- Steinkoralle
-
Steinkorallen 
Steinkorallen im Flachwasser
Systematik Unterreich: Vielzellige Tiere (Metazoa) Abteilung: Gewebetiere (Eumetazoa) Stamm: Nesseltiere (Cnidaria) Klasse: Blumentiere (Anthozoa) Unterklasse: Hexacorallia Ordnung: Steinkorallen Wissenschaftlicher Name Scleractinia Bourne, 1900 Steinkorallen (Scleractinia, früher Madreporaria) sind Tiere, die den Hauptanteil an der Entstehung der Korallenriffe haben, der artenreichsten marinen Lebensräume auf der Erde. Die meist winzigen, sessilen Tiere haben die Fähigkeit, an ihrer Basis Kalk abzuscheiden und bilden so Riffe und im Laufe der Jahrtausende gewaltige Riffstrukturen. Viele Inseln, z. B. die Bahamas und die Bermudas im Atlantik, die Malediven im Indischen Ozean oder Tuvalu und Kiribati im Pazifik sind durch das allmähliche Wachstum von Steinkorallen gebildet worden. Bei der Kalkbildung hilft den meisten Steinkorallen eine Symbiose mit Zooxanthellen, kleinen, einzelligen Algen aus der Gruppe der Dinoflagellaten, die auch für die Ernährung der Korallen wichtig sind.
Alle Steinkorallen leben im Meer, die meisten in den Tropen. Sie sind sehr einfach gebaute Tiere und gehören, wie Quallen, zu den Nesseltieren (Cnidaria) und darin zu den Sechsstrahligen Blumentieren (Zoantharia/Hexacorallia). Ihre nächsten Verwandten sind die Seeanemonen (Actiniaria) und andere, weniger bekannte Gruppen. Mit den nicht riffbildenden Weichkorallen (Alcyonacea) sind sie nur entfernt, mit den ebenfalls riffbildenden Feuerkorallen (Milleporidae) nur sehr entfernt verwandt.
Inhaltsverzeichnis
Verbreitung
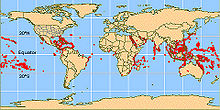 Verbreitung der tropischen Korallenriffe
Verbreitung der tropischen KorallenriffeDie Verbreitung der riffbildenden Steinkorallen wird durch die Lichtansprüche der Zooxanthellen geprägt. Außerdem sollte die Wassertemperatur 20 °C möglichst nicht unter- und 29 °C nicht überschreiten. Steinkorallen kommen deshalb überwiegend in flachen, lichtdurchfluteten, tropischen Küstengewässern vor. Ihr Verbreitungsgebiet erstreckt sich ungefähr auf einen Bereich zwischen 30° nördlicher und 30° südlicher Breite. Dabei gibt es zwei Verbreitungsschwerpunkte: Den tropischen Indopazifik, mit dem Zentrum in der Inselwelt Indonesiens, der Philippinen und Neuguineas, und die wesentlich weniger artenreiche Karibik.
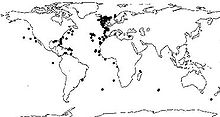 Verbreitung der Kaltwasserkoralle Lophelia pertusa
Verbreitung der Kaltwasserkoralle Lophelia pertusaDaneben gibt es aber auch Steinkorallen in gemäßigten und kalten Meeren sowie in der Tiefsee bis in 6000 Metern Tiefe. In europäischen Meeren gibt es Kaltwasserkorallen vor allem an der Küste Norwegens sowie am Kontinentalabhang unterhalb von 200 Metern.
Steinkorallen brauchen einen Salzgehalt von mindestens 2,7 % und fehlen in der Nähe von Flußmündungen und in brackisch geprägten Gewässern wie Lagunen und Nebenmeeren. Daneben verhindern Weichböden ihre Ansiedlung und trübes Wasser behindert ihr Wachstum.
