Zelle: V-DJ-Umordnung der schweren Kette
- große Prä-B-Zelle: μ-Kette als Teil des Prä-B-Zell-Rezeptors auf der Oberfläche
- kleine Prä-B-Zelle: V-J-Umordnung der leichten Kette
- unreife B-Zelle: IgM-Rezeptor auf der Oberfläche
- reife B-Zelle: IgM und IgD Rezeptoren auf der Oberfläche.
Funktion
 Plasmazelle
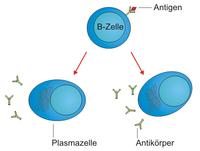
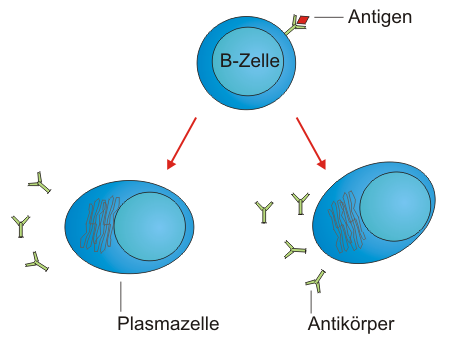
PlasmazelleB-Zellen sind in der Lage, mit ihren B-Zell-Rezeptoren bestimmte, in der Regel körperfremde Strukturen – sogenannte Antigene – zu erkennen und daraufhin gegen diese Antigene gerichtete Antikörper zu produzieren. Diese B-Zellen beginnen jedoch erst dann mit der Produktion von Antikörpern, wenn sie zuvor vollständig aktiviert wurden. Naive B-Lymphozyten (reife B-Zellen, die noch keinen Kontakt zu ihrem Antigen hatten) zirkulieren im Blut und den lymphatischen Organen (Thymus, Milz, Lymphknoten, Knochenmark) des Körpers von Wirbeltieren. Sobald eine B-Zelle mit ihrem B-Zell-Rezeptor an ein fremdes Antigen bindet und gleichzeitig ein costimulatorisches Signal von T-Helferzellen (die ebenfalls dasselbe Antigen erkannt haben müssen) bekommt, wandert sie zu den so genannten Keimzentren in Lymphknoten oder Milz. Hier teilt sie sich stark (Proliferation), differenziert zur sogenannten Plasmazelle und sezerniert anschließend Antikörper. Diese Antikörper besitzen dieselbe Spezifität wie der B-Zell-Rezeptor der Zelle, das heißt, sie binden an dasselbe Antigen. Zusätzlich werden in ihre Antikörper-Gene Mutationen eingeführt, die zur Verbesserung der Antikörper-Affinität für das erkannte Antigen führen können (somatische Hypermutation). Außerdem kann hierbei ein Klassenwechsel des konstanten (konservierten) Teils der Antikörper, der die Funktion (z. B. als Membranrezeptor) determiniert, stattfinden. Das wiederum ist wichtig für die Art, wie die Antikörper im Weiteren auf den Erreger wirken beziehungsweise wohin die Antikörper im Körper gelangen.
Beim Menschen gibt es rund 109 bis 1010 verschiedene, spezifische B-Lymphozyten, die sich in ihren Antigen-Rezeptoren unterscheiden und durch V(D)J-Rekombination entstehen.
B-Lymphozyten tragen auf ihrer Oberfläche eine Reihe von Eiweißen (die so genannten Oberflächenmarker), die funktionell wichtig sind und zu ihrer Identifizierung z. B. im menschlichen Blut oder in Gewebeproben verwendet werden können. Neben den membranständigen Immunglobulinen (Antikörpern) zählen dazu z. B. CD19, CD20 und CD21.
Antigenerkennung durch B-Zellen
Ein grundlegender Unterschied zwischen B- und T-Zellen liegt in der Form, in der sie ihr entsprechendes Antigen erkennen. B-Zellen binden in ungebundener (löslicher) Form vorliegende Antigene direkt mit Hilfe ihres membrangebundenen B-Zell-Rezeptors. T-Zellen hingegen binden mit Hilfe ihres T-Zell-Rezeptors lediglich Peptide-Fragmente des Antigens, nachdem diese (nach entsprechender Prozessierung des Antigens) auf der Oberfläche von Antigenpräsentierende Zellen an deren MHC-Molekül als Antigen-MHC-Komplex präsentiert werden.
