- Selaginella
-
Moosfarne 
oben: Dorniger Moosfarn (Selaginella selaginoides)
unten: Schweizer Moosfarn (Selaginella helvetica)Systematik Unterreich: Gefäßpflanzen (Tracheobionta) Überabteilung: Gefäßsporenpflanzen (Pteridophyta) Abteilung: Bärlapppflanzen (Lycopodiophyta) Ordnung: Moosfarnartige Familie: Moosfarngewächse Gattung: Moosfarne Wissenschaftlicher Name der Ordnung Selaginellales Wettst. Wissenschaftlicher Name der Familie Selaginellaceae Rchb. Wissenschaftlicher Name der Gattung Selaginella P.Beauv. Die Moosfarne (Selaginella) sind eine Pflanzengattung, die zu den Bärlapppflanzen (Lycopodiophyta) gehört. Moosfarne sind die einzige Gattung der Familie der Moosfarngewächse (Selaginellaceae) und der Ordnung der Moosfarnartigen (Selaginellales). Die Gattung umfasst weltweit etwa 700 Arten. Es sind meist kleine, krautige Pflanzen, die sich durch ihre Heterosporie, das heißt verschiedenartige Sporen, auszeichnen. Der Verbreitungsschwerpunkt sind die Tropen, mit relativ wenigen Arten in den gemäßigten Zonen.
Inhaltsverzeichnis
Merkmale
Die Moosfarne ähneln in ihrem Habitus ein wenig manchen Moosen, was sich im ersten Teil des Gattungsnamens niederschlägt. In ihren anatomischen Merkmalen und ihrer Fortpflanzung sind sie jedoch eindeutig Gefäßsporenpflanzen.
Sprossachse
Sie besitzen meist niederliegende oder aufrechte, reich gabelig (dichotom) verzweigte Sprossachsen. Einige Arten sind rasenbildend. Wenige Arten klettern an Sträuchern empor und werden mehrere Meter hoch.
An den Gabelungsstellen des Sprosses sitzen häufig zylindrische, gestreckte, nach unten gerichtete farb- und blattlose Sprosse, die Wurzelträger oder Rhizophoren. Als Sprossachsen entstehen diese exogen (an der Oberfläche) im Gegensatz zu den endogen (im Inneren) entstehenden Wurzeln, und verzweigen sich ebenfalls gabelig. An ihren Enden stehen Wurzelbüschel.
Die Leitbündel der Sprossachse können eine zentrale Protostele bilden, aber auch eine Distele oder Siphonostele. Ein sekundäres Dickenwachstum fehlt. Sehr selten kommen bereits echte Tracheen vor, in denen die Querwände zwischen den Zellen aufgelöst sind, während die Seitenwände die charakteristischen treppenförmigen Verdickungen aufweisen. Die Endodermis einiger Arten (etwa S. kraussiana) wird von röhrenförmigen Zellen mit Casparyschen Streifen gebildet (Trabeculae).
Blätter
 Sporophyllstand von Selaginella sp.
Sporophyllstand von Selaginella sp.Die Blätter sind klein und schuppenartig (Mikrophylle). Sie sitzen schraubig oder – häufiger – gegenständig in vier Zeilen an der Sprossachse. Meist gibt es zwei Reihen von kleinen Oberblättern und zwei Reihen von großen Unterblättern (Anisophyllie). Bei niederliegenden Formen weist bei den Oberblättern die Unterseite nach oben, bei den Unterblättern die Oberseite. Die Blätter haben eine unverzweigte Mittelrippe. Das Mesophyll ist nur selten in Schwamm- und Palisadenparenchym differenziert, in der Regel jedoch dem Schwammparenchym ähnlich. Die Epidermiszellen besitzen Chloroplasten, häufig nur einen schüsselförmigen pro Zelle. Auch im Mesophyll besitzen die Zellen mancher Arten nur einen schüsselförmigen Chloroplasten. In der Blattachsel, am Grund der Blattoberfläche sitzt eine kleine, häutige, chlorophyllfreie Schuppe (Ligula). Sie besitzt bei einigen Arten einen direkten Anschluss an die Tracheiden der Sprossachse und dient der raschen Wasseraufnahme nach Regenfällen.
