- Acetabularia
-
Schirmalgen 
Acetabularia spec.
Systematik ohne Rang: Chloroplastida ohne Rang: Chlorophyta Klasse: Ulvophyceae Ordnung: Dasycladales Familie: Polyphysaceae Gattung: Schirmalgen Wissenschaftlicher Name Acetabularia J.V.Lamour. Die Schirmalgen (lat. Acetabularia) sind eine Gattung der einzelligen Algen, welche zwischen 0,5 und 10 Zentimeter groß werden können. Die Gattung besteht aus ca. 15 verschiedenen Arten.[1] Die einzelnen Arten werden durch die Form des Schirmes unterschieden. Wegen der Einlagerung von Kalk in ihrer Zellwand ist die Gattung der Schirmalgen maßgeblich am Entstehungsprozess eines Riffes beteiligt.
Inhaltsverzeichnis
Lebensraum und Verbreitung
Algen aus der Familie der Schirmalgen kommen in tropischen und subtropischen Meeren vor. Im Mittelmeer und am Ostatlantik trifft man häufig die Arten Parvocaulis parvulus (Synonyme: Acetabularia parvula und Acetabularia wettsteinii) und Acetabularia acetabulum an. (In den meisten älteren Büchern wird letztere Art noch als A. mediterranea geführt. Neuere Forschungen zeigten aber, dass A. mediterranea und A. acetabulum zur selben Art gehören). [2] Die Art Acetabularia crenulata ist ein weiterer Vertreter dieser Gattung, sie kann bis zu fünf Zentimeter lang werden und kommt in der Karibik vor. Schirmalgen sind auch um die Westindischen Inseln zu finden.
Aufbau
Die Acetabularia-Zelle lässt sich äußerlich in drei Abschnitte gliedern:
- in das wurzelähnliche Rhizoid,
- den meist verkalkten, radial-symmetrisch gebauten Thallus (Vegetationskörper), und
- den Schirm.
Alle diese Abschnitte weisen spezifische Funktionen auf.
Bestandteil Funktion Rhizoid - Verankerung im Boden
- beinhaltet Primärkern
Thallus - besitzt Auswüchse mit Seitenästen
- enthält viele kleine Chloroplasten
Schirm - Fortpflanzung
Die Zellwände weisen häufig starke Kalkeinlagerungen auf. Eine weitere Besonderheit ist, dass in den vielen kleinen Chloroplasten keine Pyrenoide vorhanden sind. Dadurch wird keine Stärke, sondern es werden Fructosane gebildet, welche mit Hilfe von Insulin abgebaut werden.
Lebenszyklus
 Lebenszyklus einer A. acetabulum
Lebenszyklus einer A. acetabulumDer Lebenszyklus dieser Algengattung beträgt 3 Jahre. In der neben stehenden Abbildung wird er am Beispiel der A. acetabulum gezeigt. Die Zystenwand platzt im vierten Frühjahr nach dem letzten Durchlaufen des Zyklus auf und setzt viele Isogameten frei (Bildteil 1). Eine Verschmelzung findet ausschließlich zwischen Isogameten derselben Alge und unterschiedlicher Zysten statt (Bildteil 2). Es entwickelt sich im ersten Jahr zuerst ein unseparierter, aufrechter Schlauch (Bildteil 3), später ein Rhizoid mit Basalblase. In ihr ist der Zygotenkern enthalten, welcher bis auf die Größe von 100 µm heran wächst. Er wird dann als Primärkern bezeichnet (Bildteil 4). Im Herbst stirbt der Schlauch ab und die A. acetabulum überwintert im Rhizoid und in der Basalblase. Im Frühling des zweiten Jahres bildet die Schirmalge einen weiteren Schlauch aus. Er besitzt an seiner Spitze mehrere Wirtel aus verzweigten dichotomen Ästchen (Bildteil 5). Wenn der Herbst wieder Einzug hält, stirbt auch dieser Schlauch ab und die A. acetabulum begibt sich zur Winterruhe. Das dritte Jahr ist von der Vorbereitung der Vermehrung geprägt. Es wird ein dritter Schlauch ausgebildet, an dessen Ende bildet sich ein Wirtel mit unverzweigten, breiten, lappenförmigen Ausstülpungen (Sektorkammern). Man nennt diese Gebilde dann Schirm oder Schirmstrahlen. Wenn der Schirm ausgebildet ist, zerfällt der Primärkern in bis zu 15000 haploide Sekundärkerne, welche mit dem Plasmastrom zu den Schirmstrahlen transportiert werden. Sie lagern sich gleichmäßig an der Zellwand ab und bilden Zysten. Dort nimmt ihr Volumen stark zu und sie werden zu primären Zystenkernen. Sie bilden scharf abgegrenzte, dickwandige, mit Kalkkrusten und Deckeln versehene Hüllen. Dort teilen sich die Primärkerne sehr rege mitotisch (Bildteil 6). Der Schlauch verwelkt und es bleiben nur noch die Zysten übrig, welche sich in Zystenruhe befinden. Zum Ende dieser Winterruheperiode teilen sich die primären Zystenkerne meiotisch und bilden wieder die Isogameten (Bildteil 7).