Aktivierung von B-Zellen
Die Erkennung des Antigens durch die B-Zelle ist nicht unbedingt das einzige notwendige Element, das für ihre Aktivierung erforderlich ist. Naive B-Zellen, also solche, die noch keinen Kontakt zu ihrem Antigen hatten, benötigen oft eine zusätzliche Stimulierung durch T-Zellen um aktiviert zu werden. Je nach Art des Antigens können B-Zellen in einer T-Zell-abhängigen oder T-Zell-unabhängigen Weise aktiviert werden.
T-Zell-abhängige Aktivierung
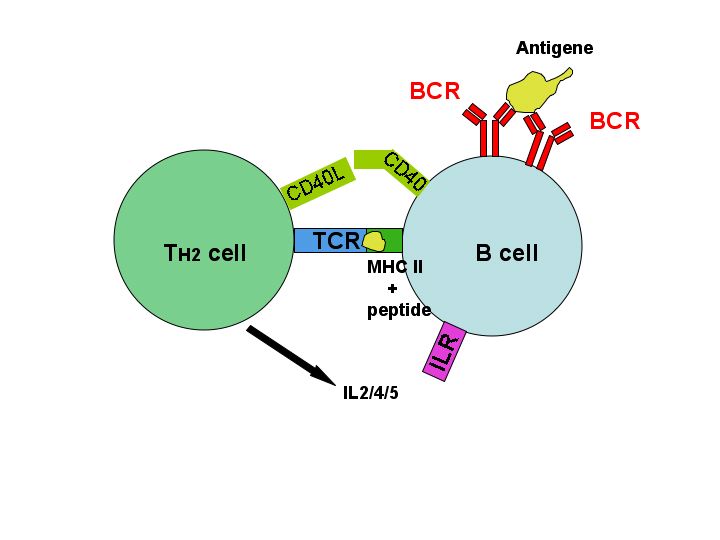
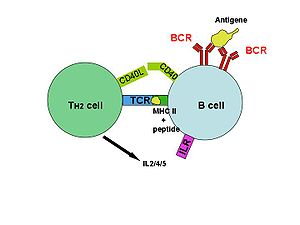 B-Zelle wird durch T-Helferzelle aktiviert nachdem sie ihr das internalisierte Antigen als Peptid zusammen mit dem MHC-II-Komplex präsentiert hat. Weitere Interaktionen erfolgen durch CD40 und dessen Ligand (CD40L), sowie durch Interleukine (IL 2/4/5), die die T-Zelle ausschüttet.
B-Zelle wird durch T-Helferzelle aktiviert nachdem sie ihr das internalisierte Antigen als Peptid zusammen mit dem MHC-II-Komplex präsentiert hat. Weitere Interaktionen erfolgen durch CD40 und dessen Ligand (CD40L), sowie durch Interleukine (IL 2/4/5), die die T-Zelle ausschüttet.Die meisten Antigene sind T-Zell-abhängig, das heißt für eine maximale Antikörperproduktion ist eine Beteiligung von T-Zellen erforderlich. Zwei unterschiedliche Signale sind hierbei nötig, um die B-Zelle zu aktivieren: Das erste entsteht durch die Kreuzvernetzung des Antigenrezeptors auf der Oberfläche der Zelle, nachdem dieser ein entsprechendes Antigen gebunden hat. Das zweite erhält die B-Zelle von einer sogenannten T-Helferzelle (TH2-Zelle). Dabei wird das Antigen – nach der Bindung durch den B-Zell-Rezeptor – ins Zellinnere der B-Zelle aufgenommen und zusammen mit einem MHC-Molekül an der Oberfläche präsentiert. Hier kann eine entsprechende T-Zelle (T-H2-Helferzelle) mit Hilfe ihres T-Zell Rezeptors an den Antigen-MHC-Komplex binden. Die T-Zelle aktiviert nun die B-Zelle indem sie bestimmte Zytokine ausschüttet. Daraufhin vermehrt sich die B-Zelle (klonale Expansion) und differenziert zu einer Antikörper-produzierenden B-Zelle (Plasmazelle). Der Klassenwechsel der B-Zelle, nach dem nun auch Antikörper der Klassen IgG, IgA und IgE gebildet werden können, sowie die Ausbildung von Gedächtnis-B-Zellen sind T-Zell-abhängige Antworten einer B-Zelle.