Sporophyllstände
Die Sporophyllstände sind endständig. Sie sind einfach oder verzweigt, vierkantig radiär oder dorsiventral. An jedem Sporophyll steht nur ein einzelnes Sporangium, das an der Blattachsel entspringt. Die Sporangien bilden große (weibliche) Megasporen oder kleine (männliche) Mikrosporen, die Moosfarne sind also heterospor. Beide Sorten kommen an ein und demselben Sporophyllstand vor, die Geschlechtsbestimmung erfolgt bereits in der Diplophase (diplomodifikatorische Geschlechtsbestimmung). Meist befinden sich die Megasporangien am unteren Teil des Sporophyllstandes.
Im Megasporangium entwickelt sich nur eine Megasporenmutterzelle weiter, die anderen gehen zugrunde. Aus ihr entwickeln sich durch Meiose vier Megasporen mit buckliger Wand. Bei einigen Arten ist die Zahl reduziert, so bildet S. rupestris nur eine Megaspore pro Sporangium; bei anderen ist die Anzahl vermehrt, S. willdenowii bildet bis zu 42. Die Sporangienwand öffnet sich bei Reife mit Klappen.
In den flach gedrückten Mikrosporangien entwickeln sich viele Mikrosporen, die nach dem Öffnen mit zwei Klappen ausgestreut werden.
Die Sporangienwand ist aus drei Zellschichten zusammengesetzt, wobei die mittlere im reifen Sporangium sehr schmal ist. Die innerste ist die Tapetenschicht, die der Ernährung der sich entwickelnden Sporen dient, jedoch erhalten bleibt und sich nicht auflöst, also ein Sekretionstapetum ist. Die Öffnung der Sporangien erfolgt durch einen Kohäsionsmechanismus entlang einer vorbezeichneten Linie. Dabei werden die Sporen ausgeschleudert. Bei einigen Arten verbleiben jedoch die Megasporen an der Mutterpflanze (S. rupestris), sodass die Embryos an der Mutterpflanze keimen.
