- Vierblättriges Kleeblatt
-
Klee 
Wiesen-Klee (Trifolium pratense), die Typusart der Gattung
Systematik Ordnung: Schmetterlingsblütenartige (Fabales) Familie: Hülsenfrüchtler (Fabaceae) Unterfamilie: Schmetterlingsblütler (Faboideae) Tribus: Trifolieae Gattung: Klee Wissenschaftlicher Name Trifolium L. Klee (Trifolium) ist eine 243 Arten umfassende Pflanzengattung. Sie wird zur Unterfamilie der Schmetterlingsblütler (Faboideae) aus der Pflanzenfamilie der Hülsenfrüchtler (Fabaceae) gezählt. Im Sprachgebrauch werden allerdings häufig nicht nur Arten dieser Gattung mit Klee bezeichnet, sondern auch Arten aus den nahverwandten Gattungen Schneckenklee (Medicago) und Steinklee (Melilotus) sowie aus der taxonomisch weit entfernten Gattung Sauerklee (Oxalis). Charakteristische Merkmale der Gattung sind dreifiedrige Blätter und vielblütige, kopfige Blütenstände. Klee ist auf allen Kontinenten, mit Ausnahme Australiens und Antarktikas, natürlich verbreitet. In Australien wurden verschiedene Kleearten durch den Menschen eingeführt.
In der Landwirtschaft werden 16 verschiedene Kleearten als Futterpflanzen verwendet. Kleeanbau ist sowohl in Europa als auch in Nordamerika von ökonomischer Bedeutung. In Europa wurde Wiesen-Klee (Trifolium pratense) schon ab dem 4. Jahrhundert nach Christus angebaut. Durch die Fähigkeit, in Symbiose mit Knöllchenbakterien (Rhizobiaceae) an den Wurzeln Stickstoff aus der Luft zu binden, ist Klee auch für die Verbesserung der Bodenfruchtbarkeit bedeutend.
Das Kleeblatt fand in die Symbolik vieler Kulturen Eingang. Es repräsentiert stets die Dreiheit, wie zum Beispiel im Christentum als Symbol der Dreifaltigkeit. Weiterhin steht Klee für den Sommer oder symbolisiert die Liebe. Das vierblättrige Kleeblatt gilt als Glücksbringer.
Inhaltsverzeichnis
Beschreibung
Kleearten sind zumeist ausdauernde, selten aber auch einjährige und sehr selten zweijährige krautige Pflanzen. Fast alle Arten verzweigen sich direkt an der Basis oder entlang der ganzen Sprossachse. Einige wenige Arten entwickeln jedoch über einem dichten Büschel vieler Blätter direkt an der Basis einen langen, blattlosen und unverzweigten Schaft. Die Sprossachse kann aufrecht, liegend oder kriechend sein. Viele Kleearten, wie zum Beispiel der Kaukasische Klee (Trifolium ambiguum), bilden Rhizome aus. Falls Rhizome vorhanden sind, machen sie häufig einen erheblichen Teil der Trockenmasse der Pflanzen aus, beim Kaukasischen Klee sind dies etwa 44 % der Gesamtmasse.[1] Weniger verbreitet ist die Entwicklung von Ausläufern (Stolonen). Sie findet sich beispielsweise beim Weiß-Klee (Trifolium repens). Die Dichte der Behaarung der Pflanzen variiert von Art zu Art. Das Spektrum reicht von dicht behaart bis fast kahl oder gänzlich unbehaart. Die Ausrichtung der Haare kann anliegend oder abstehend sein. Sie sind nach oben oder nach unten gerichtet.
Wurzeln
Das Wurzelsystem der Kleearten ist häufig fischgrätenförmig. Die Form variiert dabei abhängig vom Substrat nur schwach, lediglich die Anzahl der Verzweigungen scheint in Sandböden höher zu sein als in Lehmböden.[2] Einjährige Arten haben feinfasrige Wurzeln, ausdauernde Arten haben verdickte holzige Wurzeln und selten sogar Knollen.
Wurzelanschwellungen (Knöllchen), die durch Knöllchenbakterien hervorgerufen werden, sind wie bei allen Hülsenfrüchtlern typisch. Die verschiedenen Kleearten wurzeln unterschiedlich tief. Trifolium purpureum beispielsweise wurzelt bis zu 2,1 Meter tief, wohingegen Wurzeln des Schild-Klees (Trifolium clypeatum) selten tiefer als 40 Zentimeter in das Substrat eindringen. Bei zehn untersuchten Arten betrug der Anteil der Wurzeln an der Trockenmasse der Pflanzen zwischen 14 % und 30 %.[3]
Blätter
 Typisches Kleeblatt (hier vom Weiß-Klee (Trifolium repens)) – Gut sichtbar sind die fast parallelen, lateral verlaufenden Blattrippen
Typisches Kleeblatt (hier vom Weiß-Klee (Trifolium repens)) – Gut sichtbar sind die fast parallelen, lateral verlaufenden BlattrippenDie Laubblätter sind bei fast allen Arten dreizählig gefiedert. Nur Arten der Untersektion Lupinaster in der Sektion Lotoidea haben fünf- bis neunzählig gefingerte Blätter. Die Blätter sind in der Regel wechselständig. Bei einigen Arten erscheinen die oberen Blätter gegenständig; hier stehen die oberen Knoten so dicht beieinander, dass sich ein falscher Eindruck von Gegenständigkeit ergibt. Eines der beiden Blätter ist in diesem Fall aber häufig nicht voll entwickelt.
Die unteren (basalen) Blätter weichen in Größe und Gestalt häufig markant von den oberen Blättern ab. Die Fiedern sind hier häufig eiförmig, umgekehrt-eiförmig oder breit-elliptisch, immer sind sie deutlich breiter als die Fiedern der oberen Blätter. Die Fiederform ist ein relativ stabiles Merkmal. Sie kann jedoch in geringem Umfang sowohl an einem Individuum als auch innerhalb einer Art variieren. Der Blattrand ist ganzrandig oder fein bis grob gezähnt. Die Spreite ist beidseitig oder nur auf der Blattunterseite behaart, bei einigen Arten aber auch kahl. Die Blattnervatur ist innerhalb der Arten einheitlich. Die von der Hauptrippe abzweigenden, sekundären Blattrippen verlaufen annähernd parallel. Sie oder ihre Verzweigungen erreichen den Blattrand.
Auch die Gestalt der Nebenblätter ist innerhalb bestimmter Gruppen stark konserviert (einheitlich). Das Spektrum reicht von schmal und abrupt zugespitzt in der oberen Hälfte bis breit-elliptisch. Sie sind in allen Sektionen außer Involucrarium und Paramesus ganzrandig, bei diesen sind sie gezähnt, oder tief eingeschnitten mit lanzettlichen Lappen. Bei den Arten der afrikanischen Untersektion Ochreata sind die Blattstiele der Laubblätter auf ganzer Länge mit den Nebenblättern verwachsen und die Internodien so ummantelt. Ähnliche Verwachsungen, allerdings nur bei den Blattstielen der oberen Laubblättern und nicht so ausgeprägt, existieren aber auch bei anderen Arten.
Blütenstände
 Ährenartiger Blütenstand beim Schmalblättrigen Klee (Trifolium angustifolium)
Ährenartiger Blütenstand beim Schmalblättrigen Klee (Trifolium angustifolium)Die Blütenstände sind für gewöhnlich achselständig und gestielt oder sehr selten stiellos sitzend. Bei einigen wenigen Arten sind sie aber auch endständig. Meistens sind sie kopfig und vielblütig. Blütenstände mit wenigen Einzelblüten sind die Ausnahme.
Der Botaniker Michael Zohary untersuchte 1972 die Formen der Blütenstände in der Sektion Lotoidea. Dort ergab sich, dass fast alle Arten rispenartige Blütenstände ausbilden, sich jedoch eine Tendenz zu ährenartigen Blütenständen abzeichnet. Evolutionär höher entwickelte Arten scheinen auch kürzere Blütenstiele zu bilden. Es ist jedoch unklar, ob diese Ergebnisse für die ganze Gattung repräsentativ sind.[4]
Tragblätter finden sich vor allem in den primitiveren Sektionen der Gattung, immer zusammen mit rispenartigen Blütenständen. Insgesamt sind zwei verschiedene evolutionäre Trends in der Gattung erkennbar: auf der einen Seite die Zurückbildung zu Tragblatt-Rudimenten oder das vollkommene Verschwinden der Tragblätter und auf der anderen Seite die Aggregation der Tragblätter direkt unter dem Kopf, was zu Involukralbildung (Hüllblättern) führt. Ausgeprägte Tragblätter gibt es zum Beispiel in der Sektion Mistyllus, die zu den evolutionär primitiven Sektionen der Gattung zählt. Rudimentär oder nicht vorhanden sind die Tragblätter zum Beispiel in der Sektion Trifolium, Hüllblätter finden sich beispielsweise in der Sektion Involucrarium.
Blüten
Der Kelch in der Gattung Klee ist zumeist symmetrisch und röhren- oder glockenförmig mit fünf nahezu gleichmäßigen Kelchlappen oder Zähnen. Drei Variationen des Kelchs sind verbreitet: Bei den Arten der Untersektionen Loxospermum, Calycospatha und der Sektion Mistyllus ist die Kelchröhre blasenförmig verdickt. Bei einigen anderen Arten, vor allem in der Sektion Chronosemium, sind die oberen beiden Kelchzähne gegenüber den drei unteren verkürzt. In der Sektion Trifolium finden sich schließlich häufig geschlossene Kelchröhren.
Kleeblüten sind fünfzählige Schmetterlingsblüten. Die Krone variiert in der Farbe von rein-weiß über gelb bis dunkel-purpurn. Zweifarbige Kronen sind verbreitet. Häufig wechselt die Farbe nach der Anthese (Vorgang des Blühens) und die Blütenstände erscheinen mehrfarbig. Die vier unteren Kronblätter (Schiffchen und Flügel) sind lose verwachsen und an die Staubblätter angewachsen. Das fünfte Kronblatt (die Fahne) ist frei und vergrößert. Die Flügel sind in der Regel deutlich länger als das Schiffchen. Die Staubblätter sind zehn an der Zahl, neun von ihnen sind zu einem U-förmigen Tubus verwachsen. An der Basis des zehnten, oberen, freien Staubblatts befindet sich eine kleine Honigdrüse (Nektarium). Bei einigen Arten gibt es die Tendenz zu einer Reduktion der Anzahl der Pollensäcke pro Staubblatt, etwa bei Trifolium badium, repens, campestre und dubium; in einer Blüte kommen Staubblätter mit den normalen vier, aber auch drei und zwei Pollensäcken vor, wobei jeweils die ventralen Pollensäcke reduziert werden.[5] Der Fruchtknoten liegt innerhalb des Tubus aus den Staubblättern. Die Krone ist in der Regel frühzeitig abwerfend und nur in einigen Arten bleibend. In der Sektion Mistyllus trocknet die verbleibende Krone ein. Nur in der Sektion Chronosemium wirkt sie unterstützend auf das Verwehen der Samen (Windbestäubung).
Pollen
Die Pollenkörner der Gattung sind prolat, das bedeutet sie haben die Form von Rotationsellipsoiden. Die Exine, das heißt die äußere Schicht des Sporoderms (der Wand des Pollenkorns), sind tricolporat, das heißt, sie besitzt drei längsgestreckte Keimfalten (Colpi), in deren Mitte sich je eine rundliche Keimpore (Porus) befindet. Die Pollenkörner haben einen Durchmesser von 14 bis 35 Mikrometer. Sie sind 21 bis 49 Mikrometer lang und haben ein Verhältnis von Länge zu Durchmesser zwischen 1,19 und 1,64.
Die kleinsten Pollenkörner finden sich beim Kleinblütigen Klee (Trifolium retusum) und beim Schmächtigem Klee (Trifolium suffocatum). Die größten Pollenkörner wurden beim Wiesen-Klee (Trifolium pratense) beobachtet. Bei einigen amerikanischen Arten, wie Trifolium albopurpureum, sind die Pollenkörner fast kugelförmig. Die Oberflächenstruktur (Skulpturierung) der Pollenkörner variiert innerhalb der Gattung stark.
Früchte
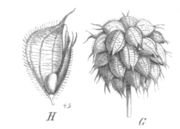 Illustration der Früchte des Erdbeer-Klees (Trifolium fragiferum) – ein- bis zweisamige Hülsen, die an zwei Nähten aufreißen
Illustration der Früchte des Erdbeer-Klees (Trifolium fragiferum) – ein- bis zweisamige Hülsen, die an zwei Nähten aufreißenAls ursprüngliche Fruchtform der Gattung werden vielsamige, an zwei Nähten aufreißende Hülsenfrüchte angesehen. Innerhalb der Gattung gibt es sowohl diesen archaischen Fruchttyp als auch viele evolutionär davon abgeleitete Formen, die sich der Schließfrucht immer weiter annähern. In der Sektion Lotoidea, in der die meisten evolutionär primitiven Charakteristika der Gattung erhalten sind, findet sich eine ganze Anzahl von Arten, wie zum Beispiel der Vogelfuß-Klee (Trifolium ornithopodioides) oder der Westalpen-Klee (Trifolium alpinum), mit solchen typischen, vielsamigen Hülsen, die an zwei Nähten aufplatzen. Von dieser Ausgangsform existieren viele abgeleitete Formen: im extremen Fall ein einsamiger Schlauch (Utriculus) oder ein Nüsschen mit einer sehr dünnen Fruchtwand (Perikarp), die in einigen Fällen sogar nur aus Epidermiszellen (Abschlussgewebe) besteht. Bei diesen extrem reduzierten Hülsen fehlt die Naht, und so kann die Frucht nicht mehr aufreißen, sondern teilt sich querlaufend oder unregelmäßig. Solche Früchte sind vor allem in der Sektion Trifolium, zum Beispiel beim Ausgebreiteten Klee (Trifolium diffusum), aber auch in vielen anderen Sektionen und sogar auch in Lotoidea zu finden.
Eine Zwischenform ist die einsamige Hülse, bei der beide Nähte vorhanden sind. Früchte dieser Form gibt es zum Beispiel beim Usambara-Klee (Trifolium usambarense), bei Trifolium burchellianum aber auch bei vielen anderen Arten. Eine weitere Zwischenform ist die einsamige Hülse mit nur noch einer Naht. Dieser Fruchttyp findet sich in mehreren Sektionen, ist aber eher selten.