T-Zell-unabhängige Aktivierung
Einige Antigene sind T-Zell-unabhängig, sie benötigen also nur ein einziges Signal, das durch Kreuzvernetzung der B-Zell-Rezeptoren erzeugt wird. Vor allem sich wiederholende Polysaccharide, wie sie an der Oberfläche von Bakterien vorkommen, können auf diese Weise erkannt werden. Die B-Zelle wird dadurch aktiviert, vermehrt sich und bildet Antikörper der Klasse IgM. Ein Klassenwechsel unterbleibt bei dieser Form der Aktivierung ebenso wie die Bildung von Gedächtniszellen. Aus diesem Grunde führt die Impfung mit Polysaccharid-Impfstoffen in der Regel nur zu einem zeitlich befristeten Schutz von 3 bis 6 Jahren.
B-Zell-Typen
Neben den verschiedenen Reife- und Endstadien von B-Zellen gibt es zwei grundlegend unterschiedliche Typen von B-Zellen.
- B2-Zellen: Sie stellen das Gros der B-Zellen, man könnte sie als die „gewöhnlichen“ B-Zellen bezeichnen.
- B1-Zellen: B1-Zellen sind größer als B2-Zellen und kommen vor allem in der Bauchhöhle vor. In der Milz machen sie insgesamt nur etwa 5 % der B-Zellen aus, in peripheren Lymphknoten fehlen sie. Sie reagieren relativ schwach auf Proteinantigene, dafür besser auf Kohlenhydratantigene und zeigen im Vergleich zu B2-Zellen weniger somatische Hypermutation (siehe unter Funktion) und weniger Klassenwechsel. B1-Zellen unterscheiden sich durch bestimmte Oberflächenmarker von B2-Zellen, so tragen sie im Gegensatz zu B2-Zellen weniger IgD, mehr IgM, kein CD23 aber dafür CD43. Ursprünglich wurden B1-Zellen von B2-Zellen durch die Anwesenheit des T-Zell-Oberflächenmarkers CD5 unterschieden. Mittlerweile wurde unter den B-Zellen der Bauchhöhle allerdings eine Subpopulation entdeckt, die zwar kein CD5 trägt, aber in anderen Oberflächenmerkmalen identisch mit B1-Zellen ist. Daher wurden die CD5 positiven B1-Zellen mit dem Namen B1a-Zellen, die CD5 negativen mit dem Namen B1b-Zellen belegt. Ob B1a-Zellen einer separaten Linie entspringen und ihren Ursprung im Fötus haben oder ob sie sich aus gewöhnlichen B2-Zellen entwickeln können, ist noch umstritten. Die genannten Merkmale beziehen sich in erster Linie auf Mäuse, die diesbezüglich am besten untersucht sind. Im Menschen existieren zwei Klassen von CD5 positiven B-Zellen. Eine davon gleicht auch in anderen Aspekten den B1a-Zellen von Mäusen. In Ratten sind dagegen keine CD5 positiven B-Zellen bekannt. In Hasen, Rindern und Hühnern tragen die meisten peripheren B-Zellen CD5.[1]
- Marginalzonen B-Zellen: Marginalzonen B-Zellen (MZ B-Zellen) findet man in der Marginalzone der Milz, sie machen nur etwa 5 % der B-Zellen einer Milz aus, sind aber ein wichtiger Bestandteil der frühen Immunantwort gegen Krankheitserreger des Blutes.[2]
Siehe auch
Literatur
- Charles A. Janeway jr. u. a.: Immunologie. 5. Auflage. Spektrum Akademischer Verlag Gmbh, Heidelberg, Berlin 2002, ISBN 3-8274-1078-9.
- Janis Kuby et al.: Immunology. 5. Auflage. W. H. Freeman and Company, New York 2003, ISBN 0-7167-4947-5.
Einzelnachweise
- ↑ Robert Berland, Henry H. Wortis: Origins and Functions of B-1 Cells With Notes on the Role of CD5. In: Annual Review of Immunology. Vol. 20: 253-300 (Veröffentlichungsdatum April 2002).
- ↑ Thiago Lopes-Carvalho, John F. Kearney: Development and selection of marginal zone B cells. In: Immunological Reviews. Volume 197 Issue 1, Page 192-205, February 2004.
Wikimedia Foundation.