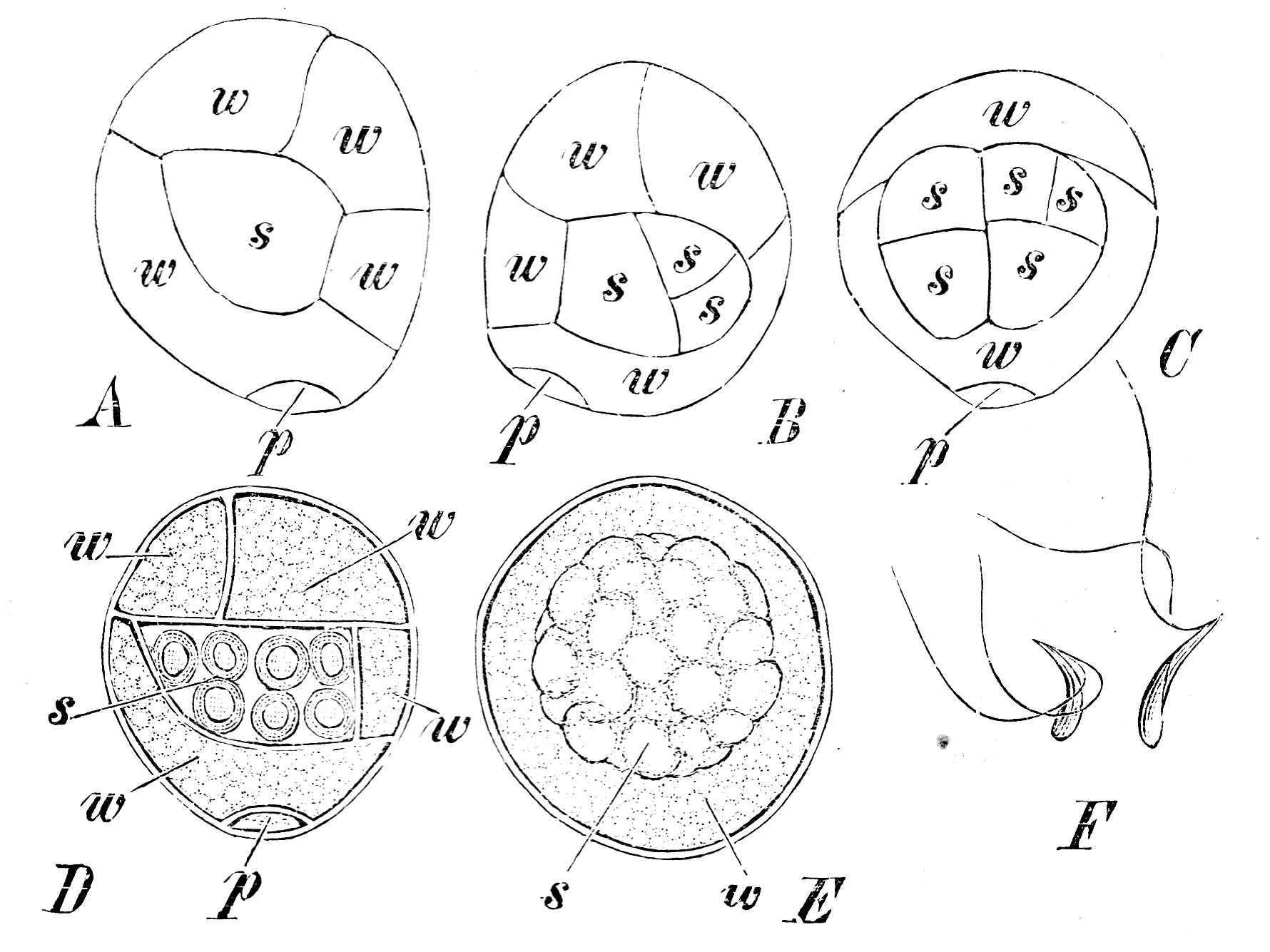
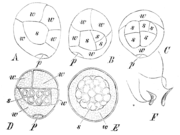 S. stolonifera (A-E): Keimung der Mikrosporen. p Prothalliumzelle, w Antheridiumwandzelle, s spermatogene Zelle. A,B,D von der Seite, C vom Rücken. In E ist die Prothalliumzelle nicht sichtbar, die Wandzellen aufgelöst, umgeben die Spermatozoidmutterzellen. F S. cuspidata, Spermatozoiden.
S. stolonifera (A-E): Keimung der Mikrosporen. p Prothalliumzelle, w Antheridiumwandzelle, s spermatogene Zelle. A,B,D von der Seite, C vom Rücken. In E ist die Prothalliumzelle nicht sichtbar, die Wandzellen aufgelöst, umgeben die Spermatozoidmutterzellen. F S. cuspidata, Spermatozoiden.Gametophyten
Die Gametophyten sind stark reduziert und verlassen die Sporenwand nicht beziehungsweise kaum, sie sind also endospor.
Die Mikrosporen entwickeln sich meist bereits im Sporangium weiter. Die Sporenzelle teilt sich in eine kleine, linsenförmige Zelle, die alleine das Prothallium vorstellt und meist als Rhizoide gedeutet wird, sowie in eine große Zelle, aus der das einzige Antheridium entsteht. Sie bildet acht sterile Wandzellen und zwei oder vier zentrale spermatogene Zellen. Letztere teilen sich noch mehrmals und bilden die sich abrundenden Spermatozoidmutterzellen. Die Wandzellen lösen sich auf und bilden eine Schleimschicht, in deren Mitte sich die Spermatozoidmutterzellen befinden. Die Prothalliumzelle bleibt allerdings erhalten. Das Ganze ist weiterhin von der Sporenwand umgeben. Bricht die Mikrosporenwand auf, entlassen auch die Mutterzellen die Spermatozoiden. Diese sind keulenförmig, schwach gekrümmt und besitzen zwei lange Geißeln. Pro Mikrospore werden 128 bis 256 Spermien gebildet.
