- Wandteichonsäure
-
Teichonsäuren sind polymere Bausteine der Zellwand gram-positiver Bakterien und können 20 bis 40 % deren Zellwand-Trockenmasse ausmachen. Sie sind meist kovalent in der Peptidoglycanschicht (oder Mureinschicht) verankert. Teichonsäuren bestehen entweder aus Ribitol-Phosphat-Polymeren (Ribitol-Teichonsäuren) oder aus Glycerol-Phosphat-Polymeren (Glycerol-Teichonsäuren), die kettenartig nach außen ragen.
Als Lipoteichonsäuren bezeichnet man Glycerol-Teichonsäuren, die kovalent an Glycolipide der Plasmamembran gekoppelt sind. Diese sind damit in der Zytoplasmamembran verankert und durchdringen die gesamte Peptidoglycanschicht.
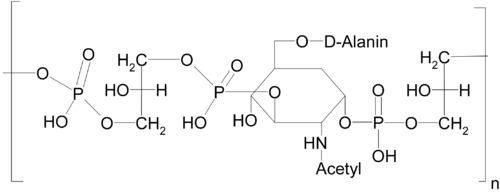 Strukturbeispiel einer Teichonsäure aus Micrococcusarten
Strukturbeispiel einer Teichonsäure aus MicrococcusartenDie Teichonsäuren üben bei der Interaktion der Bakterien- mit der Wirtszelle wichtige Funktionen aus, etwa bei der Adhäsion und als sogenannte Virulenzfaktoren.[1]
Inhaltsverzeichnis
Lipoteichonsäuren
Lipoteichonsäuren (LTA) kommen hauptsächlich in grampositiven Eubakterien vor, bei denen der Guanin- und Cytosin-Gehalt (GC-Gehalt) der DNA weniger als 50 % beträgt. In grampositiven Bakterien mit mehr als 50 % GC-Gehalt findet man meist Lipoglycane anstelle von Lipoteichonsäuren.[2] Lipoteichonsäuren bestehen aus einer hydrophilen Kette aus Alditolphosphaten und einem amphiphilen Glycolipid, das als Membrananker fungiert.[3][4] Bei Staphylococcus aureus findet man Polyglycerolphosphat-LTA. Dieser WTA-Typ ist am weitesten verbreitet und wurde auch bei Gattungen der Arten Bacillus, Enterococcus, Lactobacillus, Lactococcus, Leuconostoc, Listeria und Streptococcus gefunden. Die Polyglycerolphosphatkette der LTA von S. aureus ist aus 16–40 1,3-verknüpften Glycerolphosphateinheiten aufgebaut und über eine Phosphodiesterbindung an das Glycolipid gebunden.[2] Die Glycerolphosphatreste der meisten LTA sind teilweise mit D-Alaninestern modifiziert, und in vielen Bakterien findet man auch Glycosylsubstituenten.[2] Auch bei S. aureus finden sich D-Alaninester und Glycosylsubstituenten.
Die LTA scheint wichtige Funktionen in der Physiologie der Zellwand auszuüben. So wurden verschiedene Funktionen vorgeschlagen, wie beispielsweise die Bindung von Magnesiumionen, Adhäsion an Wirtszellen oder die Regulation von Autolysinen.[5][6][2] Die Biosynthese der LTA scheint ein Transmembranprozess zu sein, da alle Bestandteile Lipidcharakter haben. Die Zusammensetzung der LTA beginnt mit einem Glycerolphospattransfer von Phosphatidylglycerol auf das Glycolipid, welches als Lipidanker dient, wobei Phosphatidylglycerol auch als Glycerolphosphatspender zur Kettenverlängerung dient. Zusätzlich sind möglicherweise noch zwei Glycerolphosphat-Transferasen beteiligt. Eine, die das Glycolipid erkennt und eine zweite die für die Kettenverlängerung zuständig ist. Für jedes Glycerolphosphat, welches übertragen wird, entsteht an der inneren Membran ein Diacylglycerol, das wieder zu Phosphatidylglycerol recycelt wird. Ein Teil des Diacylglycerols wird zur Glycolipidsynthese verwendet.[7][8]
Wandteichonsäuren
Wandteichonsäuren (WTA) sind im Peptidoglycan lokalisiert. Strukturell sind sie gegenüber den LTA komplexer aufgebaut. Die Ankerstruktur, welche über eine Phosphodiesterbindung an den Sauerstoff der N-Acetylmuraminsäure des Peptidoglycans gebunden ist, besteht aus einem Molekül N-Acetylglucosamin, einem Molekül N-Acetyl-Mannosamin und drei Molekülen Glycerolphosphat. Über Modifikationen dieser Ankerstruktur wurde berichtet, jedoch scheint der GlcNAc-1-Phosphat Terminus überall gleich zu sein. An die Ankerstruktur schließt sich eine Kette aus alternierenden Alditolphosphaten an, die den Hauptteil der WTA ausmachen. Am weitesten verbreitet sind Ketten aus Glycerolphosphat- und Ribitolphosphateinheiten. S. aureus produziert eine WTA, die aus ~40 Ribitolphosphateinheiten zusammengesetzt ist, wohingegen andere Staphylokokken auch Glycerolphosphatketten besitzen. Die Ribitolphosphateinheiten sind teilweise modifiziert durch N-Acetylglucosamin und D-Alanin.[9][10][11]
Wirkung beim Menschen
Teichonsäuren und Lipoteichonsäuren sind starke exogene Pyrogene, d.h. sie zählen zu den Substanzen, die beim Menschen nach einer bakteriellen Infektion durch gram-positive Bakterien eine fieberhafte Reaktion hervorrufen. Sie werdem von dem Toll-like Rezeptor TLR-2 erkannt, der auf Monocyten und Dendritischen Zellen, B- und T-Lymphocyten sowie Makrophagen exprimiert wird. Weiterhin sorgen sie für die Ausschüttung von Zytokinen und sind damit einer der Hauptfaktoren für die Entzündungsreaktion nach einer solchen Infektion.[1]
Wegen ihrer Antigen-Eigenschaft sind sie auch interessant zur Herstellung von synthetischen Impfstoffen.[12]
Quellen
- Thomas Kohler: Untersuchungen zur Biosynthese und Wechselwirkung der Zellwandteichonsäuren von Staphylococcus aureus. 2005.
Einzelnachweise
- ↑ a b H. Hof, R. Dörries: Medizinische Mikrobiologie. 3. Auflage, S. 273, 2004, Georg Thieme Verlag, ISBN 3131253134
- ↑ a b c d Fischer, W. (1990)
- ↑ J. M. Ghuysen and R. Hakenbeck (Eds), 1994
- ↑ Fischer 1994
- ↑ Baddiley, J. 1988
- ↑ Fischer, W. 1988
- ↑ Deutsch, R., Engel, R. and Tropp, B.E. 1980
- ↑ Brisette, J.L., Cabacungan, E.A. and Pieringer, R.A. 1986
- ↑ Endl, J., Seidl, H.P., Fiedler, F. & Schleifer, K.H.,1983
- ↑ Araki, Y. and ITO, e. 1989
- ↑ Mikusova, K., Mikus, M., Besrat, G. S., Hancock, I. and Brennan, P. J., 1996
- ↑ G. G. Habermehl, H. C. Krebs, P. E. Hammann: Naturstoffchemie. S. 390, 2002, Springer-Verlag, ISBN 3540439528
Wikimedia Foundation.