Damit ergibt sich ein diplohaplontischer (also teils diplontischer, teils haplontischer) Entwicklungsgang, obwohl nur eine Form, nämlich der Gametophyt existiert. Durch die meiotische Teilung des Kernes (bzw. der Kerne) reduziert sich der Ploidiegrad des ansonsten diploiden Gametophyten und er wird im letzten Vegetationszyklus somit haploid. Die aus der Gametenverschmelzung entstehenden neuen Schirmalgen sind wieder diploid. Ein und dieselbe biologische Form ist also (je nach Vegetationszyklus) diploid oder haploid.
Bedeutung
Die Gattung der Acetabularia hat in verschiedenen Bereichen eine große Bedeutung. Zum einen in der Geologie, da durch die schon erwähnte starke Kalkeinlagerung sehr viele Fossilien entstanden sind. Zehn der heute noch bestehenden Arten lassen sich bereits in der Kreidezeit nachweisen. In der Natur nimmt sie eine Stellung als Riffbildner ein. Auch in der Wissenschaft hat Acetabularia eine herausragende Stellung. 1932 konnte an ihr der Nachweis der Bedeutung des Zellkerns erbracht werden. In der damaligen Zeit war eine molekularbiologische Untersuchung unmöglich. Der Nachweis wurde auf der mikroskopischen Ebene geführt. Man verwendete dazu verschiedene Pfropf- und Teilungsexperimente. Einige von ihnen werden im Folgenden vorgestellt. Allgemein lässt sich eine enorme Regenerationsfähigkeit dieser Algenart verzeichnen. Sie ist in der Lage nach dem Verlust des Zellkerns noch 3-7 Monate lebensfähig zu bleiben, Zellulose und Eiweiß zu produzieren und Formbildung zu durchlaufen.
Teilungsexperimente
Amputation des Rhizoids
Wenn Hut und Stiel entfernt werden, wird vom Rhizoid aus beides neu gebildet.
 Versuchsverlauf
Versuchsverlauf
# Entfernung des Rhizoids
# nach einiger Zeit Neubildung eines kernlosen RhizoidsAmputation des Hutes
Nach der Amputation wird vom Zellkern aus der Befehl zur Ausschüttung morphogener Stoffe gegeben, welche nach kurzer Zeit eine Neubildung des Hutes initiieren.
 Versuchsverlauf
Versuchsverlauf
# Entfernung des Hutes
# nach einiger Zeit Neubildung eines HutesAmputation von Rhizoid und Hut
Der Stiel regeneriert sich je nach Ort der Teilung anders. Je größer das Stück desto einfacher wird das fehlende Stück neu gebildet. Man unterscheidet hierbei 3 Szenarien:
- Der Stiel wird dicht über dem Rhizoid getrennt. Dies führt zur Ausbildung zweier kernloser Rhizoide.
- Es wird ein Stück Stiel ungefähr aus der Mitte zwischen Rhizoid und Hut entnommen. Das hat zur Folge, dass ein kernloses Rhizoid und ein Hut ausgebildet wird.
- Man entfernt ein Stück des Stiels unterhalb des Hutes und entfernt auch das Rhizoid. Als Ergebnis werden hier ein normaler Hut und ein "Kümmerhut" anstelle des Rhizoids ausgebildet.
Man kann hieraus eine polare Verteilung von mindestens 2 morphogenen Regenerationsstoffen, die sich im gegenläufigen Stoffgefälle befinden, ersehen. Die Entwicklung der Regenerate hängt von der Menge dieser Stoffe ab. Im kernlosen Abschnitt werden diese Substanzen nicht mehr regeneriert.