Samen
 Samen des Schweden-Klees (Trifolium hybridum)
Samen des Schweden-Klees (Trifolium hybridum)Die Gestalt der Samen ist typischerweise eiförmig, ellipsoid oder länglich-ellipsoid und seitlich gestaucht, variiert aber in Dimension, Gewicht und Form. Variationen der Form resultieren aus der unterschiedlichen Position des Hilums, der Ansatzstelle des Nabelstrangs (Funiculus) über den der Samen mit dem Fruchtknoten verbunden ist. Das Hilum trennt im Embryo die Wurzelanlage (Radicula) von den Keimblättern (Kotyledonen).
Bei einigen Arten, zum Beispiel in den drei Untersektionen Lotoidea, Oxalioidea und Ochreata in der Sektion Lotoidea, liegt das Hilum endständig. Hier sind Wurzelanlage und die Keimblätter fast gleich lang. Bei fast allen anderen Arten liegt das Hilum seitlich oder am Rand. Dann ist die Wurzelanlage deutlich kürzer als die Keimblätter. Die Samenschale (Testa) kann glatt, angeraut, warzig, faltig oder körnig sein. Die Gestalt der Testa scheint aber nicht mit der taxonomischen Einteilung der Gattung zu korrelieren.
Zellbiologie
Etwa 70 % der Arten der Gattung wurden auf die Anzahl der Chromosomen untersucht. Die Arten sind üblicherweise diploid, enthalten also zwei Chromosomensätze pro Zelle. Es treten aber auch andere Ploidiegrade auf. Bei 38 Arten tritt Polyploidie, oder Polyploidie und Diploidie in zwei Linien nebeneinander, auf. Von diesen sind 31 mehrjährige aber nur sieben einjährige Arten.[6]
Die Basiszahl, das heißt die Anzahl der Chromosomen eines einzelnen Satzes, ist für 80 % der untersuchten Arten acht, für 15 % der Arten sieben, sechs in zwei Prozent und fünf in drei Prozent der Fälle. Auch bei den anderen Arten der Tribus Trifolieae scheint die Basiszahl acht zu dominieren. Reduzierte Chromosomenzahlen und Aneuploidie, das heißt, dass einzelne Chromosomen zusätzlich zum üblichen Chromosomensatz vorhanden sind oder fehlen, sind vor allem in den Sektionen Trichocephalum und Trifolium verbreitet. Dies ist ein weiteres Indiz dafür, dass diese Sektionen stärker abgeleitete Formen sind als die anderen.
Die Gestalt der Chromosomen wurde bei 14 afrikanischen und 15 europäischen diploiden Arten untersucht.[7][8] Es gab jeweils ein bis drei Paare metazentrischer (Centromer in der Mitte), vier bis sechs Paare submetazentrischer (Centromer zwischen Mitte und Ende) und ein Paar akrozentrischer Chromosomen (Centromer am Ende, der kürzere Arm sehr klein).
Verbreitung
 Verbreitungskarte der Gattung
Verbreitungskarte der GattungKlee ist sehr weit verbreitet und in insgesamt vier Florenreichen vertreten, der Neotropis, Paläotropis, Holarktis und Capensis. Die Gattung fehlt jedoch in der Australis. Das Verbreitungsgebiet kann in drei distinkte Zentren aufgeteilt werden: Eurasien, Afrika und Amerika.
Etwa 59 % aller Arten sind in Eurasien beheimatet. Das Mannigfaltigkeitszentrum, das heißt das Gebiet mit der höchsten Artenvielfalt (insgesamt 110 Arten aus sieben Sektionen), liegt im Mittelmeerraum. Im Euro-Sibirischen Raum und im Turan-Iranischen Raum finden sich hingegen zusammen nur 8 % der Arten. In fast jedem Mittelmeer-Anrainerstaat existieren einige endemische Arten. Die meisten Endemiten gibt es jedoch in der Türkei, mit insgesamt neun endemischen von den 100 dort natürlich vorkommenden Arten. Die Nordgrenze des Verbreitungsgebietes verläuft durch Schweden und Norwegen und erreicht hier etwa den nördlichen Polarkreis. Nach Osten hin nimmt die natürliche Artenvielfalt ab und nur die beiden Arten Trifolium lupinaster und Trifolium gordejevi sind im Fernen Osten Russlands, auf Sachalin und in Japan zu finden.
In Afrika sind 15 % aller Arten verbreitet. Der Großteil der in Nordafrika vorkommenden Arten ist wie in Europa im Mittelmeerraum heimisch. Hier existieren aber nur zwei endemische Arten, beide im Atlasgebirge. Die Hälfte aller nordafrikanischen Arten werden ebenso auf den Makaronesischen Inseln gefunden. Fast alle nicht in Europa vorkommenden Arten finden sich im Hochland von Abessinien. Klee fehlt auf der Arabischen Halbinsel, mit Ausnahme des jemenitischen Hochlands. Das Verbreitungsgebiet setzt sich bis an das Kap ganz im Süden von Afrika fort, dabei findet sich Klee hier nur im tropischen Hochland und fehlt im Tiefland. Klee fehlt auf Madagaskar, in Namibia, Botsuana und in der Sahelzone. Die einzige Art, die den Nordrand der Sahara erreicht, ist der Filz-Klee (Trifolium tomentosum).
Die restlichen 26 % der Arten sind in Amerika verbreitet. In Nordamerika fehlt Klee in Alaska und fast ganz Kanada, in den Vereinigten Staaten fehlt er im Nordosten. Das Mannigfaltigkeitszentrum liegt in Kalifornien. Im Süden reicht das Verbreitungsgebiet etwa bis 43° südlicher Breite in Südamerika. Dabei ist der Klee auch in den Tropen Amerikas nur im Hochland zu finden und fehlt im Tiefland. Klee fehlt auf den Westindischen Inseln, in Panama und in Guayana.[9]
Zu den einheimischen Arten kommen insbesondere in Nordamerika noch viele Neophyten. Schon von den 1739 von Jan Frederik Gronovius in der Flora Virginica beschriebenen zehn Arten stellten sich neun als eingeschleppt heraus. 1994 waren in den Vereinigten Staaten 93 Kleearten verbreitet, von denen 64 natürlich dort vorkommen und 29 Neophyten sind.[10]
Auch in Australien, wo Klee natürlich nicht vorkommt, leben viele Arten neophytisch. So werden in Süd-West Australien 35 neophytisch vorkommende Kleearten gezählt.[11] Eingeschleppter Klee findet sich aber auch in den alpinen Regionen Ostaustraliens. Insgesamt zehn Arten werden als invasiv eingestuft.[12] Ebenso wurde Klee auf Neuseeland und Hawaii eingeschleppt.
Ökologie
Die Gattung Klee ist in extrem verschiedenen Habitaten heimisch. Das Verbreitungsgebiet reicht von subarktischen Regionen, durch Gebiete mit mesischen Bedingungen (Gebiete mittlerer Feuchte), über subtropische bis in tropische Regionen. Etwa 75 % der amerikanischen, 87 % der afrikanischen und 50 % der europäischen Arten sind jedoch auf gebirgige Standorte in Höhenlagen bis 4.000 Meter beschränkt.
Knöllchenbakterien
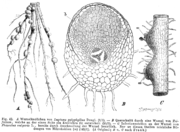 B: Querschnitt durch eine Kleewurzel, die an der Seite ein Wurzelknöllchen (b) entwickelt hat
B: Querschnitt durch eine Kleewurzel, die an der Seite ein Wurzelknöllchen (b) entwickelt hatWie auch die anderen Hülsenfrüchtler lebt Klee in einer mutualistischen Symbiose mit Knöllchenbakterien (Rhizobiaceae). Diese besitzen die Fähigkeit, elementaren, molekularen Stickstoff zu binden, also in Verbindungen zu überführen und damit biologisch verfügbar zu machen.
Anders als andere Leguminosen finden sich an Klee häufig Bakterien, die sich auf bestimmte Kleearten spezialisiert haben und mit anderen Hülsenfrüchtlern keine erfolgreiche Symbiose etablieren können. Man spricht in diesem Fall von Biovaren. Dies betrifft insbesondere Bakterien der Gattung Rhizobium. So geht zum Beispiel der Wiesen-Klee (Trifolium pratense) ausschließlich eine Symbiose mit Rhizobium trifolii ein, wohingegen zum Beispiel der Weiß-Klee (Trifolium repens) auch mit Rhizobium leguminosarum symbiotisch lebt.[13]
Bestäubung
 Schmetterling (hier Baumweißling (Aporia crataegi)) beim Nektarsaugen an einer Kleeblüte
Schmetterling (hier Baumweißling (Aporia crataegi)) beim Nektarsaugen an einer KleeblüteKlee hat typische Schmetterlingsblüten für Insektenbestäubung (Entomophilie). Dabei werden die Blüten vornehmlich von Insekten mit langem Saugrüssel besucht, wie zum Beispiel von Honigbienen (Apis), Hummeln (Bombus) und einigen Schmetterlingen (Lepidoptera). Diese sammeln dort Nektar und Pollen. Wenn die besuchenden Insekten vornehmlich mit ihrem Kopf Druck auf Fahne und Flügel ausüben, werden Narbe und Antheren aus dem Schiffchen herausgestreckt und an die Unterseite des Insektenkopfes gedrückt. So kann die Bestäubung stattfinden. Wird der Druck auf die Fahne gelöst, kehren Narbe und Antheren an ihre alte Position im Schiffchen zurück. Der Stempel ist dazu für gewöhnlich länger als die Staubblätter und leicht gebogen. Die Antheren sind aufreißend, sogar schon in der Knospe. Nektar wird an der Basis der Kronröhre sekretiert.
Eine Blüte des Weiß-Klees (Trifolium repens) produziert durchschnittlich zwischen 0,02 und 0,08 Mikroliter Nektar mit einer Zuckerkonzentration zwischen 42 % und 65 %. Bei kultivierten Arten liegt dieser Anteil im Mittel zwischen 22 % und 66 %.[14]
Die meisten Kleearten sind allogam, das heißt die Bestäubung erfolgt durch Fremdbestäubung. Die höchste Befruchtungsquote wird mit Pollen einer anderen Pflanze derselben Art erreicht (Xenogamie). Beim Wiesen-Klee (Trifolium pratense) findet die Befruchtung zwischen 18 und 50 Stunden, abhängig von der Temperatur, nach der Bestäubung statt. Für eine gute Pollenbildung ist eine relativ hohe Luftfeuchtigkeit von 93 % bis 98 % essentiell.[15] Einige Arten sind jedoch autogam (selbstbestäubend). Hier ist der Anteil der erfolgreichen Befruchtungen aber geringer. Einige wenige Arten sind sogar kleistogam, das heißt die Blüten öffnen sich während der Anthese überhaupt nicht.
Ausbreitungsmechanismen
 Detail der Blüte des Hasen-Klees (Trifolium arvense). Gut zu erkennen sind die fedrigen Auswüchse, die die Verwehung der Früchte begünstigt.
Detail der Blüte des Hasen-Klees (Trifolium arvense). Gut zu erkennen sind die fedrigen Auswüchse, die die Verwehung der Früchte begünstigt.Michael Zohary gliederte die verschiedenen Ausbreitungsmechanismen des Klees in zwei große Gruppen, Telechorie und Topochorie, die jeweils einige Unterausprägungen haben[4]:
- Als Telechorie wird die Art der Ausbreitung bezeichnet, bei der der Same zum Beispiel durch Wind oder Tiere über weite Strecken hinweg verbreitet wird. Dies ist die übliche Art der Ausbreitung innerhalb der Gattung Klee mit folgenden Ausprägungen:
- Anemochorie, die Ausbreitung durch Wind findet bei leichten Früchten statt, in einigen Fällen trägt der Kelch zusätzliche Auswüchse ähnlich wie bei einem Pappus. Diese Auswüchse können fedrig (zum Beispiel beim Hasen-Klee (Trifolium arvense)), geflügelt (zum Beispiel beim Feld-Klee (Trifolium campestre)) oder blasenförmig sein (zum Beispiel beim Persischen Klee (Trifolium resupinatum)).
Bei einigen Arten wird auch der ganze Fruchtstand verweht (zum Beispiel beim Filz-Klee (Trifolium tomentosum) oder bei Trifolium bullatum). In anderen Fällen wird die befruchtete Blüte verweht, wobei die Krone zu einem löffel- oder bootförmigen Flugapparat umgebildet ist. Hier reift der Same dann in der bereits abgetrennten Blüte. Diese Strategie findet sich zum Beispiel in der Sektion Chronosemium. - Epichorie, also die Ausbreitung durch Tiere, wird nur von wenigen Arten benutzt. Zum Beispiel verfangen sich die klettenartig umgebogenen Kelchzähne beim Kleinblütigen Klee (Trifolium retusum) oder Rauen Klee (Trifolium scabrum) gerne im Fell von Tieren.
- Anemochorie, die Ausbreitung durch Wind findet bei leichten Früchten statt, in einigen Fällen trägt der Kelch zusätzliche Auswüchse ähnlich wie bei einem Pappus. Diese Auswüchse können fedrig (zum Beispiel beim Hasen-Klee (Trifolium arvense)), geflügelt (zum Beispiel beim Feld-Klee (Trifolium campestre)) oder blasenförmig sein (zum Beispiel beim Persischen Klee (Trifolium resupinatum)).
- Amphikarpie stellt einen Sonderfall dar, der sich nur bei Trifolium polymorphum findet. Die Art bildet zwei verschiedene Früchte aus. Die oberen Blütenstände produzieren leichte Früchte, die vom Wind davongetragen werden, wohingegen die unteren Blütenstände nach unten gebogen sind und die Samen auf dem Boden ablegen. So wird Topochorie und Telechorie kombiniert.
- Topochorie bezeichnet das Phänomen, dass der Same oder die Frucht in der Nähe der Mutterpflanze verbleibt. Beim Klee finden sich davon folgende Ausprägungen:
- Barochorie heißt das schlichte Herunterfallen einzelner, nackter und schwerer Samen durch die Schwerkraft, aus einer an den Nähten aufreißenden, an der Mutterpflanze verbleibenden Frucht. Hierzu biegt sich die Hülse in der Fruchtreife nach unten. Dieser Mechanismus ist vor allem in den Untersektionen Lotoidea, Loxospermum und Falcatula verbreitet.