Merkmale
Anatomie des Einzelpolypen
 Anatomie eines Korallenpolypen
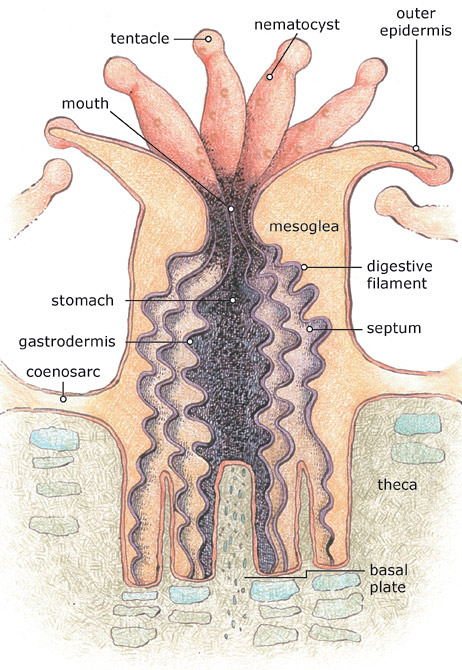
Anatomie eines KorallenpolypenSteinkorallen sind meist Kolonien aus vielen Tausenden Einzelpolypen. Jeder Polyp ist ein einzelnes Tier und ein einzelner Polyp steht auch am Beginn der Koloniebildung. Wie bei allen Nesseltieren ist ein Steinkorallenpolyp becherförmig aufgebaut und wird von zwei Zellschichten gebildet, nämlich der Außenhaut, dem Ektoderm, und dem Entoderm, das den zentralen Gastralraum umgibt. Zwischen beiden Zellschichten liegt die Mesogloea, die frei bewegliche Zellen enthält und in der der Nährstofftransport innerhalb des Polypenkörpers stattfindet. Der Gastralraum wird von mindestens sechs Mesenterien oder Septen (Sarcosepten) genannten Scheidewänden unterteilt. Dadurch wird seine innere Oberfläche vergrößert. An den Mesenterien liegen die Keimdrüsen des Polypen. Anzahl und Aufbau der Septen sind wichtige Merkmale in der Steinkorallensystematik. Im Gastralraum verdaut der Polyp seine Nahrung. Er steht durch eine zentrale Mund- und Ausscheidungsöffnung mit dem Umgebungswasser in Verbindung. Die Mundöffnung wird von sechs oder einem Vielfachen von sechs, mit Nesselzellen versehenen Tentakeln umgeben. Die Tentakeln können in einem oder mehreren Ringen angeordnet sein.
 Einzelner Polyp von Acanthastrea lordhowensis
Einzelner Polyp von Acanthastrea lordhowensisDie Polypen sitzen in einem Korallit (Korallenkelch) aus Aragonit, der in seiner Struktur die innere Anatomie des Polypenkörpers und der Septen in Form von Sklerosepten widerspiegelt und von der Fußscheibe des Polypen ausgeschieden wird. Die Sklerosepten werden bei vielen Arten durch einen Ringwall verbunden, der Theca genannt wird. Im Zentrum des Kelches befindet sich meist eine zentrale Kalksäule (Columella). Unterhalb der Polypen befinden sich Altkelche, die über waagerechte Zwischenplatten, den Tabulae, abgetrennt sind.
Bei den meisten Gattungen sind die Polypen durch ein das gesamte Korallenskelett überziehendes Gewebe miteinander verbunden, dem Coenenchym. So können sie Nahrungsstoffe austauschen und Reize weitergeben. Bei einigen großpolypigen, kolonialen Steinkorallen, wie Euphyllia paraancora, geht die Verbindung zwischen den Polypen im Laufe des Koloniewachstums verloren und die Einzelpolypen sitzen dicht an dicht an den Enden eines ansonsten toten Skelettes.
Die Größe der Einzelpolypen liegt meist bei wenigen Millimetern bis einem Zentimeter, bei einzelstehenden Steinkorallen können sie allerdings wesentlich größer werden. Der Polyp von Cynarina lacrymalis erreicht mit Wasser aufgepumpt einen Durchmesser von 35 Zentimeter, die einer großen Seeanemone zu Verwechseln ähnliche Anemonen-Pilzkoralle (Heliofungia actiniformis) gar 50 Zentimeter, mit Tentakeln von bis zu 25 Zentimetern Länge.
Meerwasseraquarianer nennen die kleinpolypigen Arten SPS-Korallen (Small Poly Scleractinia) und die großpolypigen Arten LPS-Korallen (Large Poly Scleractinia).
Wachstum
 Hirnkoralle
Hirnkoralle
Diploria labyrinthiformisKorallen wachsen, indem die Polypen einen neuen Koralliten auf ihrem alten bilden. Außerdem teilen sie sich und die neuen Einzelpolypen bilden einen neuen Korallenkelch. Es gibt die extratentakuläre Teilung bei der der neue Polyp am Rand der Basis des Elternpolyp entspringt und sofort einen neuen Koralliten bildet. Eine andere Form ist die Intratentakuläre Teilung, eine Form der Teilung, die innerhalb des Tentakelkranzes beginnt. Zunächst teilt sich die Mundöffnung. Die Mundscheide wird immer breiter, wobei die zwei Mundöffnungen auseinanderwandern. Schließlich teilt sich auch der Tentakelkranz. Als Zwischenstadium existiert ein Polyp, der zwei Mundöffnungen und zwei dicht nebeneinander sitzende Tentakelkränze hat. Die Teilung setzt sich dann von oben nach unten fort, erfasst als nächstes den Gastralraum und ist erst abgeschlossen wenn beide Polypen einen eigenen Koralliten auf dem alten gemeinsamen gebildet haben.
Steinkorallen können die unterschiedlichsten Wuchsformen haben. Koloniebildende Korallen können ast-, busch-, geweih-, tischförmig oder krustig wachsen. Andere ähneln Hirnen, Pilzen, Zungen oder Seeanemonen. Die letzten drei Wuchsformen kommen besonders bei solitären, d. h. nicht koloniebildenden Korallen vor. Arten mit unterschiedlicher Wuchsform können zur gleichen Familie, Arten mit ähnlicher Wuchsform zu unterschiedlichen Familien gehören.