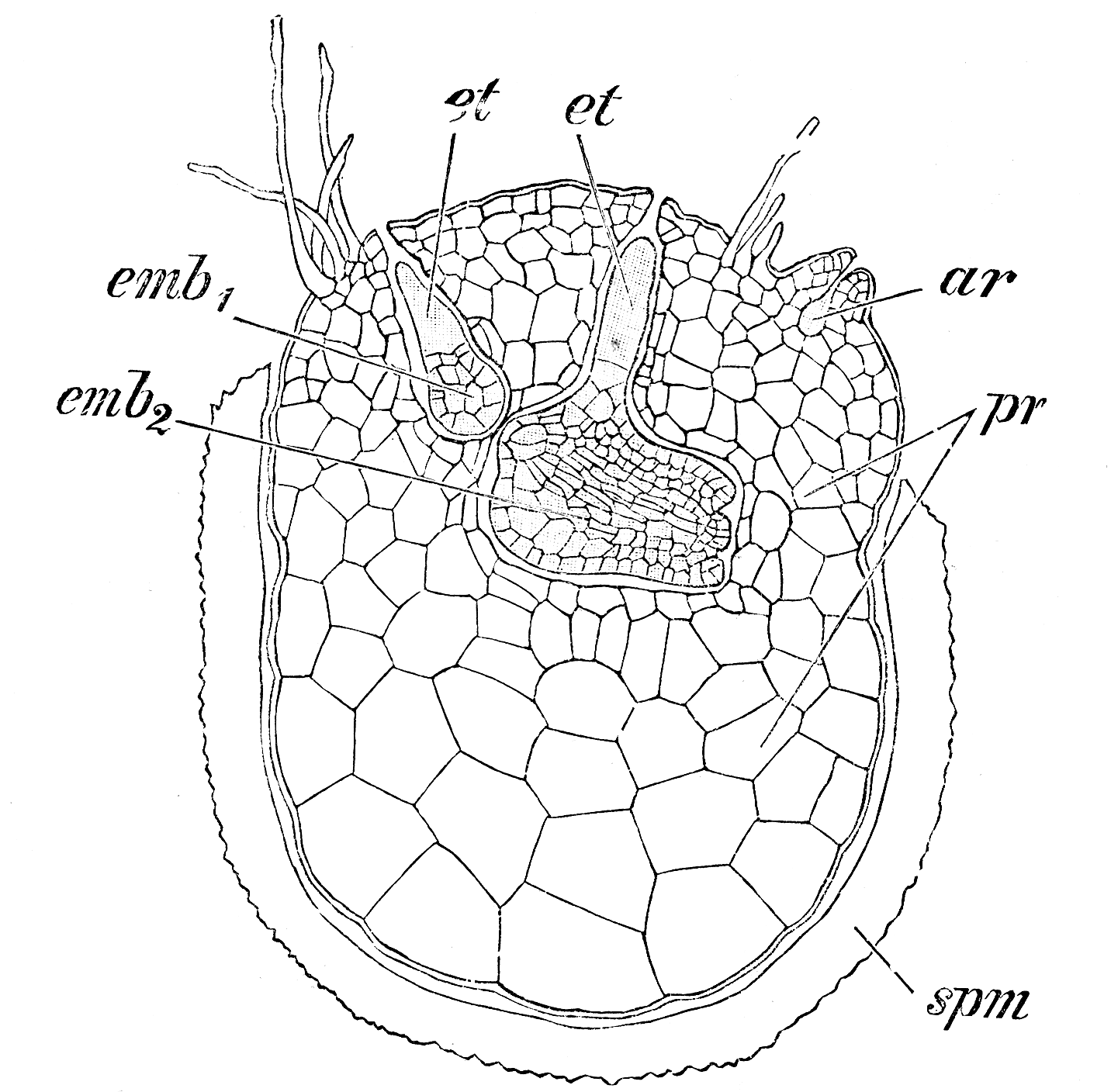
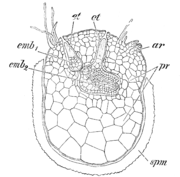 S. martensii. Weibliches Prothallium, aus der am Scheitel geöffneten Megasporenwand spm hervortretend, ar unbefruchtet gebliebenes Archegonium, emb1, emb2 Embryonen mit den Embryoträgern et.