- Unter Calycobolie wird das Phänomen verstanden, bei dem der Kelch, der die Frucht umschließt, bei der Fruchtreife von der Rhachis abbricht und die ganze Frucht zu Boden fällt. Hier kann sie verweht werden oder bei schweren Früchten liegen bleiben. Diese Methode findet sich bei vielen Arten der Sektionen Mistyllus, Trifolium und Vesicaria.
- Bei der Synaptospermie fällt nicht eine einzelne Frucht, sondern der ganze Blütenstand mit mehreren Früchten herunter. Hierzu bricht er von der Sprossachse ab. Diese Fruchtstände sind üblicherweise zu schwer, um verweht zu werden. Synaptospermie gibt es in den Sektionen Mistyllus, und Trichocephalum, aber auch bei Arten in der Sektion Trifolium, wie zum Beispiel bei Cherlers Klee (Trifolium cherleri) oder Trifolium scutatum.
- Bei der Aestatiphorie fällt die Frucht gar nicht herab, bis die Pflanze zerfällt. Hier bleibt die Frucht geschlossen. Dieses Phänomen existiert beispielsweise beim Igel-Klee (Trifolium echinatum) oder bei Trifolium latinum.
- Basikarpie heißt der Fall, in dem die schweren Früchte den Blütenstand zu Boden drücken und dort keimen. Dies kommt unter anderem beim Schmächtigen Klee (Trifolium suffocatum) vor.
- Geokarpie schließlich findet sich bei zwei Arten der Sektion Trichocephalum. Hier werden einige sterile Blüten zu Bohrern umgebildet, die die fertilen Blüten einschließen und den Boden penetrieren. Dort reifen die Früchte heran und die Samen können keimen. Auch die Arten Trifolium batmanicum und Trifolium chlorotrichum können unterirdische Früchte bilden. Hier wird allerdings kein Bohrapparat ausgebildet, sondern die Blütenstandsachsen wachsen lang nach unten.
Parasiten und Pathogene
 Blütenstand des Kleewürgers (Orobanche minor) einem spezialisierten Kleeparasiten
Blütenstand des Kleewürgers (Orobanche minor) einem spezialisierten KleeparasitenKleearten können wie fast alle Pflanzen einer Vielzahl von Parasiten und Pathogenen als Wirt dienen. Einige Pflanzen, Tiere und Pilze haben sich aber auf Klee als Wirt spezialisiert.[16]
Die Klee-Seide (Cuscuta ephithymum ssp. trifolii) beispielsweise ist eine chlorophylllose Pflanze ohne Blätter, die auf Klee parasitiert. Die rötlichen Stängel der Klee-Seide winden sich um die Kleepflanze und dringen mit so genannten Haustorien in deren Leitungsgewebe (Phloem) ein und entziehen ihr die zum Wachstum nötige Nährlösung.
Der Kleewürger (Orobanche minor) ist eine 10 bis 40 Zentimeter hohe Pflanze, die meist auf Wiesen-Klee (Trifolium pratense) und dem Mittleren Klee (Trifolium medium) parasitiert. Erreicht die Keimwurzel des Kleewürgers eine Kleewurzel, dringt sie in diese ein und bildet wie die Klee-Seide ein Haustorium, durch das sie dem Klee die Nährstoffe entzieht.
Kleeälchen (Ditylenchus dipsaci) sind eine Art der Fadenwürmer (Nematoda), sie können unterirdisch, oder bei Überschwemmungen auch oberirdisch, in die Sprossachse junger Kleepflanzen eindringen und ernähren sich von diesen. Betroffene Pflanzenteile werden welk und fallen ab.
Beim Kleekrebs (Sclerotinia trifoliorum) handelt es sich um einen pflanzenpathogenen Pilz aus der Ordnung der Helotiales. Er befällt die Stängelbasis der Kleepflanzen und zersetzt diese.
Der Nördliche Stängelbrenner (Kabatiella caulivora) und der Südliche Stängelbrenner (Colletotrichum trifolii) sind andere auf Klee spezialisierter Pilze. Ist die Pflanze infiziert verursachen sie schwarz verfärbte Läsionen an Stängel und Laubblättern.[17]
Etymologie
Trivialname
Das deutsche Wort „Klee“ geht auf das althochdeutsche chlê zurück. Wahrscheinlich war auch die Form chlêo gebräuchlich, wobei sich das „o“ aus dem ursprünglichen „w“ der Stammeslaute (Indogermanische Ursprache) entwickelt hat. In anderen germanischen Sprachen blieb das „w“ großteils erhalten. So heißt es im Niederdeutschen klever und im Altenglischen claver, woraus sich dann das englische clover entwickelte. Auch in einigen Skandinavischen Sprachen hat sich das „w“ erhalten (dänisch: klever oder klöver, schwedisch: klöver, klövrar und norwegisch: klyver).
Aus dem Althochdeutschen entwickelte sich im Mittelhochdeutschen klê mit dem Genitiv klêwes. Eine Nebenform war klei oder cley. Eine Theorie hierzu ist, dass sich das „e“ aus „ei“ oder „ai“ entwickelt hat. So wird für die Gotische Sprache die Bezeichnung kláivs vermutet. Weitere Variationen waren ein „h“ im Genitiv und Dativ. Es wurde gesetzt um das dreifache „e“ in kleee zu vermeiden, das dann zu klehe wurde. Vor allem in Norddeutschland und um Aachen findet sich noch die weibliche Form die Klee.[18]
Wissenschaftlicher Name
Die wissenschaftliche Bezeichnung der Gattung Trifolium wurde von Carl von Linné aufgrund der vorlinnéischen Benennung einer Pflanzengruppe gewählt, unter der nicht nur die meisten Arten der heutigen Gattung, sondern auch eine ganze Reihe anderer Sippen wie zum Beispiel der heutige Fieberklee (Menyanthes trifoliata) oder viele Arten der heutigen Gattung Sauerklee (Oxalis) subsumiert wurden. Ursprung des Namens ist wohl das lateinische trifolium, das schon bei Plinius dem Älteren für Klee verwendet wird.[19] Dabei handelt es sich um eine Lehnübersetzung des altgriechischen τριφύλλον triphyllon, das von Herodot im Abschnitt 132 im ersten Buch seiner Historien, später auch von Pedanios Dioscurides erwähnt wird.[20]
Eine einleuchtende Erklärung für diese Bezeichnung wäre die Zusammensetzung aus dem lateinischen tri, das dem griechischen τρι, tri entspricht, und drei bedeutet und folium, φύλλον, phyllon was Blatt heißt. Es ist jedoch auffällig, dass eine so bekannte und in Südeuropa weit verbreitete Pflanzengattung, die auch in der Antike schon als Futterpflanze verwendet wurde, in den antiken Quellen nur spärlich erwähnt wird. Andere Arten aus derselben Tribus hingegen erscheinen deutlich häufiger in der antiken Literatur. Einige Autoren vertreten aus diesem Grund die Theorie, dass mit triphyllon tatsächlich kein Klee sondern eine andere Gattung oder Art mit dreigeteilten Blättern, wie zum Beispiel der Asphaltklee (Bituminaria bituminosa), gemeint war.[21][22]
Systematik
Bei den Arten der Sektion Lotoidea haben sich die meisten ursprünglichen Merkmale erhalten. Die Sektion gilt daher als die älteste und evolutionär primitivste Sektion der Gattung. Zohary nimmt an, dass sich alle anderen Sektionen direkt aus den Lotoidea entwickelt haben.[21]
Wo die Gattung entstanden ist, ist unklar. Jan Bevington Gillett nimmt an, dass die Gattung aus dem Mittelmeerraum stammt. Dies folge aus dem Mannigfaltigkeitszentrum in dieser Region.[23] Eine andere Theorie besagt, dass die Gattung im Hochland Ostafrikas entstanden ist und erst später nach Europa kam.[24] Eine frühe Untersuchung der Chromosomenformen stützt letztere These.[7] Zohary ist der Ansicht, dass es zwei Entstehungspunkte in Europa und Afrika gegeben haben könnte, an denen der Klee gleichzeitig entstand.[21]
Unklar ist, wie die Gattung Nordamerika erreichte. Eine Migration über die Beringstraße oder den Pazifik erscheint unwahrscheinlich, da sich im fernen Osten Russlands nur zwei Arten finden, die die Region auch nur dünn besiedeln. Eine Migrationsbewegung von Europa aus ist aber ebenfalls unwahrscheinlich, da das Mannigfaltigkeitszentrum der Gattung in Nordamerika an der Westküste liegt. Auch ist nicht geklärt, wie sich die Gattung in Amerika und Afrika über zum Teil erhebliche Lücken hinweg so weit in Nord-Süd-Richtung ausbreiten konnte.[9]
Botanische Geschichte
Die Gattung Klee war schon den antiken Naturforschern wohlbekannt. Theophrastos von Eresos glaubte sie zum Hornklee (Lotus) gehörend. Pedanios Dioscurides meinte mit seinem triphyllon den Asphaltklee (Bituminaria bituminosa).[20] Auch Plinius erwähnte die Gattung schon.[19]
 Illustration zu Trifoly in William Turners The herbal
Illustration zu Trifoly in William Turners The herbalIn der vorlinnéischen Zeit wurde Klee vor allem als Futterpflanze erwähnt. Leonhart Fuchs beschrieb und illustrierte 1543 in seinem New Kreüterbuch insgesamt sieben Kleearten, von denen vier noch heute zur Gattung zählen.[25] William Turner beschrieb 1548 in seinem Buch The names of the herbs fünf Arten unter dem Namen Trifoly, eine davon gehört aktuell noch zur Gattung.[26] John Gerard behandelte 1597 in seinem Werk The Herball or Generall Historie of Plantes sogar 21 Kleearten, von denen heute noch 10 zur Gattung gehören.[27] Klee findet sich auch bei Clusius (1601, 7 Arten[28]) und Bauhin (1620, 24 Arten[29]). Bauhin war es in seinem zweiten Buch Pinax theatri botanici 1623 auch, der zuerst versuchte, die Gattung in Untergruppen (bei ihm sind es elf) zu gliedern.[30] Sein Bruder Johann Bauhin trennte als erster 1651 zusammen mit Johann Heinrich Cherler die Gattung Schneckenklee (Medicago) klar von Trifolium ab, vermischt diesen aber immer noch mit den Gattungen Hornklee (Lotus) und Steinklee (Melilotus).[31] Weitere Autoren dieser Zeit, die den Klee abhandeln, sind Robert Morison (1680, 45 Arten[32]), Rivinus (1690–99[33]), Plukenet (1696[34]) und Tournefort (1700, 44 Arten[35]). Pier Antonio Micheli nannte 1729 die Gattung Trifoliastrum und teilte sie in sieben Gruppen.[36] Diese sieben Gruppen sind der heutigen Einteilung in Sektionen der Gattung nicht unähnlich. Jan Frederik Gronovius beschrieb 1739 in seiner Flora virginica zehn Kleearten aus Nordamerika[37], von denen aber nur eine natürlich dort vorkommt und die anderen neun bereits eingeschleppt waren. Auch Adriaan van Royen versuchte zusammen mit Albrecht von Haller eine Einteilung des Klees in Untergattungen, diese war aber weniger genau als die von Micheli.[38]
Das Hauptproblem all dieser Autoren war die saubere Trennung der Gattung Steinklee (Melilotus) vom Klee (Trifolium). Auch Carl von Linné selbst gelang dies nicht vollständig. Er schloss die Gattung Melilotus als Gruppe Trifolium-Melilotus in die Gattung Trifolium mit ein. [39]
Nach Linné fanden sich Arten der Gattung fast in jeder erschienenen lokalen Flora. Bemerkenswert ist der 1872 von Pierre Edmond Boissier verfasste Beitrag zur Flora Orientalis, in der er bereits 113 Spezies auflistete und diese in klar abgegrenzte Sektionen einteilte, die der heutigen Einteilung fast komplett entsprechen.[40]
Trotz der unzähligen Erwähnungen der Arten in der Literatur gab es nur wenige monographische Abhandlungen der Gattung. Die früheste ist von Nicolas Charles Seringe aus dem Jahr 1825, in der er 150 Arten (einschließlich einiger nordamerikanischer Arten) behandelte.[41] Im Jahr 1832 teilte Karl Bořiwog Presl die Gattung in neun eigenständige Gattungen auf.[42] Dies lehnte die botanische Gemeinschaft, mit Ausnahme Bobrows, allerdings ab. Karl Heinrich Koch gliederte wahrscheinlich in Unkenntnis von Presls Werk die Gattung in Sektionen, die mit Presls Einteilung fast übereinstimmte.[43] Ladislav Josef Čelakovský versuchte 1874 die bis dahin beschriebenen Sektionen der Gattung neu zu fassen, war jedoch eher wenig erfolgreich.[44] Der erste Schlüssel der Gattung enthielt bereits 211 Arten und wurde 1883 von Francesco Lojacono veröffentlicht.[45] Er teilte die Gattung in zwei Untergattungen, Trifoliastrum und Lagopus, was aber unnatürlich erschien und von anderen Botanikern abgelehnt wurde. Bobrow schuf 1967 analog zu Presl elf eigenständige Gattungen und begründete sogar eine eigene Tribus Lupineae für einige dieser Gattungen.[46] Diese Einteilung war aber schlecht begründet und entspricht nicht der heutigen Lehrmeinung. Michael Zohary und David Heller veröffentlichten 1984 eine große Monographie der Gattung.[21] Dieses umfangreiche Werk teilt sich in einen allgemeinen und einen taxonomischen Teil. Letzterer wurde sehr gelobt und beinhaltet 237 Arten, die dort in die heute aktuellen acht Sektionen eingeteilt werden. Der allgemeine Teil gilt jedoch als schwächer und insbesondere bei den Verbreitungsangaben als fehlerbehaftet.[9] John Montague Gillett veröffentlichte 2001 zusammen mit Norman L. Taylor eine weitere Monographie der Gattung.[47] Diese verfügt nur über einen sehr rudimentären allgemeinen Teil und enthält keine taxonomische Einordnung der Arten, vielmehr werden 228 Arten illustriert und beschrieben. Neben diesen Monographien über die Gattung wurde eine Vielzahl von Monographien zu einzelnen Sektionen veröffentlicht.