Kalkbildung und Symbiose
 Querschnitt durch ein Korallenskelett
Querschnitt durch ein KorallenskelettMan unterscheidet hermatype (riffbildende) Steinkorallen, das ist die Mehrzahl, und ahermatype (nicht riffbildende) Arten. Hermatype Steinkorallen scheiden an ihrem Fuß ein Kalkskelett aus, das zu 98 bis 99,7 % aus Calciumcarbonat in der Modifikation Aragonit besteht. Sie nehmen im Meerwasser gelöste Calciumionen und Hydrogencarbonat-Ionen auf und fällen daraus Calciumcarbonatkristalle. Bei der Reaktion entsteht je ein Wasser- und Kohlenstoffdioxid-Molekül:
Da es sich hier um eine Gleichgewichtsreaktion handelt, kann der Prozess auch in die entgegengesetzte Richtung ablaufen. Das in Wasser gelöste CO2, das auch aus der Luft oder Atmung der Korallenpolypen stammt, reagiert sauer und würde das gebildete Calciumcarbonat wieder auflösen. Im tropischen Flachwasser lebenden Korallen hilft hierbei die Symbiose mit den Zooxanthellen, einzelligen Algen, die in der Haut der Korallenpolypen leben. Die Zooxanthellen betreiben Photosynthese und verbrauchen dabei das Kohlenstoffdioxid, das zusammen mit Sauerstoff zu Kohlenhydraten (Zucker) umgesetzt wird. Die so gebildeten Nährstoffe kommen auch der Ernährung der Polypen zugute. Das Zusammenleben von Zooxanthelle und Koralle ist ein klassisches Beispiel für eine echte Symbiose, da beide beteiligten Organismen deutliche Vorteile gewinnen. Die Koralle kann leichter ihr Kalkskelett fällen und wird mit zusätzlicher Nahrung versorgt, während die Algen einen geschützten Lebensraum im Körper der Polypen besitzen. Steinkorallen, die mit Zooxanthellen zusammen leben (zooxanthelate Korallen) haben eine zehn mal höhere Kalkbildungsrate als die Steinkorallen ohne symbiotische Algen (azooxanthelate Korallen). Die ein besonders poröses, ästiges Kalkskelett bildenden Acroporen erreichen Längenzuwächse von 16 bis 25 Zentimeter im Jahr. Die Masse des jährlich von Steinkorallen gebildeten Calciumcarbonat soll 900 Millionen Tonnen betragen [1].
Ernährung
 Die Sternkoralle (Astroides calycularis) ernährt sich ausschließlich durch Planktonfang
Die Sternkoralle (Astroides calycularis) ernährt sich ausschließlich durch PlanktonfangHermatypische, tropische Steinkorallen beziehen den Hauptteil der benötigten Nährstoffe von den Zooxanthellen. Sie können in gut beleuchteten Meerwasseraquarien völlig ohne Fütterung jahrelang überleben und dabei auch noch wachsen. In der Natur fangen sie jedoch, hauptsächlich in der Nacht, winzige Planktonorganismen, die nachts aus größeren Tiefen aufsteigen. Die tagsüber oft zusammengezogenen Polypen strecken dann ihre mit Nesselzellen besetzten Tentakel aus. Nicht mit Zooxanthellen in Symbiose lebende Steinkorallen wie die Kaltwasserkorallen oder in dunklen Höhlen und Felsspalten lebenden Gattungen wie Tubastraea sind vollständig auf den Nahrungsfang angewiesen. Korallen können im Wasser gelöste organische Stoffe direkt durch die Haut aufnehmen.
Konkurrenz und Aggression
 Kampf um Lebensraum, Montipora capitata überwächst Porites lobata.
Kampf um Lebensraum, Montipora capitata überwächst Porites lobata.Steinkorallen konkurrieren mit anderen sessilen Lebewesen, wie Schwämmen, Manteltieren um den Platz zum Wachsen. Untereinander sowie mit zooxanthellen Weichkorallen, Feuerkorallen und Algen stehen sie im Wettbewerb um Licht. Ein schnelles Wachstum, wie sie vor allem die meist ästigen oder buschförmigen Acroporen haben, ermöglicht es diesen Arten, andere, konkurrierende Korallen abzuschatten und zum Absterben zu bringen. Eine andere Form ist die direkte Aggression mit Hilfe von speziell ausgebildeten Wehrtentakeln, die bei Kontakt mit einer anderen Koralle aus gewöhnlichen Fresstentakeln gebildet werden. Bei Galaxea fascicularis werden die Wehrtentakeln bis über zehn Zentimeter lang, während die Fresstentakel eine Länge von nur drei bis fünf Millimetern haben. Mit Hilfe der Wehrtentakel wird das erreichbare Gewebe der konkurrierenden Koralle vernesselt und abgetötet. Generell haben kleinpolypige Korallen eine höhere Wachstumsgeschwindigkeit, sind aber bei direkter Aggression unterlegen. Großpolypige Steinkorallen wachsen langsam, können den Konkurrenten aber stark vernesseln.