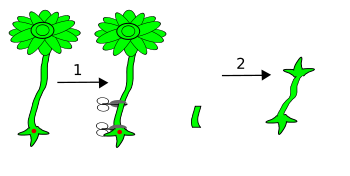 Versuchsverlauf Szenario 1
Versuchsverlauf Szenario 1 Versuchsverlauf Szenario 2
Versuchsverlauf Szenario 2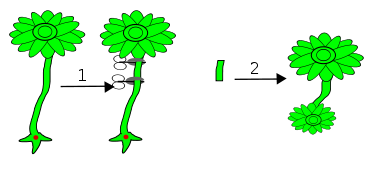 Versuchsverlauf Szenario 3
Versuchsverlauf Szenario 3
Doppelter Regenerationsversuch
Bei diesem Versuch wurde nachgewiesen, dass vom kernhaltigen Stück die für die Regeneration verantwortlichen Stoffe produziert werden. Er wird in zwei Schritten durchgeführt. Zuerst wird ein Stück der Stielspitze entfernt und später auch das Rhizoid. Die Stielspitze wurde nachgebildet, weil die Bildungsstoffe bereits produziert und dann in die Spitze transportiert wurden. Daraus folgt, dass der Zellkern für die Ausbildung verantwortlich ist.
 Versuchsverlauf
VersuchsverlaufPfropfversuche
Rhizoid von Parvocaulis parvulus und Stiel von A. acetabulum
Ein kernhaltiges Rhizoid der Art Parvocaulis parvulus wird von seinem Stiel und seinem Hut getrennt und der Stiel von A. acetabulum wird auf dieses Rhizoid aufgepfropft. Es entwickeln sich zuerst A. acetabulum-Wirtel, aber die Ausprägung des Hutes ist eindeutig Parvocaulis parvulus.
Rhizoid von A. acetabulum und Stiel von A. crenulata
In diesem Experiment wird auf das Rhizoid der A. acetabulum ein Stiel der A. crenulata aufgepfropft. Das Ergebnis unterscheidet sich aber von dem des vorangegangenen Versuches. Es entsteht ein intermediärer Hut. Wird dieser abgetrennt, so entwickelt sich nur noch ein Hut der A. acetabulum.
Rhizoid der A. crenulata und Rhizoid der A. acetabulum
Verpfropft man zwei Rhizoide unterschiedlicher Acetabularia-Arten, so entstehen intermediäre Hüte.
Zwei Rhizoide der A. crenulata und ein Rhizoid A. acetabulum
Bei dieser Kombination entstehen wiederum intermediäre Hüte, welche aber mehr der A. crenulata ähneln.
Austausch von Pflanzenteilen
Wenn man ein altes Rhizoid mit einem jungen Stiel verbindet, so verlängert sich der Entwicklungsprozess. Wenn das Verhältnis von alt und jung entgegengesetzt ist, so wird dieser Prozess verkürzt. Der Primärkernteilungsprozess beginnt erst, wenn der Hut vollständig ausgebildet ist. Er verschiebt sich bei der Amputation des Hutes nach hinten. Wird nach Beginn dieses Zerfallsprozesses der Hut abgetrennt, ist die Schirmalge nicht mehr in der Lage einen neuen Hut auszubilden. Wird ein neuer Hut aufgepfropft, so bilden sich Zysten und der Vermehrungsprozess schreitet fort.
Systematik
- Acetabularia acetabulum (L.) P.C.Silva (syn. Acetabularia mediterranea)
- Acetabularia antillana (Solms) Egerod
- Acetabularia calyculus J.V.Lamour.
- Acetabularia crenulata J.V.Lamour.
- Acetabularia dentata Solms
- Acetabularia farlowii Solms
- Acetabularia kilneri J.Agardh
- Acetabularia major G.Martens
- Acetabularia myriospora A.B.Joly & Cord.-Mar.
- Acetabularia peniculus (R.Br. ex Turner) Solms
- Acetabularia ryukyuensis Okamura & Yamada
- Acetabularia schenckii K.Möbius
- Acetabularia toxasii Troño, A.E.Santiago & Ganz.-Fort.
Quellen
Einzelnachweise
- ↑ http://www.algaebase.org/browse/taxonomy/?id=8297&
- ↑ http://www.algaebase.org/speciesdetail.lasso?species_id=16755
Bücher
- Günter Vogel und Hartmut Angermann: Taschenbuch der Biologie. 2. Auflage. VEB Gustav Fischer Verlag, Jena 1979, Lizenznummer:261 700/174/79 Bd.1
- Dr.rer.nat Erik Heinz Benedix, Dr.rer.nat.habil S. Jost Casper u. a.: Urania Pflanzenreich. 1. Auflage. Urania-Verlag, Leipzig/Jena/Berlin 1974, VLN. 212-475/26/74 Bd.: Niedere Pflanzen
- Wolfgang Miram und Karl-Heinz Scharf (Hrsg.): Biologie heute SII. Schroedel Schulbuchverlag, Hannover 1988, ISBN 3-507-10540-3
Weblinks









Wikimedia Foundation.