Äußere Systematik
 Die Gattung der Steinklees (Melilotus), hier Kleinblütiger Steinklee (Melilotus indicus), ist eng mit dem Klee verwandt.
Die Gattung der Steinklees (Melilotus), hier Kleinblütiger Steinklee (Melilotus indicus), ist eng mit dem Klee verwandt.Die Gattung Klee ist Teil der Familie der Hülsenfrüchtler (Fabaceae) in der Unterfamilie der Schmetterlingsblütler (Faboideae). Sie wird der Tribus Trifolieae und der Subtribus Trifoliinae zugeordnet.
Die Tribus Trifolieae enthält fünf oder sechs Gattungen und teilt sich in drei Subtribus. Die Gattungen, die neben dem Klee (Trifolium) sicher zu den Trifolieae zählen, sind Trigonella, Schneckenklee (Medicago) und Steinklee (Melilotus). Nicht sicher ist der Status der Gattungen Parochetus und Hauhechel (Ononis), die eine Sonderrolle spielen.[48] Zohary zählte noch die Gattung Factorovskya zur Tribus, diese wurde aber 1984 in die Gattung Schneckenklee (Medicago) überführt.[49]
Nach morphologischen Gesichtspunkten werden die Gattungen Trigonella, Medicago und Melilotus in die eine Subtribus Trigonellinae eingeordnet, wohingegen Trifolium als einzige Gattung in die Subtribus Trifoliinae zählt. Für die unklare Gattung Parochetus wurde eine eigene Subtribus Parochetinae geschaffen, wohingegen Ononis in eine eigene Tribus Ononideae gestellt wurde.[50]
Neuere molekulargenetische Untersuchungen zeigen jedoch ein leicht anderes Bild.[51] Der enge Zusammenhang der Tribus Trigonellinae wird zunächst bestätigt. Medicago ist monophyletisch und Schwestertaxon zu Trigonella. Melilotus hingegen ist mit Trigonella verschachtelt (paraphyletisch). Nach dieser Untersuchung ist Trifolium aber näher mit der Tribus der Fabeae (früher unter dem Namen Vicieae bekannt[52]), bestehend aus den Wicken (Vicia), den Linsen (Lens), den Erbsen (Pisum), den Platterbsen (Lathyrus) und der Gattung Vavilovia, verwandt, als mit den Trigonellinae. Ebenso ist die Gattung Ononis näher mit den Trigonellinae verwandt als mit Trifolium, wohingegen Parochetus sehr weit von diesen Arten entfernt ist.[53] Eine noch neuere Untersuchung bestätigt diese Ergebnisse.[54] Danach ergäbe sich folgendes Kladogramm:
┌───────── Fabeae ┌──┤ │ └───────── Klee (Trifolium) ─┤ │ ┌──────── Hauhecheln (Ononis) └────┤ ┌───── Schneckenklee (Medicago) └────┤ └───── Steinklee (Melilotus) / TrigonellaInnere Systematik
Nach der monographischen Bearbeitung der Gattung durch Michael Zohary und David Heller 1984 umfasst sie 237 Arten, sieben Arten wurden seitdem neu erstbeschrieben, einer der Artstatus aberkannt, so dass der Gattung gegenwärtig 243 Arten zugerechnet werden. Zohary teilt die Gattung nach morphologischen Kennzeichen in acht Sektionen ein:
- Die Sektion Lotoidea ist mit 99 Arten in Amerika, Afrika und Eurasien die größte Sektion der Gattung und die taxonomisch schwierigste. Die Arten sind so heterogen, dass die Sektion in neun Untersektionen und 13 Serien aufgeteilt wurde. Vor allem die Einteilung der amerikanischen Arten ist schwierig. Hauptkennzeichen der Sektion sind: Schirmartiger Blütenstand, gestielte Blüten mit Tragblättern und zwei- bis vielsamige Hülsenfrüchte. Die Sektion gilt als die evolutionär primitivste Sektion der Gattung.
- Die Sektion Paramesus mit nur zwei Arten in Eurasien ist umstritten. Einige Autoren schlagen vor, sie in die Sektion Lotoidea einzuschließen. Merkmale sind die drüsigen Zähne auf den Nebenblättern und am Kelch.
- Die Sektion Mistyllus mit neun Arten in Afrika und Eurasien ist durch die für Klee einmalige Struktur des symmetrischen, blasenförmigen Kelches und der persistenten Krone klar von den anderen Sektionen unterschieden.
- Auch die Sektion Vesicaria mit sieben Arten in Eurasien ist durch die für Klee einmalige Struktur des asymmetrischen, blasenförmigen Kelches klar von den anderen Sektionen unterschieden. Auch die auf dem Kopf stehenden Blüten finden sich exklusiv in dieser Sektion.
- Die Sektion Chronosemium mit 17 Arten in Eurasien ist klar von allen anderen Sektionen unterschieden. Merkmale sind der auffällige zweilippige Kelch, die persistente Krone mit löffel- oder bootförmiger Fahne und die einsamigen Früchte.
- Die Sektion Trifolium ist mit 74 Arten in Südafrika und Eurasien die zweitgrößte Sektion der Gattung. Sie teilt sich in 17 relativ kleine aber natürliche Gruppen, die als Untersektionen beschrieben werden. Die Blüten sind hier sitzend und ohne Tragblätter. Die Kronröhre ist oft behaart und die Früchte sind einsamig ohne Nähte.
- Die Sektion Trichocephalum mit neun Arten in Eurasien unterscheidet sich stark von allen anderen Arten durch den Fakt, dass die meisten Blüten steril sind und zu Haaren oder Borsten für die Samenverbreitung umgebildet werden.
- Die Sektion Involucrarium mit 26 Arten nur in Amerika ist von den anderen Sektionen durch die Gestalt der Kelchzähne, die Zähne an den Nebenblättern und die ausgeprägten Hüllblätter abgegrenzt.
Eine molekulargenetische Untersuchung aus dem Jahr 2006 ergab, dass wahrscheinlich eine Einteilung in zwei Untergattungen Chronosemium und Trifolium mit acht Sektionen gerechtfertigt ist. In diesem Fall würde die Sektion Lotoidea zerrissen und die Sektionen Mistyllus und Vesicaria zusammengefasst. Bislang wurden die nach dieser Untersuchung vorgeschlagenen Untergattungen aber noch nicht gültig erstbeschrieben.[55]
Die Arten der Gattung Trifolium und ihre Zuordnung nach Zohary und Heller sind:
Innere Systematik und Arten der Gattung Klee (Trifolium) Westalpen-Klee (Trifolium alpinum) – Habitus
Westalpen-Klee (Trifolium alpinum) – Habitus Blütenstände des Berg-Klees (Trifolium montanum)
Blütenstände des Berg-Klees (Trifolium montanum) Braun-Klee (Trifolium badium) – Blütenstände
Braun-Klee (Trifolium badium) – Blütenstände Blütenstand des Stern-Klees (Trifolium stellatum)
Blütenstand des Stern-Klees (Trifolium stellatum) Stein-Klee (Trifolium saxatile) – Blütenstand
Stein-Klee (Trifolium saxatile) – Blütenstand Bodenfrüchtiger Klee (Trifolium subterraneum) – Illustration
Bodenfrüchtiger Klee (Trifolium subterraneum) – Illustration- Sektion Lotoidea Crantz
- Untersektion Falcatula (Brot.) Aschers. et Graebn.
-
- Vogelfuß-Klee (T. ornithopodioides (L.) Sm.)
-
- Untersektion Loxospermum (Hochst.) Čelak.
-
- T. chilaloense Thulin
- T. decorum Chiov.
- T. elgonense Gillett
- T. multinerve A.Rich.
- T. schimperi A.Rich
-
- Untersektion Lupinaster (Adans.) Belli
-
- T. andersonii A.Gray
- T. friscanum (S.L.Welsh) S.L.Welsh[56]
- T. gordejevi (Kom.) Z.Wei[57]
- T. gymnocarpon Nutt.
- T. lemmonii S.Wats.
- T. lupinaster L.
- T. macrocephalum (Pursh) Poir.
- T. polyphyllum C.A.Mey.
- T. thompsonii Morton
-
- Untersektion Ochreata (Lojac.) Gillett
-
- T. cheranganiense Gillett
- T. cryptopodium A.Rich.
- T. mauginianum Fiori
- T. polystachyum Fresen.
- T. simense Fresen.
- T. somalense Taub.
- T. stolzii Harms
- T. ukingense Harms
- Usambara-Klee (T. usambarense Taub.)
- T. wentzelianum Harms
-
- Untersektion Lotoidea
- Serie Grandiflora Heller et Zoh.
- Westalpen-Klee (T. alpinum L.)
- T. attenuatum Greene
- T. eximium Steph. ex Ser.
- T. nanum Torr.
- T. pilczii Adamović
- Einblütiger Klee (T. uniflorum L.)
- Serie Phyllodon Heller et Zoh.
- T. bejariense Moric.
- T. carolinianum Michx.
- Serie Brachyantha Heller et Zoh.
- Kleinblütiger Klee (T. retusum L.)
- Serie Lotoidea
- T. amabile Humb., Bonpl. et Kunth
- T. angulatum Wladst. et Kit.
- T. bifidum A.Gray
- T. bivonae Guss.
- T. breweri S.Wats.
- T. burchellianum Ser.
- T. calcaricum J.L.Collins et Wieboldt[58]
- T. cernuum Brot.
- T. euxinum Zoh.
- T. gillettianum Jac.-Fél.
- T. gracilentum Torr. et A.Gray
- Schweden-Klee (T. hybridum L.)
- T. latifolium (Hook) Greene
- T. leibergii Nels. et Macbride
- T. masaiënse Gillett
- Michelis-Klee (T. michelianum Savi)
- Schwarzwerdender Klee (T. nigrescens Viv.)
- Bleicher Klee (T. pallescens Schreb.)
- T. parnassi Boiss. et Hohen.
- T. purseglovei Gillett
- T. radicosum Boiss. et Hohen.
- T. reflexum L.
- Weiß-Klee (T. repens L.)
- T. semipilosum Fresen.
- T. stoloniferum Muhlenberg
- Rasiger Klee (T. thalii Vill.)
- T. virginicum Small
- Serie Pectinata Heller et Zoh.
- T. ciliolatum Benth.
- Serie Curvicalyx Heller et Zoh.
- T. eriocephalum Nutt.
- Serie Producta Heller et Zoh.
- T. beckwithii Brewer
- T. bolanderi A.Gray
- T. brandegei S.Wats.
- T. haydenii Porter
- T. howellii S. Wats.
- T. kingii S.Wats.
- Serie Acaulia (Baker) Heller et Zoh.
- T. acaule A.Rich.
- T. petitianum A.Rich.
- T. vestitum Heller et Zoh.
- Serie Grandiflora Heller et Zoh.
- Untersektion Oxalioidea (Gillett) Heller
-
- T. polymorphum Poir.
-
- Untersektion Platystylium Willk.
- Serie Platystylium
- T. abyssinicum Heller
- T. africanum Ser.
- Kaukasischer Klee (T. ambiguum M.Bieb.)
- T. baccarinii Chiov.
- T. bilineatum Fresen.
- Schmalfrüchtiger Klee (T. isthmocarpum Brot.)
- T. lanceolatum (Gilett) Gillett
- T. longipes Nutt.
- Berg-Klee (T. montanum L.)
- T. owyheense Gilkey
- T. pichisermollii Gillett
- T. rueppellianum Fresen.
- T. spananthum Thulin
- T. tembense Fresen.
- Serie Macrochlamis Heller et Zoh.
- T. dasyphyllum Torr. et A.Gray
- T. parryi A.Gray.
- T. riograndense Burkart
- Serie Phyllocephala Heller et Zoh.
- T. andinum Nutt.
- Serie Micrantheum (C.Presl.) Heller et Zoh.
- Knäuel-Klee (T. glomeratum L.)
- T. pachycalyx Zoh.
- Schmächtiger Klee (T. suffocatum L.)
- Serie Altissima (Abrams) Heller
- T. douglasii House
- Serie Platystylium
- Untersektion Calycospatha (Chiov.) Heller et Zoh.
-
- T. mattirolianum Chiov.
- T. pseudostriatum Baker, f.
-
- Untersektion Neolagopus Lojac..
-
- T. albopurpureum Torr. et A.Gray
- T. dichotomum Hook et Arn.
- T. macraei Hook et Arn.
- T. plumosum Dougl. ex Hook
-
- Untersektion Falcatula (Brot.) Aschers. et Graebn.
- Sektion Paramesus (C.Presl.) Endl.
-
-
- T. glanduliferum Boiss.
- Steifer Klee (T. strictum L.)
-
-
- Sektion Mistyllus (C.Presl.) Godr.
-
-
- T. aintabense Boiss. et Hausskn.
- T. argutum Sol.
- T. lugardii Bullock
- T. mutabile Portenschl.
- Schaum-Klee (T. spumosum L.)
- T. steudneri Schweinf.
- Blasenfrüchtiger Klee (T. vesiculosum Savi)
- T. quartinianum A.Rich.
-
-
- Sektion Vesicaria Crantz
-
-
- T. bullatum Boiss. et Hausskn.
- T. clusii Godr. et Gren.
- Erdbeer-Klee (T. fragiferum L.)
- T. physodes Stev. ex M.B.
- Persischer Klee (T. resupinatum L.)
- Filz-Klee (T. tomentosum L.)
- T. tumens Stev. ex M.B.
-
-
- Sektion Chronosemium Ser.
-
- Serie Stipata Zoh.
- T. sintenisii Freyn
- Serie Comosa Gib. et Belli
- T. billardieri Spreng.
- T. boissieri Guss.
- T. brutium Ten.
- T. grandiflorum Schreb.
- T. philistaeum Zoh.
- Serie Badia Bobr.
- Braun-Klee (T. badium Schreb.)
- T. erubescens Fenzl
- T. sebastianii Savi
- Moor-Klee (T. spadiceum L.)
- Serie Agraria Bobr.
- Gold-Klee (T. aureum Poll.)
- Feld-Klee (T. campestre Schreb.)
- Spreiz-Klee (T. patens Schreb.)
- T. velenovskyi Vandas
- Serie Filiformia Gib. et Belli
- T. dolopium Heldr. et Hausskn.
- Faden-Klee (T. dubium Sibth.)