Ökologische Bedeutung
 Korallenriff an der Küste der Sinai-Halbinsel, darüber Fahnenbarsche
Korallenriff an der Küste der Sinai-Halbinsel, darüber FahnenbarscheLebensraum Korallenstock
Kleine, riffgebundene Korallenfische wie Riffbarsche, Schleimfische oder Korallengrundeln sind oft auf buschförmige Korallen als Schutz angewiesen und verteidigen diese gegen Fressfeinde. Große verästelte Steinkorallenstöcke sind oft von einer roten Wolke von Fahnenbarschen, oder von einer grünblauen oder schwarzweißen der Riffbarschgattungen Chromis oder Dascyllus umgeben. Nähert sich ein Fressfeind oder ein Taucher verschwindet der oft mehrere hundert Köpfe zählende Fischschwarm zwischen den Korallenästen.
Den Büschelbarschen dienen exponiert stehende Korallenstöcke als Ansitz, von dem aus sie auf kleine Beutefische und Krebstierchen lauern. Die Fische düngen durch ihre Ausscheidungen den heimatlichen Korallenstock, der so im normalerweise extrem nährstoffarmen Wasser des Korallenriffs zusätzliche gelöste organische Stoffe erhält.
Zwischen den Ästen vieler Korallen lebt eine Vielzahl von Krebstieren, unter anderem Porzellankrebse, die ebenfalls die Korallen gegen Fressfeinde verteidigen. In speziellen, von der Koralle gebildeten Korallengallen leben Gallkrabben (Cryptochiridae). Außerdem sind symbiotische Röhrenwürmer und Bohrmuscheln bekannt. Diese Symbiosen sind noch ungenügend untersucht, und es ist nicht in jedem Fall sicher, ob und wie die Partner gegenseitig voneinander profitieren.
Fressfeinde
 Dornenkronenseestern vor der Küste Thailands
Dornenkronenseestern vor der Küste ThailandsSteinkorallen werden von Papageifischen, Falterfischen und Palettenstachlern gefressen. Während die Falterfische und Palettenstachler nur einzeln Korallenpolypen fressen, und dabei das Skelett nicht beschädigen, raspeln die Papageifische mit ihren schnabelartigen Zähnen die Korallenoberfläche und dabei das lebende Gewebe und die oberste Schicht des Skelettes ab. Gesunde Korallenstöcke überwachsen die kahl gefressenen Stellen in kurzer Zeit wieder.
Außerdem ernähren sich die kleinen, nachtaktive Schnecken der Gattungen Drupella und Coralliophila ausschließlich von Korallenpolypen.
Im Great Barrier Reef tritt von Zeit zu Zeit eine Massenvermehrung des korallenfressenden Dornenkronenseesterns (Acanthaster planci) ein.[2] Man nimmt an, das die Larven des Seesterns durch die zunehmende Eutrophierung bessere Wachstums- und Überlebenschancen haben. Auch verschiedene Arten der Kissenseesterne fressen Steinkorallenpolypen.
Bohrende Organismen
 Bohrtätigkeit von Cliona in der Schale der Meeresschnecke Nassarius reticulatus aus dem Eem
Bohrtätigkeit von Cliona in der Schale der Meeresschnecke Nassarius reticulatus aus dem EemEine Vielzahl von Organismen bohren sich auf chemischem oder mechanischem Weg in das Korallenskelett um sich vor Feinden zu schützen. Am wichtigsten sind die Bohrschwämme, meist die Gattung Clionia, die ein umfangreiches Tunnelsystem aus millimeterweiten Gängen im Skelett anlegen und die mit ihrem Gewebe die Gänge auskleiden. Von außen sind nur die millimetergroßen grün, gelb oder rot ausgekleideten Öffnungen zu sehen durch die das Wasser ein- und ausströmt. Bohrschwämme können ein Korallenskelett so stark schädigen, dass es zerbricht. Weitere bohrende Tiere sind Muscheln, besonders die Seedatteln (Lithophaga) sowie die Gattung Gastrochaena, die Eingewachsene Riesenmuschel (Tridacna crocea), Spritzwürmer, Federwürmer, Kalkröhrenwürmer und Bohrseeigel. Algen aus den Gruppen der Grün- und Rotalgen, sowie Cyanobakterien leben in jedem Korallenskelett und lösen Hohlräume wahrscheinlich mit Hilfe des CO2 aus ihrer Atmung auf. Die gesamte Bioerosion durch bohrende Organismen trägt mehr zur Zerstörung von Korallenriffen bei als mechanische Beschädigungen durch Wellenschlag, Brandung und Stürme.