S. martensii. Weibliches Prothallium, aus der am Scheitel geöffneten Megasporenwand spm hervortretend, ar unbefruchtet gebliebenes Archegonium, emb1, emb2 Embryonen mit den Embryoträgern et.Die Megasporen beginnen ihre Entwicklung teilweise auch schon im Sporangium. Die Entwicklung unterscheidet sich dabei teilweise je nach Art. Eine Megaspore hat etwa den zehnfachen Durchmesser einer Mikrospore. Die weiblichen Prothallien sind daher weniger stark reduziert als die männlichen. Der Zellkern befindet sich zunächst am Scheitel der Spore. Er teilt sich durch freie Kernteilungen in viele Tochterkerne, die sich dem Wandplasma entlang nach unten verteilen. Nach oder noch während dieses Vorgangs beginnt vom Scheitel her die Bildung von Zellwänden. Es entstehen zunächst große Prothallienzellen, die sich weiter in kleine Zellen teilen. Am Scheitel werden dann einige wenige Archegonien gebildet. Die Bildung der Archegonien findet meist erst statt, wenn die Sporen das Sporangium verlassen haben. Die Sporenwand platzt dann am Scheitel entlang der drei Sporenkanten auf und das Prothallium quillt etwas über den Sporenrand hinaus. Das Prothallium ist und bleibt chlorophyllfrei. Das Prothallium bildet drei Höcker, an denen Rhizoiden sitzen. Deren Funktion ist die Aufnahme von Wasser und auch die Verankerung des Prothalliums. Bei manchen Arten (etwa S. galeotti) sind diese Höcker weit aus der Hülle ragende „Arme“. Ein oder zwei Archegonien werden befruchtet. Dazu ist wie bei allen Gefäßsporenpflanzen die Anwesenheit von Wasser nötig, damit die Spermatozoiden zur Eizelle im Archegonium gelangen können.
Embryonalentwicklung
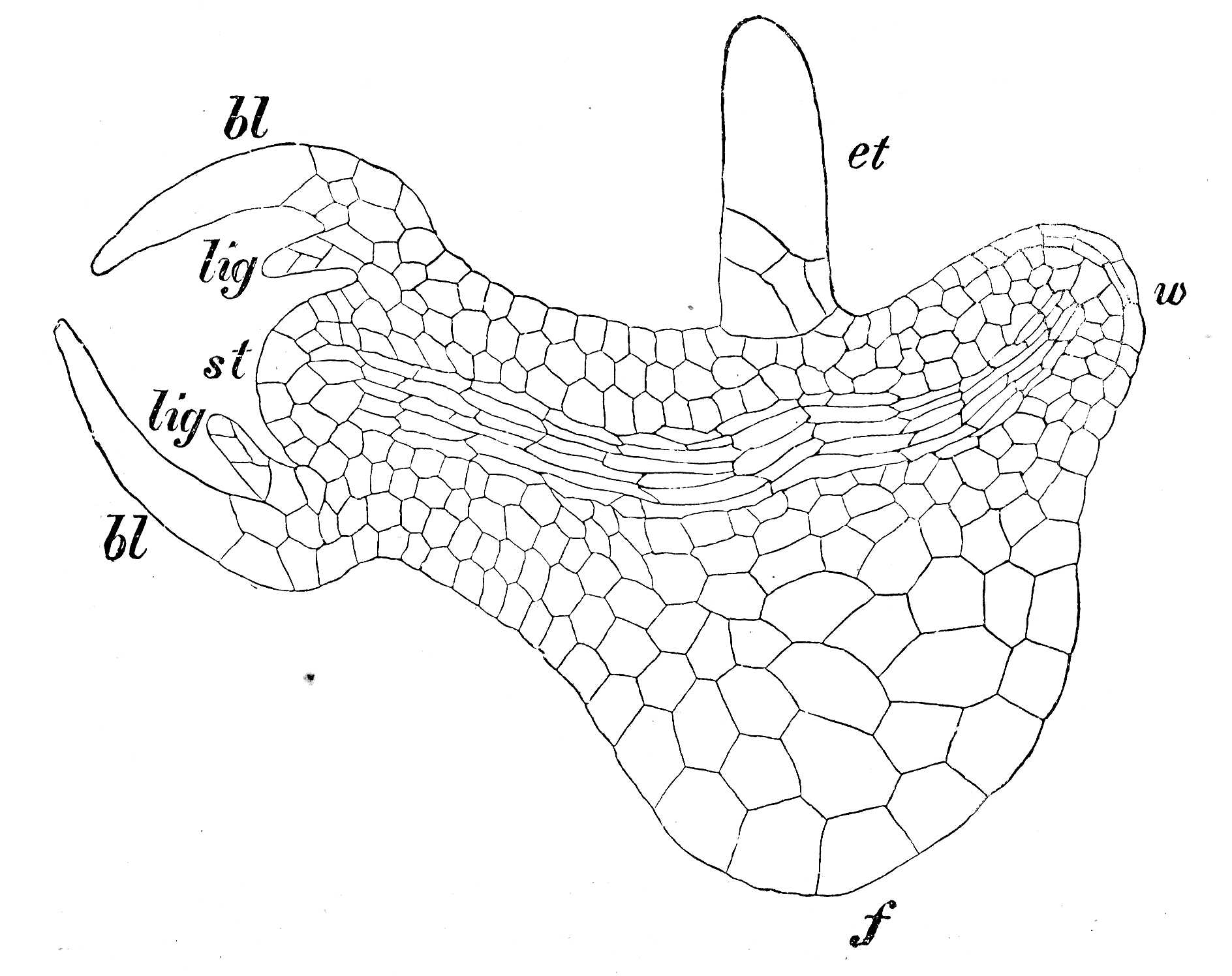
