- Dinophyta
-
Dinoflagellaten Systematik Klassifikation: Lebewesen Domäne: Eukaryoten (Eucaryota) ohne Rang: Chromalveolata Unterreich: Alveolaten (Alveolata) Stamm: Dinozoa Unterstamm: Dinoflagellaten Wissenschaftlicher Name Dinoflagellata (Bütschli 1885) Cavalier-Smith 1991 Die Dinoflagellaten (Dinoflagellata) sind ein Unterstamm, der vorwiegend Einzeller umfasst. Zu ihren kennzeichnenden Merkmalen gehören zwei Flagellen während des mobilen Lebenszyklus, Chromosomen, die während der Interphase kondensiert sind, sowie das Fehlen von Histonen. Weltweit werden heute rund 1.000 rezente Arten in 120 Gattungen[1] unterschieden, die großteils im Meer leben und dabei einen Hauptteil des Phytoplanktons bilden. Der Unterstamm umfasst autotrophe als auch heterotrophe Arten.
Inhaltsverzeichnis
Merkmale
Grundsätzliche Merkmale
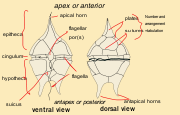 Basaler Bauplan eines Dinoflagellaten
Basaler Bauplan eines DinoflagellatenInnerhalb der Dinoflagellaten herrscht eine extrem große Formenvielfalt. Diese können jedoch als Modifikationen einer Basisform angesehen werden. Die Größe reicht von 2 µm (Gymnodinium simplex) bis zu 2 mm (Noctiluca sp.), wobei die meisten Arten zwischen 20 und 200 µm groß werden. Die Form der freischwimmenden Zelle (Mastigote) ist eiförmig bis rundlich, wobei das Anterior meist mehr zugespitzt ist als das Posterior. Die Dinoflagellaten besitzen zwei lange Geißeln wobei eine nach hinten gerichtet (Transversale Geißel) ist und die andere in einer Ebene senkrecht dazu schlägt (Longitudinale Geißel). Diese Anordnung der Geißeln wird als dinokont bezeichnet.
Bei vielen Arten sind die direkt unterhalb der Zellmembran liegenden Alveolen mit Zellulose gefüllt, und so zu mehr oder weniger massiven Platten verstärkt[2]. Wenn solche intrazellulären Platten vorhanden sind, wird diese Hülle als Theka und die entsprechenden Arten thekat bezeichnet. Wenn die Alveolen nicht oder nur sehr wenig verstärkt sind, werden die Arten athekat oder nackt genannt. Die Theka bildet ein Mosaik aus einzelnen Platten, und kann zur Artbestimmung benutzt werden.
Eine Furche, der sogenannte Gürtel (Cingulum) läuft rund um die Zelle und teilt diese somit in ein Anterior (Episoma) und Posterior (Hyposoma). Ist eine Theka vorhanden, werden die Teile als Epitheka bzw. Hypotheka bezeichnet. Zum Posterior verläuft eine weitere Furche, der sogenannte Sulcus. Die transversale Geißel schlägt im Cingulum, die longitudinale Geißel im Sulcus.
Genetik
Innerhalb der Eukaryonten besitzen Dinoflagellaten einzigartige Eigenschaften. Die Chromosomen sind auch während der Interphase kondensiert und im Elektronenmikroskop sichtbar. Die Chromosome bilden eine Girlandenstruktur, wobei die einzelnen Fibrillen nur 2,5 nm im Durchmesser haben, was auf das Fehlen von Histonen zurückzuführen ist. Die übrigen Eukaryonten besitzen Fibrillen mit zehnfachem Durchmesser mit einem zentralen Nucleohistonstrang. Weiters ist nur innerhalb der Dinoflagellaten die modifizierend Base Hydroxymethyluracil (HOMeU) in der DNA nachgewiesen. Mit einem Gesamtanteil von 4-19% ersetzt sie 12-70% der Thymin-Basen[3] Die Chromosomenzahl schwankt zwischen 5 bei Syndinium turbo und 274 bei Ceratium hirundinella.[4]
Biolumineszenz
 Biolumineszente Welle zusammen mit "Roter Tide", Carslbad, Californien 2005
Biolumineszente Welle zusammen mit "Roter Tide", Carslbad, Californien 2005Einige Arten sind zur Biolumineszenz fähig, wobei dieses Leuchten eine Reaktion auf mechanische Stimulation ist. In der Natur sind dies Deformationen der Zellmembran, die durch Scherkräfte hervorgerufen werden. Stark aufgewühltes Wasser, wie brechende Wellen oder schnell schwimmende Fische können solche Stimulationen auslösen. Im Labor kann auch mittels Chemikalien eine Reaktion induziert werden. Zu den Dinoflagellaten gehören die einzigen biolumineszenten autotrophen Lebewesen wie etwa Vertreter der Gattungen Gonyaulax, Protogonyaulax, Pyrodinium und Pyrocystis. Auch bei heterotrophen Arten wie Noctiluca miliaris oder einige Vertreter der Gattungen Ceratium kann Biolumineszenz beobachtet werden.
Das emittierte Licht ist blau-grün und hat ein Maximum bei 474-476 nm. Da diese Wellenlänge nahe dem maximalen Transmissionsgrad des Meerwassers liegt, wird angenommen, dass die Sichtbarkeit des Lichtes den selektiven Vorteil verursacht. In Experimenten mit leuchtenden und nicht-leuchtenden Spezies konnte gezeigt werden, dass im Falle von Biolumineszenz die Prädation vermindert wurde. Vermutlich werden Feinde durch den Lichtblitz abgeschreckt. Wie bei fast allen Arten der Biolumineszenz ist dies auf eine Reaktion von Luciferasen und Luciferinen zurückzuführen.