Reproduktion
Sexuelle Fortpflanzung
 Steinkoralle bei der Eiabgabe
Steinkoralle bei der EiabgabeBei der sexuellen Fortpflanzung laichen die Korallenpolypen, oft gesteuert durch die Mondphasen, ab. Steinkorallen sind je nach Art zwittrig oder getrenntgeschlechtlich. Letztere geben, um eine Selbstbefruchtung zu vermeiden, Eizellen und Spermien zu unterschiedlichen Zeitpunkten ab.
Bei den Korallenarten mit interner Befruchtung geben die Korallenpolypen nur die Spermienzellen ab, und die Befruchtung der Eizellen erfolgt im Muttertier. Es werden dann, zu einem späteren Zeitpunkt, schon fertige Planula-Larven abgegeben. Die größte Anzahl der Korallenarten vermehrt sich jedoch durch externe Befruchtung, dabei geben die Korallenpolypen gleichzeitig Spermien und Eizellen ab. Die Befruchtung, durch die Masse der abgegebenen Keimzellen begünstigt, findet dann im freien Wasser statt. Die befruchteten Eizellen entwickeln sich zu Planula-Larven, die einige Tage, längstens sechs Wochen frei im Wasser treiben und sich dann an geeigneten Standorten ansiedeln. Aus der bilateral symmetrischen Planula-Larve, die sich festgeheftet hat, entwickelt sich ein radiärsymmetrischer Polyp, der ein Skelett bildet, sich weiter teilt und so den Grundstock einer neuen Kolonie bildet.
An der Küste Australiens folgt auf das Massenablaichen der Korallen eine Planktonblüte. Carnivore Zooplankter wie Krebstiere und Pfeilwürmer vermehren sich wegen des Überangebotes an Nahrung massenhaft und locken ihrerseits die planktonfressenden Walhaie an die Küste.
Eingeschlechtliche Fortpflanzung
Eine weitere Möglichkeit ist die Bildung von Planula-Larven durch Parthenogenese (Jungfernzeugung) im Gastralraum. Die Planula-Larven müssen, wie die Larven, die aus der sexuellen Fortpflanzung hervorgegangen sind, ein geeignetes Substrat zur Siedlung finden.
Asexuelle Vermehrung
 Steinkorallen
SteinkorallenZerbricht eine Korallenkolonie z. B. durch Wellenschlag, so haben die Bruchstücke, wenn sie an einen günstigen Siedlungsplatz getrieben werden, die Fähigkeit, weiter zu wachsen und eine neue Kolonie zu bilden. Diese Form der Vermehrung kommt besonders bei ästig wachsenden Geweihkorallen der Gattung Acropora vor. Diese Arten sind meist sehr schnellwüchsig. Meerwasseraquarianer nutzen diese Möglichkeit, um Steinkorallen künstlich zu vermehren.
Weitere Möglichkeiten der ungeschlechtlichen Vermehrung sind die Polypenausbürgerung und die Produktion von Anthocauli. Bei der Polypenausbürgerung lösen sich einige Polypen ohne Skelett vom Korallenstock, lassen sich treiben, siedeln sich an einer günstigen Stelle wieder an und bilden eine neue Kolonie. Besonders bei der Gattung Pocillopora ist die Polypenausbürgerung häufig. Pilzkorallen (Fungiidae) bilden an ihrer Basis Anthocauli genannte kleine Tochterpolypen, die auch schon früh ein Skelett bilden und nach einiger Zeit vom Mutterpolypen abbrechen. Bei diesen Formen der asexuellen Vermehrung entstehen Klone der Mutterkolonie.
Stammesgeschichte
Über den Ursprung der Steinkorallen gibt es zwei unterschiedliche Theorien. Zum einen wird eine Abstammung von den Rugosa, hauptsächlich solitär lebenden Korallen aus dem Paläozoikum angenommen. Dagegen spricht der Aufbau des Rugosenskeletts aus Kalzit, sowie der völlig unterschiedliche Bau der Septen. Außerdem verschwanden die Rugosen zum Ende des Perm, während die Steinkorallen erst in der Mittleren Trias erschienen. In der Unteren Trias fehlen fossile riffbildende Hexacorallia völlig. Alternativ dazu gibt es eine Hypothese über einen nicht skelettbildenden, und deshalb fossil nicht überlieferten Vorfahren aus der Verwandtschaft der Seeanemonen. Es ist nicht erwiesen, dass die Steinkorallen monophyletisch sind. [3].