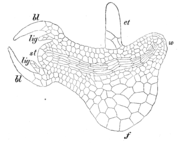 S. martensii. Längsschnitt durch Embryo. et Embryoträger, w Wurzel, f Fuß, bl Blätter, lig Ligula, st Stammscheitel
S. martensii. Längsschnitt durch Embryo. et Embryoträger, w Wurzel, f Fuß, bl Blätter, lig Ligula, st StammscheitelNach der Befruchtung teilt sich die Zygote in zwei Zellen: die obere vergrößert sich stark, teilt sich im unteren Bereich noch einige male und entwickelt sich zum Embryoträger (Suspensor). Die untere Zelle entwickelt sich zum eigentlichen Embryo, der sehr bald in ein Blattpaar, den Sprossscheitel, die Wurzel und den Fuß gegliedert ist. Bereits das erste Blattpaar im Embryo besitzt die oben beschriebenen Ligulae.
Der Suspensor schiebt den Embryo in das Prothallium hinein, das der Ernährung dient. Mit dem Fuß nimmt der Embryo die Nährstoffe auf. Der Sprossscheitel wächst nach oben, die Wurzel nach unten aus der Megaspore heraus; der junge Keimling bleibt zunächst noch mit dem Fuß im Prothalliumgewebe verankert. Er ähnelt damit einer keimenden Samenpflanze.
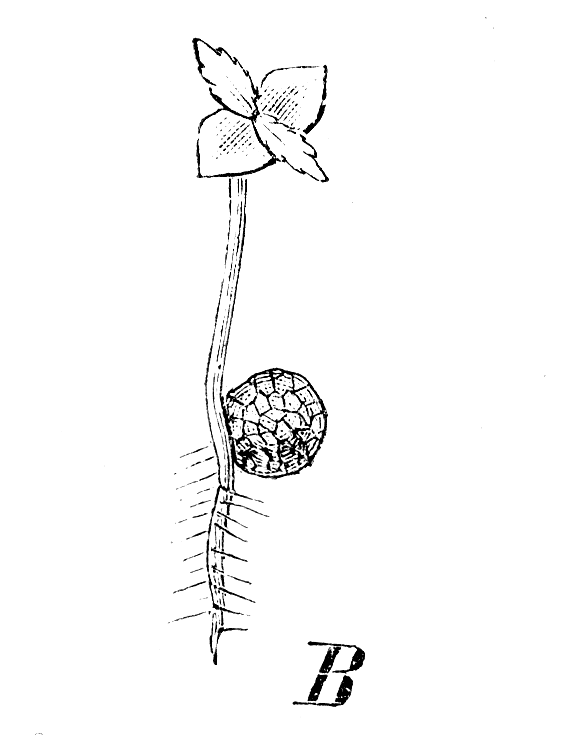
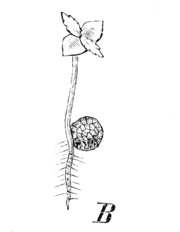 S. denticulata, Keimpflänzchen mit der Megaspore
S. denticulata, Keimpflänzchen mit der MegasporeVorkommen
Die Moosfarne haben ihren Verbreitungsschwerpunkt in den feuchten Tropenwäldern und wachsen als Bodendecker. Einige Arten kommen auch in gemäßigten Breiten vor. Die mitteleuropäischen Arten wachsen vor allem in lückigen Rasen, an Felsen und Mauern. Wenige Arten sind an Trockenstandorte angepasst. Ein Beispiel dafür ist die Unechte Rose von Jericho (Selaginella lepidophylla) aus Mittelamerika, deren Sprosse sich bei Trockenheit einrollen.