- Kleinster Klee (T. micranthum Viv.)
- Serie Stipata Zoh.
-
- Sektion Trifolium
- Untersektion Trifolium
-
- T. andricum Lassen[59]
- Ausgebreiteter Klee (T. diffusum Ehrh.)
- T. mazanderanicum Rech.f.
- Norischer Klee (T. noricum Wulf.)
- Bleicher Klee (T. pallidum Waldst. et Kit.)
- Wiesen-Klee (T. pratense L.)
-
- Untersektion Intermedia (Gib. et Belli) Bobrov
-
- T. heldreichianum (Gib. et Belli) Hausskn.
- Mittlerer Klee (T. medium L.)
- T. patulum Tausch
- Pignants Klee (T. pignantii Brogn. et Bory)
- T. velebiticum Deg.
- T. wettsteinii Dörfl. et Hay.
-
- Untersektion Ochroleuca Gib. et Belli
-
- Grau-Klee (T. canescens Willd.)
- T. caucasicum Tausch
- T. caudatum Boiss.
- T. davisii Hossain
- T. longidentatum Náb.
- Blassgelber Klee (T. ochroleucum Huds.)
- Ungarischer Klee (T. pannonicum Jacq.)
- T. trichocephalum M.B.
-
- Untersektion Alpestria Gib. et Belli
-
- Hügel-Klee (T. alpestre L.)
- Purpur-Klee (T. rubens L.)
-
- Untersektion Stellata Gib. et Belli
-
- Inkarnat-Klee (T. incarnatum L.)
- Stern-Klee (T. stellatum L.)
- Wald-Klee (T. sylvaticum Gérard ex Loisel.)
-
- Untersektion Stenosemium (Čelak.) Aschers. et Graebn.
-
- Streifen-Klee (T. striatum L.)
-
- Untersektion Trichoptera Gib. et Belli
-
- T. bocconei Savi
- T. trichopterum Panč.
-
- Untersektion Scabroidea Gib. et Belli
-
- T. dalmaticum Vis.
- T. lucanicum Gasp. ex Guss.
- Rauer Klee (T. scabrum L.)
-
- Untersektion Phleoidea Gib. et Belli
-
- T. gemellum Pourr. ex Willd.
- Ligurischer Klee (T. ligusticum Balb. ex Loisel.)
- Lieschgras-Klee (T. phleoides Pourr. ex Willd.)
-
- Untersektion Lappacea Gib. et Belli
-
- T. barbeyi Gib. et Belli
- Cherlers Klee (T. cherleri L.)
- T. congestum Guss.
- Rosen-Klee (T. hirtum All.)
- Kletten-Klee (T. lappaceum L.)
-
- Untersektion Arvensia Gib. et Belli
-
- T. affine C.Presl.
- Hasen-Klee (T. arvense L.)
- Stein-Klee (T. saxatile All.)
- T. stipulaceum Thunb.
-
- Untersektion Angustifolia Gib. et Belli
-
- Schmalblättriger Klee (T. angustifolium L.)
- T. blancheanum Boiss.
- T. dichroanthoides Rech.f.
- T. dasyurum C.Presl.
- T. dichroanthum Boiss.
- T. haussknechtii Boiss.
- T. infamia-ponertii Greuter[60]
- T. palaestinum Boiss.
- T. prophetarum Hossain
- T. purpureum Loisel.
- T. roussaeanum Boiss.
-
- Untersektion Alexandrina Zoh.
-
- Ägyptischer Klee (T. alexandrinum L.)
- T. apertum Bobrov
- T. berytheum Boiss. et Bl.
- T. meironense Zoh. et Lern.
- T. salmoneum Mout.
- T. vavilovii Eig
-
- Untersektion Squamosa Zoh.
-
- T. cinctum DC.
- Schuppen-Klee (T. squamosum L.)
-
- Untersektion Urceolata Zoh.
-
- T. constantinopolitanum Ser.
- T. daveauanum Thell.
- Juliens-Klee (T. juliani Batt.)
- T. leucanthum M.B.
- T. miegeanum Maire
- T. obscurum Savi
- Sparriger Klee (T. squarrosum L.)
-
- Untersektion Echinata Zoh.
-
- Igel-Klee (T. echinatum M.Bieb.)
- T. latinum Sebast.
-
- Untersektion Clypeata Gib. et Belli
-
- Schild-Klee (T. clypeatum L.)
- T. plebeium Boiss.
- T. scutatum Boiss.
-
- Untersektion Trifolium
- Sektion Trichocephalum Koch.
-
-
- T. batmanicum Katzn.
- T. chlorotrichum Boiss. et Bal.
- T. eriosphaerum Boiss.
- Kugeliger Klee (T. globosum L.)
- T. israëliticum D.Zoh. et Katzn.
- T. meduseum Bl. ex Boiss.
- T. pauciflorum D’Urv.
- T. pilulare Boiss.
- Bodenfrüchtiger Klee (T. subterraneum L.)
-
-
- Sektion Involucrarium Hook.
- Untersektion Involucrarium
-
- T. antucoensis Heller
- T. buckwestiorum Isely[61]
- T. chilense Hook. et Arn.
- T. microcephalum Pursh
- T. microdon Hook. et Arn.
- T. monanthum A.Gray
- T. mucronatum Willd. ex Spreng.
- T. obtusiflorum Hook.f.
- T. oliganthum Steud.
- T. pinetorum Greene
- T. polyodon Greene
- T. siskiyouense J.M.Gillett
- T. triaristatum Bert. ex Colla
- T. trichocalyx Heller
- T. tridentatum Lindl.
- T. variegatum Nutt.
- T. vernum Phil.
- T. wigginsii Gillett
- T. wormskioldii Lehm.
-
- Untersektion Physosemium (Lojac.) Heller et Zoh.
-
- T. barbigerum Torr.
- T. cyathiferum Lindl.
- T. depauperatum Desv.
- T. fucatum Lindl.
- T. jokerstii Vincent & Rand.Morgan [10]
- T. minutissimum Heller et Zoh.
- T. physanthum Hook. et Arn.
-
- Untersektion Involucrarium
Gefährdung
Im Anhang I des Washingtoner Artenschutz-Übereinkommens sowie in der Roten Liste gefährdeter Arten der IUCN sind keine Kleearten enthalten. Allerdings sind einige Arten durch Standortverluste gefährdet, so listet die Fauna-Flora-Habitat-Richtlinie (FFH) der Europäischen Union Stein-Klee (Trifolium saxatile) als gefährdet.[62]
Mehrere Kleearten sind in den Roten Listen der einzelnen deutschen Bundesländer gelistet, in der Anlage 1 zur Bundesartenschutzverordnung sind aber keine Kleearten enthalten. Die Rote Liste der Schweiz listet den Streifen-Klee (Trifolium striatum) als „stark gefährdet“ (endangered) und fünf Arten (Trifolium spadiceum, saxatile, patens, ochroleucon und fragiferum) sowie einige Unterarten als „gefährdet“ (vulnerable).[63] In Österreich gilt Trifolium retusum als vom Aussterben bedroht, Trifolium spadiceum und Trifolium striatum als stark gefährdet[64].
Der Status einzelner Kleearten in anderen Regionen ist schwer einzuschätzen, für die meisten Gebiete sind keine Gefährdungen bekannt.
Nutzung
 Kleefeld in Montana (1917)
Kleefeld in Montana (1917)Die ersten Überlieferungen über Kleeanbau betreffen Vorderasien und stammen aus der Zeit des frühen Christentums. Der hebräische Name für Klee תלתן, tiltan taucht zwar auch bereits in der Mischna auf, es ist jedoch nicht sicher, ob hier wirklich exklusiv Pflanzen der Gattung Trifolium gemeint sind. In Europa wurde zunächst Wiesen-Klee (Trifolium pratense) ab dem 4. Jahrhundert nach Christus angebaut.
Klee wurde auch schon von den kalifornischen Indianerstämmen gesammelt und siliert. Die Silage wurde dann auch von den Menschen gegessen.[65]
Im Jahr 1953 wurden 15 Kleearten kommerziell angebaut[66], 2001 waren es 16, dabei werden in Nordamerika und Europa die gleichen Arten kultiviert.[47] Sie werden vor allem als Futtermittel eingesetzt und können sowohl als Grünfutter genutzt werden, als auch siliert[67] oder zu Kleeheu getrocknet werden. Klee wird vor allem wegen seines hohen Anteils an verdaulichem Eiweiß verfüttert. Wegen der Fähigkeit, in Symbiose mit Knöllchenbakterien Stickstoff aus der Luft zu binden, dient er auch zur Erhaltung der Bodenfruchtbarkeit.
Seltener wird Klee auch als Bienentrachtpflanze und in sehr kleinem Maßstab als Küchenkraut und zu medizinischen Zwecken angebaut. So wirken verschiedene Kleearten, wie beispielsweise Faden-Klee (Trifolium dubium) blutungsstillend, andere Arten können durch die enthaltenen Phytoöstrogene bei Menstruationsbeschwerden helfen oder nutzen zur Steigerung der Milchmenge beim Stillen.[68] Wiesen-Klee (Trifolium pratense) soll auch bei Hautkrankheiten helfen und wird in der traditionellen chinesischen Medizin zur Behandlung von Krebsleiden eingesetzt.[69]
Anbau
Kleeanbau ist in ganz Europa und Nordamerika weit verbreitet. Er erstreckt sich aber auch um das ganze Mittelmeer inklusive der afrikanischen Anrainerstaaten bis nach Asien. Auch in Australien und Neuseeland findet Kleeanbau in beträchtlichem Maße statt. In Zentralafrika werden vor allem andere Leguminosen und kaum Klee angebaut.[70]
Klee kann in Hauptfrucht als Reinsaat ohne Partner, oder zusammen mit Gräsern als Deckfrucht (Kleegras, häufig auch mit Winterroggen), angebaut werden. Zudem ist Kleeanabau in Winterzwischenfrucht allein oder im Gemisch als Untersaat möglich. Als Hauptfrucht mit oder ohne Kleegras werden im Mittel Erträge von 250 Dezitonnen Grünmasse pro Hektar erzielt, unter optimalen Bedingungen und bei intensiver Düngung sind aber Erträge bis 500 dt/ha Grünmasse möglich. Beim Anbau als Winterzwischenfrucht beträgt der mittlere Grünmasseertrag 135 bis 190 dt/ha.[71]
Am Umfang der Anbaufläche und an der Bedeutung für die Fütterung gemessen, ist der Wiesen-Klee (Trifolium pratense) die wichtigste Kleeart. Auf Grünland wird häufig Weiß-Klee (Trifolium repens) mit eingesät, wo er den Nährstoffgehalt des Futters deutlich verbessert. Auf leichten Böden wird er häufig mit Knäuelgräsern (Dactylis) kombiniert.
Weitere für die Landwirtschaft bedeutende Kleearten sind Ägyptischer Klee (Trifolium alexandrinum), Persischer Klee (Trifolium resupinatum), Schweden-Klee (Trifolium hybridum) und Inkarnat-Klee (Trifolium incarnatum), letzterer vor allem als Teil des Landsberger Gemenges.
Zur Ertragssteigerung wurden auch Kreuzungen zwischen verschiedenen Arten geschaffen und getestet. Erfolgreich und mittlerweile ökonomisch bedeutend ist eine Hybride Trifolium repens × nigrescens aus Weiß-Klee (Trifolium repens) und Schwarzwerdendem Klee (Trifolium nigrescens).[72] Ebenfalls angebaut wird Trifolium repens × ambiguum aus Weiß-Klee und Kaukasischem Klee (Trifolium ambiguum).[73].
Klee als Giftpflanze
Kleearten sind wertvolle Futterpflanzen, dennoch können sie in Abhängigkeit von der Jahreszeit, der Witterung und der verfütterten Menge zu Schäden bei Tieren führen. Rinder sind durch Klee-Arten vor allem bei übermäßiger Fütterung oder bei nassem Klee gefährdet.
Viele Kleearten können durch die in ihnen enthaltenen Thiocyanate (anorganische Salze) bei Tieren phototoxische Reaktionen hervorrufen und zu einer Überempfindlichkeit gegenüber Licht führen. Diese Erkrankung wird Trifoliose oder Kleekrankheit genannt. Sie äußert sich in geröteten und geschwollenen Augenlidern und Lippen, auch andere ungeschützte Hautpartien können betroffen sein. Trifoliose tritt vor allem bei Pferden, Rindern und Schafen auf.[51]
Bei Rindern kann Klee darüber hinaus durch schaumige Gärung löslicher, cytoplasmatischer Proteine zu Gasansammlungen im Pansen (Pansentympanie) führen. In Klee enthaltene Cyanogene Glykoside, vor allem Linamarin und Lotaustralin können zur Blockierung der Atmungskette und zu Krämpfen führen. Phytoöstrogene, vor allem verschiedene Isoflavone, aus Klee können zu Fruchtbarkeitsstörungen führen.[74]
Kulturgeschichte
 Klee im Triptychon des Willem Moreel von Hans Memling (Ausschnitt vergrößert)
Klee im Triptychon des Willem Moreel von Hans Memling (Ausschnitt vergrößert) Klee auf Haupttafel des Altarretabels zum Leben Marias, Szene: Die Verkündigung (Jesuitenkirche, Cortona)
Klee auf Haupttafel des Altarretabels zum Leben Marias, Szene: Die Verkündigung (Jesuitenkirche, Cortona) Die ganze Welt in einem Kleeblatt, Karte aus Magdeburg 1581, Im Zentrum liegt Jerusalem
Die ganze Welt in einem Kleeblatt, Karte aus Magdeburg 1581, Im Zentrum liegt Jerusalem Ogata Kenzan: Pflaumen, Nelke, Klee und Schnee (1742), auch hier repräsentiert der Klee den Sommer
Ogata Kenzan: Pflaumen, Nelke, Klee und Schnee (1742), auch hier repräsentiert der Klee den Sommer Pyxis mit Dekor aus vierblättrigen Kleeblättern, erste Hälfte 13. Jahrhundert
Pyxis mit Dekor aus vierblättrigen Kleeblättern, erste Hälfte 13. Jahrhundert Fensterrosette mit mehreren Drei- und Vierpässen in der Santa Maria del Pi in Barcelona
Fensterrosette mit mehreren Drei- und Vierpässen in der Santa Maria del Pi in Barcelona Wappen von Fürth
Wappen von FürthDas Kleeblatt ist ein sehr altes Symbol, das schon im alten Perserreich Bedeutung hatte. Die Symbolik des Kleeblatts ist vielfältig.