Es sind ca. 5000 Arten fossiler Steinkorallen bekannt. Sie erschienen, nachdem die bisherigen Hauptriffbildner, die tabulaten Korallen (Tabulata) und die Rugosa im Oberperm ausgestorben waren. Zunächst bildeten sie nur kleine Riffe mit meist weniger als drei Metern Höhe in tieferem Wasser. Das lässt darauf schließen, das sie noch nicht mit Zooxanthellen in Symbiose lebten. Diese Lebensgemeinschaft kam wahrscheinlich erst in der oberen Trias oder im frühen Jura auf, als die Riffe größer wurden und mehr als 20 Arten an der Riffbildung beteiligt waren[4]. Im Jura erschienen die Caryophylliina, die viele solitäre Arten umfassen und mit Favia auch eine heute noch lebende Gattung. In der Unterkreide erschienen die Kaltwasserkorallen Oculina und Madrepora. Seit der Oberkreide gibt es die Dendrophylliina und die Acroporidae, die ein sehr leichtes Skelett haben, am schnellsten wachsen und heute etwa 40 % der Korallenfauna stellen. Die Gattungen Acropora, Galaxea, Fungia, Pocillopora und Seriatopora kamen im Verlauf des Tertiär hinzu. Am Beginn des Miozän, vor 23 Millionen Jahren, kam es zu einer großen Aussterbewelle unter den Steinkorallen. Die meisten heutigen Gattungen entstanden in den letzten 15 Millionen Jahren.
Systematik
Äußere Systematik
 Scheibenanemonen (Discosoma sp.)
Scheibenanemonen (Discosoma sp.)Steinkorallen gehören zu den Nesseltieren (Cnidaria) und dort zu den Blumentieren (Anthozoa), die sich von allen anderen Nesseltieren unter anderem dadurch unterscheiden, dass sie nie ein Medusenstadium haben. Innerhalb der Blumentiere gehören sie aufgrund der Anzahl ihrer Tentakel und weiterer Merkmale zu den Zoantharia, die auch Sechsstrahlige Blumentiere (Hexacorallia) genannt werden, und zu denen auch die Seeanemonen (Actiniaria), die Scheibenanemonen (Corallimorpharia), die Krustenanemonen (Zoanthidea) sowie die Zylinderrosen (Ceriantharia) und die Schwarzen Korallen (Antipatharia) gehören. Wegen einer Vielzahl von Gemeinsamkeiten in der Anatomie der Polypen gelten die Scheibenanemonen als die mit den Steinkorallen am nächsten verwandte Gruppe und werden oft als deren Schwestergruppe angesehen.
Eine phylogenetische Untersuchung aus dem Jahr 2006 sieht die Scheibenanemonen jedoch als Steinkorallen, die ihr Kalkskelett verloren haben. Einige Steinkorallengattungen, darunter Acropora, haben mehr Apomorphien mit den Scheibenanemonen gemeinsam, als mit anderen Gattungen der Steinkorallen. Daraus folgt, dass die Steinkorallen ohne Einbeziehung der Scheibenanemonen ein paraphyletisches Taxon und die Scheibenanemonen Steinkorallen ohne Skelett sind, von den Autoren der Studie Naked corals genannt. Der Ursprung der Scheibenanemonen soll in der unteren oder mittleren Kreidezeit, vor 132 bis 110 Millionen Jahren liegen, als die Kalkbildung durch einen abnehmenden pH-Wert des Meerwassers erschwert wurde. [5]
Innere Systematik
 Blasenkoralle (Plerogyra sp.)
Blasenkoralle (Plerogyra sp.) Pilzkoralle (Fungia sp.)
Pilzkoralle (Fungia sp.) Eusmilia fastigiata
Eusmilia fastigiata Steinkorallenpolypen
SteinkorallenpolypenIm Allgemeinen werden die Steinkorallen heute in 5 Unterordnungen und 16 Familien mit mehr als 1000 Arten eingeteilt. Die Klassifikation beruht auf morphologischen Merkmalen. Ursprünglich wurden über 2500 Arten beschrieben, viele aber nach Revisionen als lokale Varietäten schon bekannter Arten erkannt. Die hier wiedergegebene Systematik zeigt die Unterordnungen und Familien und folgt ITIS: [6]
- Astrocoeniina
- Caryophylliina
- Caryophylliidae
- Flabellidae
- Gardineriidae
- Guyniidae
- Turbinoliidae
- Dendrophylliina
- Faviina
- Anthemiphylliidae
- Faviidae
- Meandrinidae
- Merulinidae
- Mussidae
- Oculinidae
- Pectiniidae
- Rhizangiidae
- Trachyphylliidae
- Fungiina
- Agariciidae
- Fungiacyathidae
- Fungiidae
- Micrabaciidae
- Poritidae
- Siderastreidae
- Thamnasteriidae
Der australische Steinkorallentaxonom J.E.N. Veron vertritt eine andere Auffassung und teilt alle Steinkorallenarten - einschließlich der fossilen arten - dreizehn Unterordnungen zu, von denen sieben auch heute noch lebende Vertreter haben.[7]
Unterordnungen nach J.E.N. Veron:
- Stylophylliina †
- Pachythecalina †
- Archaeocoeniina
- Archaeofungiina †
- Stylina †