Systematik und Evolution
Die Moosfarne sind eine Ordnung der Bärlapppflanzen. Diese sind die basale Gruppe innerhalb der Gefäßpflanzen, das heißt, die Farne sind mit den Samenpflanzen näher verwandt als mit den Bärlapppflanzen. Innerhalb der Bärlapppflanzen haben genetische Studien gezeigt, dass die Moosfarne die Schwestergruppe der Isoetales sind. Das Kladogramm zeigt die Verwandtschaftsverhältnisse innerhalb der rezenten Bärlapppflanzen:[1]
┌──── Moosfarne ┌───┤ │ └──── Isoetales ───┤ │ └──────── LycopodialesIn Europa kommen fünf Arten vor, in Mitteleuropa sind nur die beiden ersten Arten heimisch:
- Schweizer Moosfarn (S. helvetica (L.) Spring
- Dorniger Moosfarn (S. selaginoides (L.) C. F. P. Martius)
- Selaginella kraussiana (Kunze) A.Braun
- Selaginella apoda (L.) Spring
- Selaginella denticulata (L.) Spring
Die ältesten Fossilien der Selaginellales sind aus dem Karbon vor 300 Millionen Jahren bekannt. Selaginellites war bereits heterospor und sah den heutigen Arten sehr ähnlich. Ansonsten ist die fossile Überlieferung dieser krautigen Sippe sehr spärlich.
Der Name Selagenilla ist die Verkleinerungsform von lateinisch selago = Tannen-Bärlapp, in Anlehnung an den ähnlichen Habitus.[2]
Bedeutung für den Menschen
Die Moosfarne haben keine wesentliche wirtschaftliche Bedeutung. Einige Arten werden als immergrüne Zierpflanzen kultiviert und gehandelt, etwa S. martensii, S. willdenowii, S. kraussiana und S. uncinata.[3] Etliche werden als Hängepflanzen genutzt. Da sie frostempfindlich sind, werden sie meist nicht im Freien gezogen.[3] Auch die Unechte Rose von Jericho wird im Handel angeboten.

Moosfarne werden auch als Zimmerpflanzen angeboten.

Gesteck mit Moosfarn

Selaginella pilifera

Selaginella flabellata
Belege und weiterführende Informationen
- P. Sitte, E. W. Weiler, J. W. Kadereit, A. Bresinsky, C. Körner: Strasburger – Lehrbuch der Botanik für Hochschulen. 35. Auflage, Spektrum Akademischer Verlag, Heidelberg 2002, ISBN 3-8274-1010-X
- W. Braune, A. Leman, H. Taubert: Pflanzenanatomisches Praktikum II. 3. Auflage, VEB Gustav Fischer Verlag, Jena 1990. ISBN 3-334-00301-9
- W. Zimmermann: Phylogenie der Pflanzen. 2. Auflage, G. Fischer, Stuttgart 1959, S 289-295. (ohne ISBN)
Einzelnachweise
- ↑ Yin-Long Qiu et al.: The deepest divergences in land plants inferred from phylogenomic evidence. In: Proceedings of the National Academy of Sciences 103(42), S.15511-15516. (online); D.L. Nickrent, C.L. Parkinson, J.D. Palmer, R.J. Duff: Multigene Phylogeny of Land Plants with Special Reference to Bryophytes and the Earliest Land Plants. In: Molecular Biology and Evolution 17 (12) 2000, S.1885-1895.
- ↑ Helmut Genaust: Etymologisches Wörterbuch der botanischen Pflanzennamen. 3. Auflage, Birkhäuser, Basel 1996, S. 574f. (Nachdruck ISBN 3-937872-16-7)
- ↑ a b Gordon Cheers (Hrsg.): Botanica, Könemann 2003. ISBN 3-8331-1600-5
Weblinks






Wikimedia Foundation.