Religiöse Bedeutungen
Das dreifiedrige Kleeblatt ist seit jeher ein Symbol für Dreiheit. Im Perserreich der Spätantike steht es für die Götterdreiheit (Trias) von Mithra (Mihr) neben Ahuramazda (Ohrmazd) und Anahita (Anahid).[75] Bei den vorchristlichen keltischen Druiden galt der Klee als heilige Symbol- und Zauberpflanze.
In der christlichen Symbolik wurde das Kleeblatt schon früh als Zeichen der Dreifaltigkeit übernommen und findet sich häufig in bildlichen Darstellungen. Es wurde zum Symbol des heiligen Patrick von Irland, der angeblich den Iren anhand eines Kleeblatts die Dreieinigkeit erklärte. Ein anderer Erklärungsversuch aber leitet das Symbol vom Kreuzstab mit seinem kleeblattartigen oberen Ende, den St. Patrick einer Schlange in den Rachen stieß. In Irland wird das Kleeblatt Shamrock genannt. Die ältesten noch erhaltenen Manuskripte, die es noch als sheamair oder sheamrach erwähnen stammen aus dem Jahr 1571.[76]Noch heute ist es Symbol des irisch-keltischen Nationalbewusstseins.[77][78][79]
Als Heilpflanze wurde der Klee im Mittelalter Maria, der Mutter Gottes, zugeordnet. Da Klee im Mittelalter wegen seines frischen Grüns häufig auf Gräber gepflanzt wurde, wohl auch als Hinweis auf ein Leben nach der Auferstehung, wurde es auch zum Symbol des Abschieds. [80] So lässt beispielsweise Johann Lenz in seiner Beschreibung des Schwabenkriegs die Armee vor der blutigen Schlacht zuerst durch den „grünen Klee“ ziehen:
„durch loub gras, durch den grünen cley
in stiller zucht on alles geschrey“– Johann Lenz: Der Schwabenkrieg[81]
Klee als Liebessymbol
Im Zusammenhang mit Rosen, dem Symbol der Liebe, und mit Veilchen, wegen der violetten Bußfarbe, wird der Klee auch als Liebessymbol verwendet. Zum Beispiel trägt Isolde bei ihrer Liebesbegegnung mit Tristan in Gottfried von Straßburgs Tristan und Isolde einen Schapel (reifenförmiger Kopfschmuck) aus Klee.
„Ein Schapel trug sie von Klee.
Ihrem Herren mehr denn je
Schien sie reizend, wonniglich.“– Gottfried von Straßburg: Tristan und Isolde[82]
Im Neuen Teutschen Parnass von Johann Rist steht Klee im Zusammenhang mit der Redewendung „Singen von der schönen Galathée“. Einer alten Legende nach verliebt sich ein Künstler in seine selbst geschaffene Statue der Nymphe Galathée, was sie zu einem Symbol für die Liebe machte.
„da zehlen sie die schaf' in ihrer gelben klee
und singen bald darauf ein lied von galathe“– Johann Rist: Neuer Teutscher Parnass[83]
Bedeutender in der mittelalterlichen Liebeslyrik ist der grüne Klee als Ort der Liebesbegegnung und des Stelldicheins. In diesem Zusammenhang wird er etwa in Der grünenden Jugend überflüssige Gedanken von Christian Weise verwendet. Klee ist als Symbol in dieser Bedeutung aber auch bei vielen anderen Dichtern, vor allem nach dem 16. Jahrhundert, beliebt.[80]
„und durften unterschiedlich im klee beisammen ruhn“
– Christian Weise: Der grünenden Jugend überflüssige Gedanken[84]
Lob und Übertreibung
Die weit verbreitete Redewendung „jemanden über den grünen Klee loben“ wird in der Bedeutung, den Gelobten in übertriebener Weise über die Gebühr zu rühmen, verwendet. Ursprünglich bedeutete diese Redewendung, dass dem Gelobten eine größere Lebenskraft zugeschrieben wurde.[77] In der Liebeslyrik und in Märchen des Mittelalters war das Bild des grünen Klees als Inbegriff des Frischen und Lebensvollen weit verbreitet und findet sich beispielsweise im Grimmschen Märchen Die vier kunstreichen Brüder (KHM 129).
„'Ja,' sprach der Alte zu seinen Söhnen, 'ich muß euch über den grünen Klee loben, ihr habt eure Zeit wohl benutzt und was Rechtschaffenes gelernt“
– Brüder Grimm: Die vier kunstreichen Brüder[85]
Die heutige Redensart bezieht sich wahrscheinlich auf diesen dichterischen Lobpreis des Klees im Mittelalter, der in späteren Zeiten als übertrieben erschien, so dass etwas was noch darüber hinausging als groteske Steigerung aufgefasst werden musste.[80]
Klee als Symbol des Frühlings und des Sommers
Schon in der chinesischen Symbolik ist das Kleeblatt ein Zeichen des Sommers.[75] Auch in unserem Kulturkreis wird es wegen seiner kräftigen grünen Farbe gerne als Frühlings- oder Sommersymbol verwendet, so zum Beispiel auch von Ewald Christian von Kleist in seinem Gedicht Der Frühling:
„Ein Teppich geschmückt mit Ranken und Laubwerk,
Von Büschen, Blumen und Klee, wallt auf Gefilden und Auen“– Ewald Christian von Kleist: Der Frühling[86]
In Volksliedern werden, vielleicht wegen des glücklichen Reims von „Schnee, Klee und weh“, Schnee und Klee gerne als Gegensatzpaar verwendet. Der Schnee bedeutet dann Winter und Leid, wohingegen der Klee für Sommer und Liebe steht.[80] Beispiele sind Es war eine Mutter, die hatte vier Kinder oder Ade zur guten Nacht:
„Im Sommer da wächst der Klee,
Im Winter, da schneit´s den Schnee,
Da komm ich wieder“– Volkslied aus Mitteldeutschland um 1850: Ade zur guten Nacht[87]
Das vierblättrige Kleeblatt
In der christlichen Symbolik repräsentiert das Vierblatt ursprünglich das Kreuz und die vier Evangelien.[88] Von Alters her gilt es aber besonders als Glückszeichen. Dies ist wohl nicht nur auf seine Seltenheit zurückzuführen, sondern ursprünglich auf den kräftig-vitalen Wuchs dieser Pflanzen.[78] Eine Legende besagt, dass Eva bei der Vertreibung aus dem Paradies ein vierblättriges Kleeblatt mitnahm und es so ein Stück des Paradieses verkörpert.[89]
Bereits bei den Druiden in keltischer Zeit galt das vierblättrige Kleeblatt als ein wirksames Mittel gegen böse Geister.[90] John Melton persifliert den Aberglauben an das Vierblatt bereits in seiner Satire Astrologaster. Wahrscheinlich ist dies die erste schriftliche Erwähnung von vierblättrigem Klee als Glückssymbol:[91]
“If a man walking in the fields find any four-leaved grass, he shall in a small while after find some good thing.”
– John Melton: Astrologaster[92]
„Findet ein Mann beim Spazieren durch die Felder irgendein vierblättriges Gras, dann wird er kurz darauf auch etwas Gutes finden.“
– frei übersetzt
Neben vierblättigen Laubblättern treten auch, wenngleich noch seltener, Blätter mit mehr als vier Fiedern auf. Laut Guinness-Buch der Rekorde lag 2008 der Rekord bei einem 18-blättrigen Kleeblatt.[93] Echte vierblättrige Kleeblätter werden zumeist an Weiß-Klee (Trifolium repens) gefunden. Zu ihrer Häufigkeit gibt es keine gesicherten Untersuchungen. Sammler berichten aber von einer Quote von etwa eins zu 10.000.[94] Eine in den Jahren 1979 bis 1986 in München durchgeführte Untersuchung legt ein Verhältnis zugunsten der dreiblättigen zwischen 20 zu eins und 60 zu eins nahe. [95] Die Art Glücksklee (Oxalis tetraphylla), die sich gelegentlich im Handel findet, gehört zur Gattung der Sauerklees (Oxalis) und ist kein echter Klee.
Kunst
Die von Kleeblättern abgeleiteten Ornamente: Drei- und Vierpass waren in der Gotik weit verbreitet. Dort wurden sie vor allem als Fensterformen eingesetzt, finden sich aber auch auf Münzen. Der deutsche Maler und Expressionist Paul Klee signierte seine Werke mit einem dreiblättrigen Kleeblatt und seinen Initialen K. P.
Heraldik
Auch in der Heraldik sind Abbildungen von Kleeblättern beliebt. Es kommt sowohl das vierblättrige, als auch das dreiblättrige Kleeblatt als Motiv vor. Beispiele für die Verwendung der dreiblättrigen Variante sind das Wappen der Gemeinde Cleebronn, der Gemeinde Hitzhofen oder das des ehemaligen Berliner Bezirks Tiergarten in Deutschland, Viktorsberg, Kirchbach, Angerberg oder Schönwies in Österreich und Berikon oder Löhningen in der Schweiz. Kleeblätter gibt es aber auch in den Wappen verschiedener Adelsgeschlechter, wie zum Beispiel Carlowitz oder Bodewitz.[96]
Sehr bekannt sind die Wappen der Städte Fürth und Kassel. Beim Fürther Wappen ist die Herkunft des dreiblättrigen Kleeblatts allerdings nicht genau geklärt, vermutlich geht die Dreiheit jedoch auf die Dreiherrschaft der Dompropstei Bamberg, der Markgrafschaft Ansbach sowie der Reichsstadt Nürnberg über Fürth während des Mittelalters zurück. Das Kleeblatt wurde auch im Vereinswappen der Spielvereinigung Greuther Fürth übernommen. Auch im Kasseler Wappen ist die Bedeutung der 13 Kleeblätter im Wappen umstritten. Das Wappen von Hannover zeigt eine Marienblume, und nicht, wie häufig fälschlicherweise behauptet, ein Kleeblatt. Beispiele für die Verwendung der vierblättrigen Variante sind das Wappen der Gemeinde Brest und der Gemeinde Brinkum sowie die Wappen von SC Rot-Weiß Oberhausen und des schottischen Vereins Celtic Glasgow.
Sonstiges
Eine Bauart von Autobahnkreuzen wird wegen seiner Aufsicht als Kleeblatt bezeichnet. Es ist die in Deutschland am weitesten verbreitete Form des Autobahnkreuzes. Sie besteht aus je vier Tangential- (Rampen) und Radialfahrbahnen, den so genannten Ohren.
Das Trefoil (englisch: Klee-, Dreiblatt) genannte Symbol für radioaktive und ionisierende Strahlung erinnert an ein Kleeblatt. Es wurde 1946 an der University of California in Berkeley entworfen.
Beim Französischen Kartenspiel heißt eine Spielfarbe trefle, was zu deutsch Kleeblatt bedeutet. In Deutschland wird diese Farbe häufig Kreuz genannt. Im Deutschen Blatt entspricht sie den Eicheln.
Der Wiesen-Klee (Trifolium pratense) ist seit 1895 die Staatsblume des US-Bundesstaats Vermont.
Literatur
- Michael Zohary, David Heller: The Genus Trifolium. The Israel Academy of Sciences and Humanities, Jerusalem 1984, ISBN 978-965-208-056-1.
- John M. Gillett, Norman L. Taylor, M. Gillett: The World of Clovers. Iowa State University Press, Ames 2001, ISBN 978-0-8138-2986-9.
- Asmus Petersen; Waltraud Petersen (Hrsg.): Klee und Kleeartige als Kulturpflanzen, Wildpflanzen und Unkräuter auf Acker, Wiese und Weide. 2. Auflage. Akademie Verlag, Berlin 1967.
Einzelnachweise
- ↑ M. T Abberton, T. P. T. Michaelson-Yeates, A. H. Marshall, K. Holdbrook-Smith, I. Rhodes: Morphological characteristics of hybrids between white clover, Trifolium repens L., and Caucasian clover, Trifolium ambiguum M. Bieb. In: Plant breeding. 117, Nr. 5, 1998, ISSN 0179-9541, S. 494–496 (abstract).
- ↑ J. R. Crush, D. A. Care, A. Gourdin, D. R. Woodfield: Root growth media effects on root morphology and architecture in white clover. In: New Zealand Journal of Agricultural Research. 48, Nr. 2, 2005, ISSN 0028-8233, S. 255–263 (abstract).
- ↑ A. W. Humphries and E. T. Baily: Root weight profiles of eight species of Trifolium grown in swards. In: Australian Journal of Experimental Agriculture and Animal Husbandry. 1, Nr. 3, November 1961, S. 150–152 (doi:10.1071/EA9610150).
- ↑ a b Michal Zohary: Origins and Evolution in the Genus Trifolium. In: Botaniska Notiser. 125, Nr. 4, 1972, S. 501–511.
- ↑ Peter Leins: Blüte und Frucht. Morphologie, Entwicklungsgeschichte, Phylogenie, Funktion, Ökologie. E. Schweizerbart'sche Verlagsbuchhandlung, Stuttgart 2000, ISBN 978-3-510-65194-8, S. 58.
- ↑ J. B. Morris, S. L. Greene: Defining a Multiple-Use Germplasm Collection for the Genus Trifolium. In: Crop Science. 41, 2001, S. 893–901 (volltext).
- ↑ a b A. J. Pritchard: Number and morphology of chromosomes in African species in the genus Trifolium L. In: Australian Journal of Agricultural Research. 13, Nr. 6, 1962, S. 441–445 (abstract).
- ↑ Chen Chi-Chang, P. B. Gibson: Karyotypes of Fifteen Trifolium Species in Section Amoria. In: Crop Science. 11, 1971, S. 1023–1029.
- ↑ a b c J. B. Gillett: The Genus Trifolium by Michael Zohary and David Heller. In: Kew Bulletin. 42, Nr. 1, 1987, S. 280–281 (abstract).
- ↑ a b Michael A. Vincent, Randall Morgan: Trifolium jokerstii (Leguminosae, Papilionoideae), a New Species from Butte County, California. In: Novon. 8, Nr. 1, 1998, S. 91–93 (abstract).