- Fungiina
- Astraeoina
- Faviina
- Caryophylliina
- Distichophylliina †
- Meandriina
- Poritiina
- Dendrophylliina
Inzwischen gibt es erste Ansätze zu einer phylogenetischen Systematik der Steinkorallen. Bei einer Analyse der Ribosomen RNA von 88 Arten, die 71 Gattungen und 20 Familien repräsentierten zeigte es sich, dass es zwei bisher unbenannte monophyletische Hauptkladen gibt, die vorläufig als robuste und komplexe Klade bezeichnet werden. Die Monophylie von Verons Unterordnungen Faviina, Poritiina und Dendrophylliina wurde bestätigt, die der Archaeocoeniina, Fungiina, Caryophylliina und Meandriina, sowie der Familien Faviiidae, Caryophylliidae, Poritidae und Oculinidae aber nicht. Zur robusten Klade gehören u. a. die Dendrophylliina, die Poritiina, die Fungiidae, die Siderastreidae, die Pocilloporidae, die Merulinidae, die Meandrinidae, die Oculininae sowie die Gattungen Euphyllia und Catalaphyllia. Die komplexe Klade umfasst u. a. die Acroporidae, die Agariciidae, die Galaxeinae sowie die Fungiacyathidae. [8] [9] Die Teilung in zwei Hauptkladen wurde auch von den Autoren der Studie über die Zugehörigkeit der Scheibenanemonen zu den Scleractinia bestätigt. Die Scheibenanemonen bilden zusammen mit der komplexen Klade ein unbenanntes Taxon, das die Schwestergruppe der robusten Klade bildet. Die beiden Taxa trennten sich vor 288 bis 240 Millionen Jahren. [5]
Korallenkrankheiten
Korallenbleiche [10]
 Korallenbleiche
KorallenbleicheAls Korallenbleiche wird das Phänomen bezeichnet, dass Steinkorallen unter Stress ihre Zooxanthellen ausstoßen und danach absterben. Die Korallen verlieren dabei ihre Farbe, da besonders die bräunlichen Töne von den symbiotischen Algen in der Haut kommen. Im Jahre 1988 wurde auf einem Symposium zu Korallenriffen in Townsville in Australien zum ersten Mal auf das weltweite Ausmaß des Problems aufmerksam gemacht. Das Erbleichen von Korallen wurde auch schon vorher beobachtet, z. B. nach starkem Regen und damit verbundenen Süßwassereintrag oder nach starkem Niedrigwasser. Das Phänomen trat aber immer nur lokal auf. Das weltweite Korallensterben wird heute mit der erhöhten Temperatur des Oberflächenwassers in Folge der globalen Erwärmung in Verbindung gebracht. Am stärksten trat die Korallenbleiche im Jahre 1998 während eines El Niño, im westlichen Pazifik und im Indischen Ozean auf. Monatelang lagen die Temperaturen 1 bis 3 °C über dem langjährigen Durchschnitt. Die Malediven büßten dabei 98 % der oberflächennahen Korallenfauna ein.
Black Band Disease [11]
 Black Band Disease
Black Band DiseaseDie als Black Band Disease beschriebene Korallenkrankheit tritt hauptsächlich in der Karibik auf, und wurde Anfang der 1970er Jahre zum ersten mal an der Küste Belizes registriert. Charakteristisch ist ein schwarz-violettes Band das langsam über das lebende Gewebe der Koralle kriecht, das Gewebe dabei zersetzt und ein entblößtes, totes Korallenskelett hinter sich lässt. Das tote Skelett wird schnell von Algen besiedelt. In dem Band wurden verschiedene einzellige Mikroorganismen gefunden, wie die Cyanobakterie Phormidium corallytium, Pilze, sowie weitere heterotrophe gramnegative, Sulfat reduzierende und Schwefel oxidierende Bakterien. Die Bakterienflora soll ein Mikromilieu erzeugen, das zum Absterben des Korallengewebes führt. Bis heute tritt das Black Band Disease im Indopazifik nur vereinzelt auf.
Gefährdung durch den Menschen
Steinkorallen werden heute durch anthropogene Einflüsse auf viele Arten gefährdet. Durch die Klimaerwärmung gibt es immer mehr Zeiten, in denen die Temperatur des Oberflächenwassers bei 30 °C oder höher liegt, so dass es vermehrt zur Korallenbleiche kommt. Außerdem werden Steinkorallen und andere Bewohner der Korallenriffe durch Abwässer von Industrie und Landwirtschaft, durch die Fischerei mit Dynamit und Cyanid gefährdet. Auf den Malediven wird Korallenkalk als Baumaterial für Gebäude und im Straßenbau eingesetzt. In Indonesien wird der gelöschte Kalk, der zum Genuss der Betelnuss nötig ist, oft aus Korallenkalk gewonnen. Die touristische Erschließung der Riffe führt zu direkten Zerstörungen durch das Ankern von Yachten und Ausflugsbooten an den Riffen und durch unvorsichtige Schnorchler und Taucher.
Für die Meerwasseraquaristik werden immer noch lebende Steinkorallen gesammelt. Das ist besonders bei seltenen, massiven, großpolypigen Arten, die man nicht durch einfache Fragmentation vermehren kann, ein Problem. Einige besonders farbige und gefragte Arten sind in vielen Riffen schon nicht mehr zu finden.
In der Zukunft wird die Versauerung der Meere zu einem großen Problem für Steinkorallen und andere ein Kalkskelett bildende Organismen werden, da der sinkende pH-Wert die Skelettbildung behindert [12] [13].
Quellen und weiterführende Informationen
Zitierte Quellen
Die Informationen dieses Artikels entstammen zum größten Teil aus den unter Literatur angegebenen Quellen, darüber hinaus werden folgende Quellen zitiert:
- ↑ Fossa, Nielsen, Seite 295
- ↑ A. J. Nielsen: Forschung im Great Barrier Reef, in Koralle, Meerwasseraquaristik-Fachmagazi, Nr. 42, 2006/2007, Seite 46–49
- ↑ Bernhard Ziegler: Einführung in die Paläobiologie, Tl.2, Spezielle Paläontologie, Protisten, Spongien und Coelenteraten, Mollusken. Schweizerbart’sche Verlagsbuchhandlung, ISBN 3510650360
- ↑ Steven M. Stanley: Historische Geologie: eine Einführung in die Geschichte der Erde und des Lebens. Spektrum, Akad. Verlag, Heidelberg/Berlin/Oxford 1994, ISBN 3860251538, Seite 431
- ↑ a b M. Medina, A. G. Collins, T. L. Takaoka, J. V. Kuehl, & J. L. Boore: Naked corals: Skeleton loss in Scleractinia PDF
- ↑ Integrated Taxonomic Information System: ITIS-Seite Scleractinia
- ↑ Sandra L. Romano & Stephen D. Cairns: Scleractinia The Tree of Life Project
- ↑ S.L.Romano, S.D.Cairns: Molecular Phylogenetic Hypotheses for The Evolution of Scleractinian Corals, Bulletin of Marine Science, Volume 67, Number 3, November 2000, Seiten 1043–1068 [1]
- ↑ Jarosloaw Stolarski, Ewa Roniewicz: Towards a new synthesis of evolutionary relationships and classification of scleractinia, Journal of Paleontology, Nov 2001
- ↑ H. Schumacher, K. u. W. Loch, W. R. See. Das Erbleichen der Korallen. In Biologie in unserer Zeit 3/2005: 186–191
- ↑ D. Combosch, H. Schumacher. Biotische Korallenkrankheiten. In Biologie in unserer Zeit 3/2005: 178–184
- ↑ H. Schumacher: Korallenriffe sind weltweit bedroht, in Biologie der Meere, 1991, Spektrum Akad. Verl., ISBN 3-89330-753-2
- ↑ Wissenschaftlicher Beirat der Bundesregierung Globale Umweltveränderungen (2006): Die Zukunft der Meere – zu warm, zu hoch, zu sauer. Sondergutachten, Berlin (PDF, 3,5 MB)
Literatur
- Helmut Schumacher: Korallenriffe, BLV Verlagsgesellschaft München, 1988, ISBN 3-405-13614-8
- Yossi Loya und Ramy Klein: Die Welt der Korallen, Jahr Verlag Hamburg, 1998, ISBN 3-86132-226-9
- Julian Sprung: Korallen, Dähne Verlag, 2000, ISBN 3-92168-487-0
- S. A. Fossa, A. J. Nilsen: Korallenriffaquarium, Band 4, Schmettkamp Verlag, 1995, ISBN 3-928819-05-4
- J. Sprung, J.C. Delbeek: Das Riffaquarium Band 1, 1994, ISBN 1-883693-17-9
- Baensch/Patzner: Mergus Meerwasser-Atlas Bände 2, 4 + 5, Mergus-Verlag, Melle
- Rainer Kaiser: Niedere Tiere tropischer und kalter Meere im Aquarium. Ulmer, Stuttgart 1991, ISBN 3800172224
- T. F. Goreau, N. I. Goreau, T. J. Goreau: Korallen und Korallenriffe, in Biologie der Meere, 1991, Spektrum Akad. Verl., ISBN 3-89330-753-2
Weiterführende Literatur
- J.E.N. Veron: Corals in Space and Time: The Biogeography and Evolution of the Scleractinia, Cornell University Press, 1995, ISBN 0-80148-263-1
Weblinks
- Romano, Sandra L. and Stephen D. Cairns. 2002. Scleractinia. Stony star corals. Version 28 October 2002 (under construction). The Tree of Life Web Project
- Online description of reef building corals CoralSearch
- Kleinpolypige Steinkorallen im Meerwasser-Lexikon (dt.)
- Großpolypige Steinkorallen im Meerwasser-Lexikon (dt.)
- Steinkorallen im Seewasserlexikon (dt.)





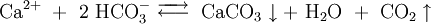
















Wikimedia Foundation.