- ↑ Leslie Watson: Trifolium. In: FloraBase: the Western Australian Flora. 2008. Abgerufen am 17. September 2008.
- ↑ Bruce Auld, Hirohiko Morita, Tomoko Nishida, Misako Ito1, Peter Michael: Shared exotica: Plant invasions of Japan and south eastern Australia. In: Cunninghamia. 8, Nr. 1, 2003, S. 147–152 (pdf).
- ↑ Herman P. Spaink, Carel A. Wijffelman, Elly Pees, Rob J. H. Okker, B. J. J. Lugtenberg: Rhizobium nodulation gene nodD as a determinant of host specificity. In: Nature. 328, 23. Juli 1987, S. 337–340 (doi:10.1038/328337a0).
- ↑ Nevin Weaver: Foraging behavior of honeybees on white clover. In: Insectes Sociaux. 12, Nr. 3, September 1965, ISSN 0020-1812, S. 231–240 (doi:10.1007/BF02223893).
- ↑ A. J. Pritchard, L. t'Mannetje: The breeding systems and some interspecific relations of a number of African Trifolium spp.. In: Euphytica. 16, Nr. 3, Dezember 1967, ISSN 0014-2336, S. 324–329 (doi:10.1007/BF00028938).
- ↑ W. Diepenbrock, G. Fischbeck, K.-U. Heyland, N. Knauer: Spezieller Pflanzenbau. 3. Auflage. Ulmer, Stuttgart 1999, ISBN 978-3-8252-0111-1, S. 347.
- ↑ Herbert Cole Jr., Houston B. Couch: Cytological Investigations of Kabatiella caulivora. In: American Journal of Botany. 46, Nr. 1, Januar 1959, S. 12-16 (abstract).
- ↑ Jacob und Wilhelm Grimm: Deutsches Wörterbuch. 11, dtv, München 1999, S. 1059–1062.
- ↑ a b Plinius der Ältere, naturalis historia 21,54.
- ↑ a b Etwa bei De materia medica 4,17: Hasenklee; 4,111 (113): Schneckenklee.
- ↑ a b c d Michael Zohary, David Heller: The Genus Trifolium. The Israel Academy of Sciences and Humanities, Jerusalem 1984, ISBN 978-965-208-056-1.
- ↑ Helmut Genaust: Etymologisches Wörterbuch der botanischen Pflanzennamen. 3. Auflage. Nikol, Hamburg 2005, ISBN 978-3-937872-16-2, S. 656f.
- ↑ Jan Bevington Gillett: The Genus Trifolium in Southern Arabia and in Africa South of the Sahara. In: Kew Bulletin. 7, S. 367–404.
- ↑ D. O. Norris: Legumes and Rhizobium Symbiosis. In: Empire Journal of Experimental Agriculture. 24, 1956, S. 247–270.
- ↑ Leonhart Fuchs: New Kreüterbuch. Vma-Vertriebsgesellschaft, Wiesbaden 2002, ISBN 978-3-928127-84-4 (online ; Stand: 22. August 2008).
- ↑ William Turner: The herbal. 2. Teil, Köln 1568, S. 158ff (online ; Stand: 25. August 2008).
- ↑ John Gerarde: The Herball or Generall Historie of Plantes. London 1597 (online ; Stand: 25. August 2008).
- ↑ Charles de L'Ecluse: Rariorum plantarum historia. Antwerpen 1602 (online ; Stand: 25. August 2008).
- ↑ Caspar Bauhin: Prodromos. Basel 1620 (online ; Stand: 25. August 2008).
- ↑ Caspar Bauhin: Pinax theatri botanici. Basel 1623 (online ; Stand: 25. August 2008).
- ↑ Johann Bauhin, Johann Heinrich Cherler: Historiæ plantarum generalis novæ et absolutæ. Mömpelgard, heute Montbéliard 1623.
- ↑ Robert Morison: Historia Plantarum Universalis Oxoniensis. Oxford 1680 (online ; Stand: 25. August 2008).
- ↑ August Quirinus Rivinus: Ordo plantarum. Leipzig 1690–99.
- ↑ Leonard Plukenet: Almagestum botanicum sive Phytographiae Pluc'netianae Onomasticon Methodo Syntheticâ digestum. London 1696.
- ↑ Joseph Pitton de Tournefort: Institutiones rei herbariae, editio altera. Paris 1700.
- ↑ Pier Antonio Micheli: Nova plantarum genera. Florenz 1729 (online ; Stand: 25. August 2008).
- ↑ Jan Frederik Gronovius: Flora virginica. Leiden 1739 (online ; Stand: 25. August 2008).
- ↑ Adriaan van Royen: Flora Leydensis Prodomus. Leiden 1740 (online ; Stand: 25. August 2008).
- ↑ Carl von Linné: Species Plantarum. Band 2, Stockholm 1753, S. 764ff (online ; Stand: 25. August 2008).
- ↑ Pierre Edmond Boissier: Trifolium. In: Flora Orientalis. 2, Genf, Basel 1872, S. 110–156.
- ↑ Nicolas Charles Seringe: Trifolium. In: A. P. de Candolle (Hrsg.): Prodromus Systematis Naturalis Regni vegetabilis. 2, Paris 1825, S. 189–207.
- ↑ Karl Bořiwog Presl: Trifolium. In: Symbolae Botanicae. 1, Prag 1832, S. 44–50.
- ↑ Karl Heinrich Koch: Trifolium. In: Synopsis Florae Germanicae et Helveticae. 1, Wilmans, Frankfurt 1835.
- ↑ Ladislav Čelakovsky: Ueber den Aufbau der Gattung Trifolium. In: Oesterreichische Botanische Zeitschrift. 24, 1874, S. 37–45 und 75–82.
- ↑ Francesco Lojacono: Clavis Specierum Trifoiorum. In: Nuovo Giornale Botanico Italiano. 15, 1883, S. 225–278.
- ↑ Jewgeni Grigorjewitsch Bobrow: Ob ob'eme roda Trifolium. In: Botaniceskjij Žurnal SSSR. 52, Nr. 11, 1967, S. 1593–1599.
- ↑ a b John M. Gillett, Norman L. Taylor, M. Gillett: The World of Clovers. Iowa State University Press, Ames 2001, ISBN 978-0-8138-2986-9.
- ↑ Ernest Small, M. Jomphe: A synopsis of the genus Medicago (Leguminosae). In: Canadian Journal of Botany. 67, 1989, S. 3260–3294.
- ↑ Ernest Small, Brenda S. Brookes: Reduction of the Geocarpic Factorovskya to Medicago. In: Taxon. 33, Nr. 4, November 1984, S. 622–635 (abstract).
- ↑ L. B. Chaudhary, M. Sanjappa: Parochetinae: A New Subtribe of Trifolieae (Leguminosae, Papilionoideae). In: Taxon. 47, Nr. 4, November 1998, S. 829–831 (abstract).
- ↑ a b Gwilym Lewis, Brian Schrire, Barbara Mackinder, Mike Lock: Legumes of the World. Royal Botanic Gardens, Kew 2005, ISBN 978-1-900347-80-8, S. 499ff.
- ↑ W. Greuter, J. McNeill, F. R. Barrie, H. M. Burdet, V. Demoulin, T. S. Filgueiras, D. H. Nicolson, P. C. Silva, J. E. Skog, P. Trehane, N. J. Turland, D. L. Hawksworth (Hrsg.): International Code of botanical nomenclature (Saint Luis Code). In: Regnum Vegetable. Koeltz Scientific Books, Königstein 2000, ISBN 978-3-904144-22-3, S. 19.4 und 18.5.
- ↑ K. P. Steele, M. F. Wojciechowski: Phylogenetic analyses of tribes Trifolieae and Vicieae, based on sequences of the plastid gene, matK (Papilionoideae: Leguminosae). In: B. Klitgaard,A. Bruneau (Hrsg.): Advances in legume systematics. Part 10, Higher level systematics. Kew: Royal Botanic Gardens, Kew 2003, ISBN 978-1-84246-054-2, S. 355–370 (pdf).
- ↑ Martin F. Wojciechowski, Matt Lavin, Michael J. Sanderson: A phylogeny of legumes (Leguminosae) based on analysis of the plastid matK gene resolves many well-supported subclades within the family. In: American Journal of Botany. 91, 2004, S. 1846–1862 (abstract).
- ↑ Nick W. Ellison, Aaron Liston, Jeffrey J. Steiner, Norman L. Taylor: Molecular phylogenetics of the clover genus (Trifolium—Leguminosae). In: Molecular Phylogenetics and Evolution. 39, Nr. 3, Juni 2006, S. 688–705 (doi:10.1016/j.ympev.2006.01.004).
- ↑ S. L. Welsh: New taxa and new nomenclatural combinations in the Utah flora. In: Rhodora. 95, Nr. 883/884, 1993, S. 392–421.
- ↑ Ernest Small: Reduction of Ursia to Trifolium. In: Taxon. 36, Nr. 3, August 1987, S. 578–583 (abstract).
- ↑ Joseph L. Collins, Thomas F. Wieboldt: Trifolium calcaricum (Fabaceae), a New Clover from Limestone Barrens of Eastern United States. In: Castanea. 57, Nr. 4, Dezember 1992, S. 282–286 (abstract).
- ↑ P. Lassen: Trifolium andricum (Fabaceae), a new species from Greece. In: Annalen des Naturhistorischen Museums in Wien. 98 B Suppl, November 1996, S. 301–301 (pdf).
- ↑ Werner Greuter: The flora of Psara (E. Aegean Islands, Greece). An annotated catalogue. In: Candollea. 31, 1976, S. 215.
- ↑ D. Isely: Innovations in California Trifolium and Lathyrus. In: Madroño. 39, Nr. 2, 1992, ISSN 0024-9637, S. 90–97 (abstract).
- ↑ Richtlinie 92/43/EWG des Rates vom 21. Mai 1992 zur Erhaltung der natürlichen Lebensräume sowie der wildlebenden Tiere und Pflanzen. 22. Juli 1992, S. 41 (pdf ; Zuletzt geändert am 20. Dezember 2006).
- ↑ BAFU: Rote Listen: Farne und Blütenpflanzen. Bundesamt für Umwelt BAFU. Abgerufen am 22. September 2008.
- ↑ Harald Niklfeld: Rote Liste gefährdeter Pflanzen Österreichs. 2. Auflage. Grüne Reihe des Bundesministeriums für Umwelt, Jugend und Familie, 1999, ISBN 978-3-85333-028-9.
- ↑ M. A. Wittrock, G. L. Wittrock: Food plants of the Indians: vegetables, grains, fruits, seasonings gleaned from the wild by the North American natives. In: Journal of the New York Botanical Garden. 43, 1942, S. 57–86.
- ↑ F. J. Hermann: A Botanical Synopsis of Cultivated Clovers. In: United States Department of Agriculture Monographs. 22, 1953, S. 45ff.
- ↑ A. H. Marshall, M. T. Abberton, T. A. Williams, T. P. T. Michaelson-Yeates , H. G. Powell: Forage quality of Trifolium repens L. × T. nigrescens Viv. hybrids. In: Grass and Forage Science. 58, Nr. 3, 2003, S. 295–301 (doi:10.1046/j.1365-2494.2003.00381.x).
- ↑ Daniel E. Moerman: Native American Ethnobotany. Timber Press, Portland 1998, ISBN 978-0-88192-453-4, S. 566f (google books ; Stand: 26. September 2008).
- ↑ James A. Duke, Edward S. Ayensu: Medicinal Plants of China. Reference Publications, Washington, DC 1984, ISBN 978-0-917256-20-2.
- ↑ R. P. Barrett: Legume species as leaf vegetables. In: J. Janick, J. E. Simon (Hrsg.): Advances in new crops: Proceedings of the First National Symposium New Crops, Research, Development, Economics, Indianapolis, Indiana, October 23–26. Timber Press, Portland 1990, ISBN 978-0-88192-166-3, S. 391–396 (online ; Stand: 24. September 2008).
- ↑ Heinz Wetterau, Siegfried Jahn, Gotthard Leisner: Kleines Futtermittel abc. Deutscher Landwirtschaftsverlag, Berlin 1968, S. 140–144.
- ↑ Athole Marshall, Terry Michaelson-Yeates, Michael Abberton, Ian Rhodes: New Clovers for the New Millennium. In: Iger Innovations. 1997, S. 29–33 (pdf).
- ↑ M. T. Abberton: Interspecific hybridization in the genus Trifolium. In: Plant Breeding. 126, Nr. 4, 2007, S. 337–342 (doi:10.1111/j.1439-0523.2007.01374.x).
- ↑ Franziska Ritter: Giftpflanzen: Übersicht über giftige Riedpflanzen zur Erkennung von Problempflanzen für die landwirtschaftliche Nutzung. Greifensee-Stiftung, 2008, S. 19f (pdf).
- ↑ a b J. C. Cooper: Lexikom alter Symbole. E. A. Seemann, Leipzig 1986, ISBN 978-3-363-00000-9, S. 93.
- ↑ Roy Vickery: Shamrock: Botany and History of an Irish Myth by E. Charles Nelson. In: Folklore. 103, 1992, S. 128 (abstract).
- ↑ a b Hans Biedermann: Knauers Lexikon der Symbole. Knauer, München 1989, ISBN 978-3-426-26400-3, S. 237.
- ↑ a b Hildegard Kretschmer: Lexikon der Symbole und Attribute in der Kunst. Reclam, Stuttgart 2008, ISBN 978-3-15-010652-5, S. 219.
- ↑ E. Charles Nelson: Shamrock: Botany and History of an Irish Myth. Boethius Press, Aberystwyth 1991, ISBN 978-0-86314-200-0.
- ↑ a b c d Lutz Röhrich: Lexikon der sprichwörtlichen Redensarten. 6. Auflage. 2, Herder, Freiburg 2003, ISBN 978-3-451-05400-6, S. 852f.
- ↑ Johann Lenz; H. von Dießbach (Hrsg.): Der Schwabenkrieg, Besungen von einem Zeitgenossen. von Drell, Zürich 1849, S. 152 (online ; Stand: 12. September 2008).
- ↑ Gottfried von Straßburg: Tristan und Isolde. F. M. Brockhaus, Leipzig 1855 (übersetzt von Karl Simrock), Kapitel 29 (online ; Stand: 12. September 2008).
- ↑ Johann Rist: Neuer Teutscher Parnass. Georg Olms Verlag, Hildesheim 1978, ISBN 978-3-487-06673-8, S. 694 (Nachdruck der Ausgabe Lüneburg 1652).
- ↑ Christian Weise: Der grünenden Jugend überflüssige Gedanken. Niemeyer, Halle 1914, S. 440 (Abdruck der Ausgabe von 1678).
- ↑ Brüder Grimm: Kinder- und Hausmärchen. Vollständige Ausgabe. 19. Auflage. Artemis & Winkler Verlag, Düsseldorf 1999, ISBN 978-3-538-06943-5, S. 608–611 (wikisource ; Stand: 15. September 2008).
- ↑ Ewald Christian von Kleist: Der Frühling. Heidegger und Compagnie, Zürich 1750, S. 8 (wikisource ; Stand: 12. September 2008).
- ↑ Ade zur guten Nacht. In: Volksliederarchiv. Abgerufen am 15. September 2008.
- ↑ Peter W. Hartmann: Kunstlexikon. Beyars, Neumarkt 1996, ISBN 978-3-9500612-0-8 (online ; Stand: 16. September 2008).
- ↑ Otto Craemer: Zur Psychopathologie der religiösen Wahnbildung. In: European Archives of Psychiatry and Clinical Neuroscience. 53, Nr. 1, Dezember 1913, ISSN 0940-1334, S. 275–301 (doi:10.1007/BF01987365).
- ↑ Norman L. Taylor: Clover Science and Technology. American Society of Agronomy, Madison Mai 1985, ISBN 978-0-89118-083-8.
- ↑ David Beaulieu: Irish Shamrocks and 4-Leaf Clovers. In: about.com. Abgerufen am 16. September 2008.
- ↑ John Melton: Astrologaster, Or, the Figvre-Caster. AMS Press, Brooklyn, 1995, ISBN 978-0-404-70173-4 (Nachdruck der Ausgabe von 1620).
- ↑ Guinness World Records 2009. 1. Auflage. Bibliographisches Institut, Mannheim 2008, ISBN 978-3-411-14079-4 (online ; Stand: 16. September 2008).
- ↑ Mary Jordan: A Little Irish Luck Goes a Long Way. In: The Washington Post. 17. März 2005, S. A15 (online ; Stand: 16. September 2008).
- ↑ Maria C. und Olav Röhrer-Ertl: Was bedeutet die Aussage "Klee ist dreiblättrig?". In: Globulus. Beiträge der Natur- und kulturwissenschaftlichen Gesellschaft e.V.. 4, Polyglon-Verlag, Buxheim, Eichstätt 1996, ISBN 3-928671-16-2, ISSN 0944-0321, S. 70-72. , S.71
- ↑ Ottfried Neubecker: Heraldik. Orbis, Luzern 1990, ISBN 978-3-572-01344-9, S. 134.
Weblinks
- Als Telechorie wird die Art der Ausbreitung bezeichnet, bei der der Same zum Beispiel durch Wind oder Tiere über weite Strecken hinweg verbreitet wird. Dies ist die übliche Art der Ausbreitung innerhalb der Gattung Klee mit folgenden Ausprägungen:



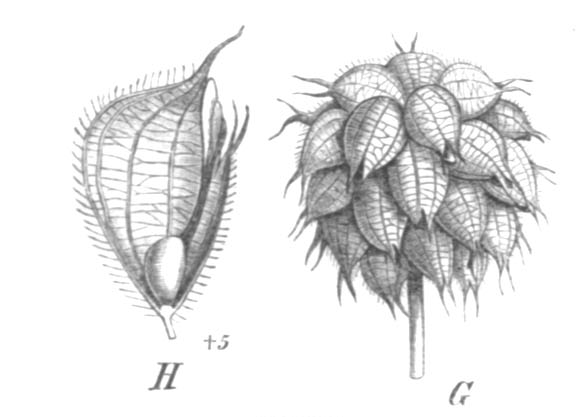

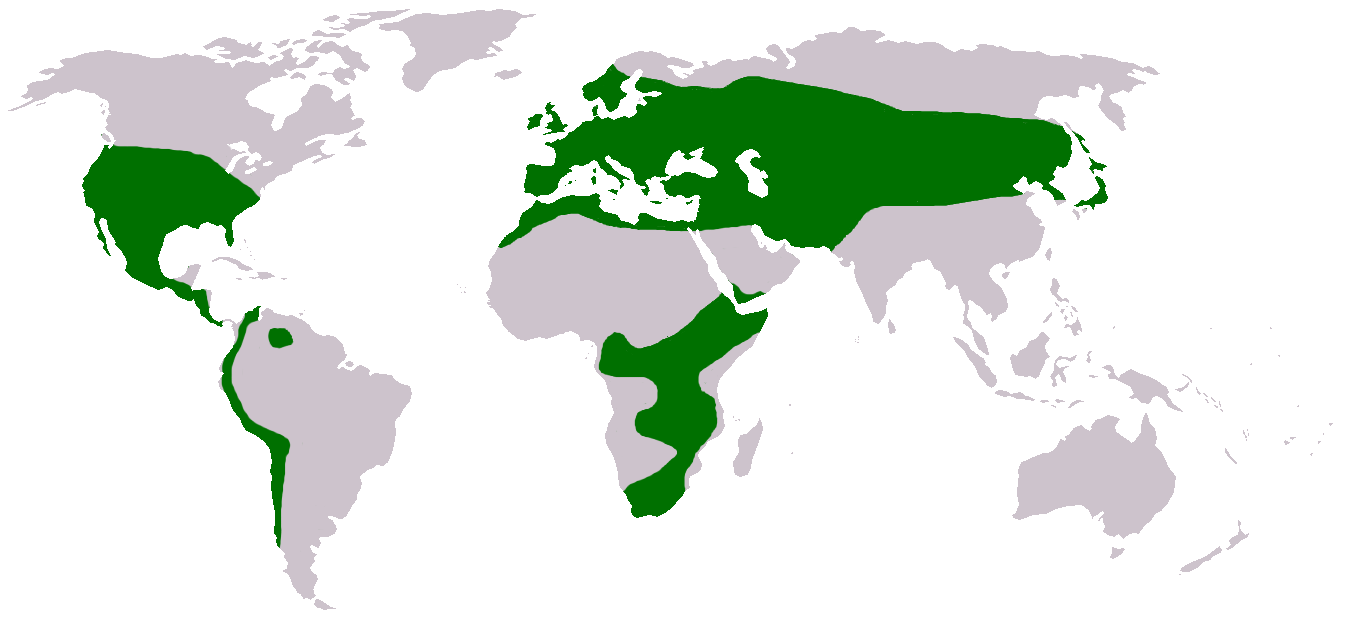
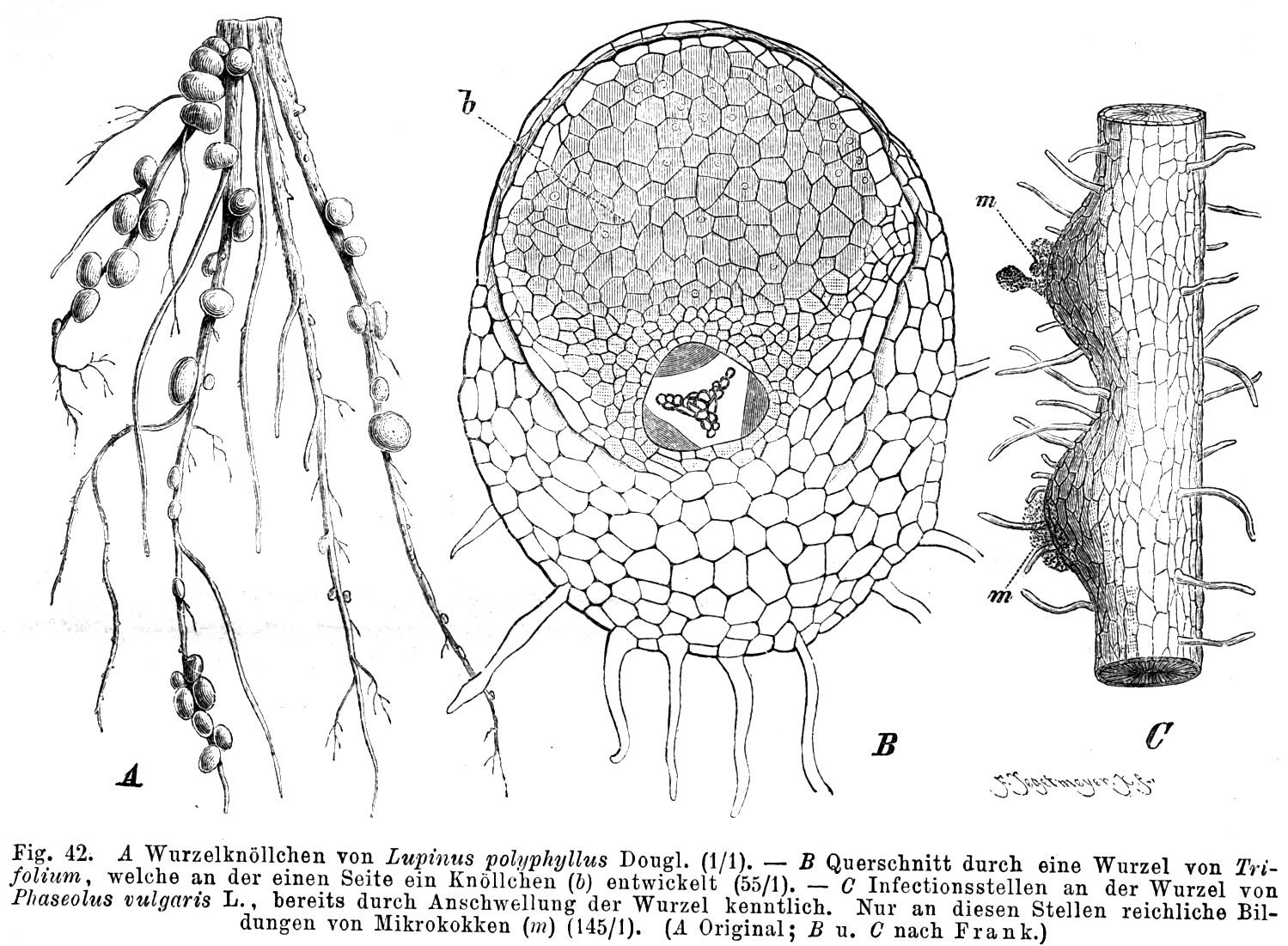













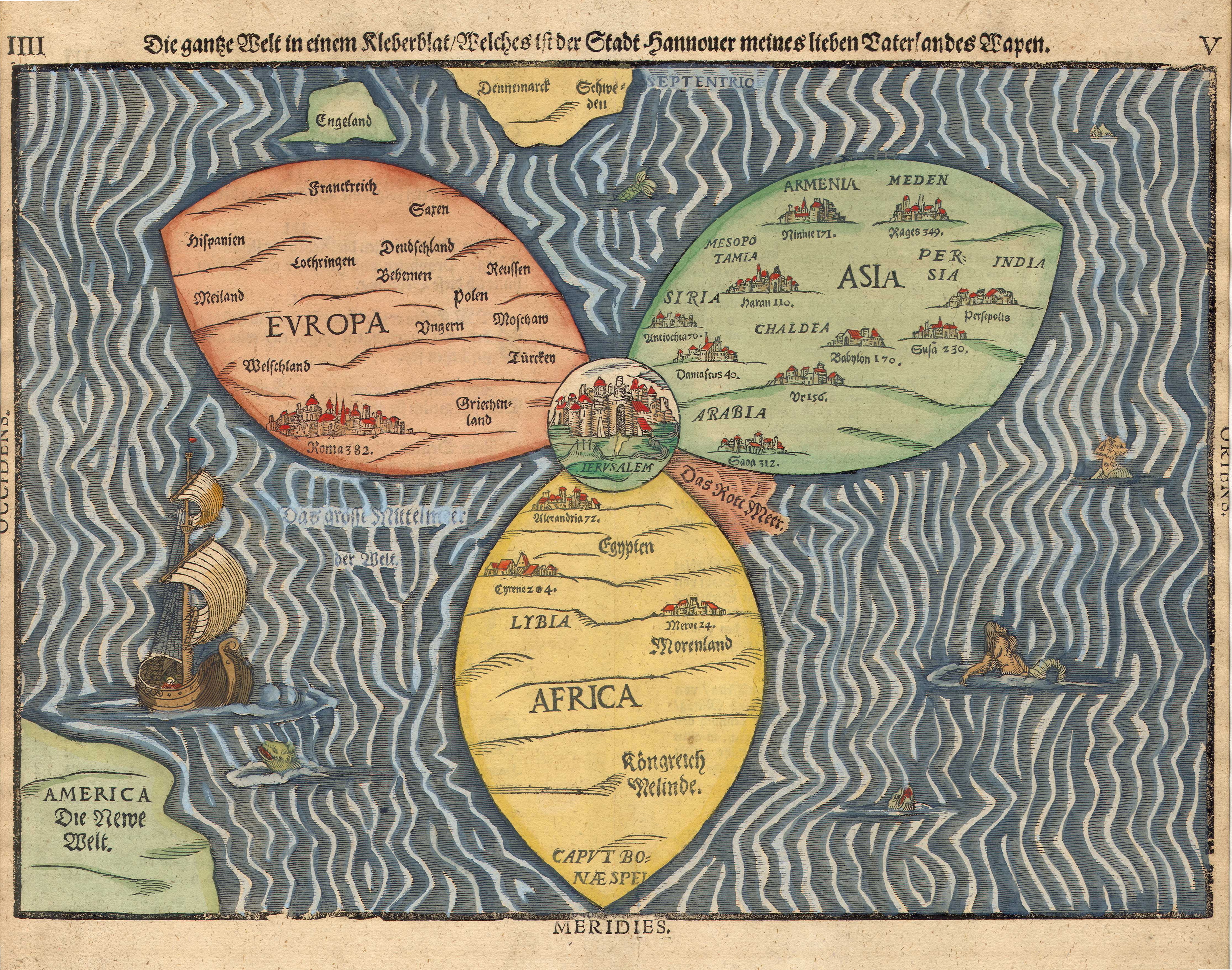





Wikimedia Foundation.