Toxine
Einige Arten produzieren äußerst starke Gifte. Das Saxitoxin beispielsweise wird von Vertretern der Gattung Alexandrium (Gonyaulax) produziert. Wenn die giftigen Dinoflagellaten von Muscheln gefressen werden, reichert sich das Gift in den Muscheln an, und kann dann auch für Menschen gefährlich werden. Bei einer Massenvermehrung von giftigen Arten wird soviel Gift produziert, dass auch Fische und andere Meereslebewesen getötet werden[5].
Die Krankheit Ciguatera[6], eine Art Fischvergiftung, wird durch Stoffwechselprodukte der Art Gambierdiscus toxicus hervorgerufen. Über die Nahrungskette gelangen die Dinoflagellaten-toxine Ciguatoxin und Maitotoxin in Fische, die dadurch ebenfalls stark giftig werden. Die Vergiftung kann unter Umständen beim Menschen tödlich verlaufen.
Das Toxin von Pfiesteria piscicida dagegen wird nicht über die Nahrungskette angereichert, sondern ist direkt giftig für Fische und Menschen[7].
Verbreitung und Lebensräume
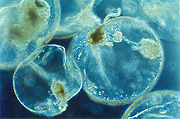 Noctiluca scintillans ist ein marines Lebewesen
Noctiluca scintillans ist ein marines LebewesenDinoflagellaten sind kosmopolitisch im Salz- auch als im Süßwasser verbreitet und können dort aufgrund ihres Formenreichtums viele Habitate besiedeln. Rund 90% aller Arten werden dem marinen Plankton zugerechnet, mit der größten Artenvielfalt in tropischen Gewässern. Sie sind aber auch benthische Lebewesen und dringen auch in die Sedimente ein. Weiters sind sie ebenfalls in der Polarregion oder in Meereis anzutreffen.
Im Süßwasser sind weniger Arten verbreitet. Bis 1970 wurden 220 Süßwasser-Arten beschrieben, die weltweit Seen, Tümpel und Moore besiedeln. Das Verbreitungsgebiet reicht etwa vom Äquator bis 78° nördlicher Breite (Spitzbergen (Insel)). Die Höhenunterschiede reiche von -209 Meter in Israel bis auf 4150 Meter in Hochgebirgsseen von Mexiko.
Da einige Arten Symbiosen eingehen oder als Parasiten leben, werden auch Lebewesen als Habitate genutzt. Autotrophe Arten sind auf lichtdurchflutete Wasserschichten angewiesen, heterotrophe Arten können auch in vollkommen dunkle Tiefen vordringen.
Ernährung
Etwa die Hälfte der Dinoflagellaten ist autotroph und können mit Hilfe Assimilation der Chloroplasten anorganischen Kohlenstoff nutzen. Jedoch sind fast sämtliche photosynthetisch aktive Arten auxotroph und benötigen Vitamine (Cobalamine, Biotin, Thiamin) für katalytische Zwecke. Diese werden über Phagocytose aufgenommen. Autotrophe Arten gehen auch eine Symbiose mit Nesseltiere (Cnidaria) insbesondere Korallen, Weichtiere (Mollusca) aber auch Foraminiferen (Foraminifera) und Wimpertierchen (Ciliata), ein.
Heterotrophe Dinoflagellaten ernähren sich von von einem vielfältigen Spektrum von Planktonorganismen, das von Nanoplankton bis zu großen Kieselalgen reicht[8][9]. Darunter fallen auch Dinoflagellaten der eigenen wie auch anderer Arten, Detritus und selbst Eier und Larven von Ruderfußkrebsen. Im einfachsten Fall wird die Nachrung durch Phagocytose aufgenommen (beispielsweise Noctiluca). Durch spezielle Zellstrukturen wie Pedunkel oder Pallium können sich heterotrophe Dinoflagellaten aber auch von Organismen ernähren, die um ein Vielfaches größer als sie selbst sind (beispielsweise Pfiesteria[10] oder Protoperidinium)[8][9].
Autotrophie
Die autotrophen Arten enthalten Plastiden mit Chlorophyll a bzw. einige Arten auch Chlorophyll c. Als Haupt-Carotinoid enthalten sie meist Peridinin anstatt von Fucoxanthin. Ihre Färbung reicht von gelbbraun bis rötlich, da das Chlorophyll von braunen und gelben Carotinoiden überdeckt wird. Stärke ist das Hauptassimilationsprodukt, das in Körnchen außerhalb der Chloroplasten gespeichert werden. Es wurden aber auch fettartige Stoffe nachgewiesen. Die Plastidenwand besteht meist aus drei Membranen, die nicht mit dem Endoplasmatischen Retikulum verbunden sind.
Grundsätzlich können Dinoflagellaten sehr verschiedene Plastiden beherbergen, die vom Grundtyp abweichen. Dies ist auf Phagotrophie zurückzuführen die auch bei autotrophen Arten aufrechterhalten wird. Dies führte in der Stammesgeschichte zu einer weiteren, tertiären Endocytobiose. Die aufgenommenen Organismen können hierbei aus unterschiedlichen Gruppen, wie Haptophyta, Cryptophyceae, Heterokontophyta oder eines Chlorophyten zurückgehen. Der ursprünglich von den Rotalgen stammende Chloroplast ist hierbei völlig oder weitgehend zurückgebildet und erscheint im letzteren Fall als inaktiver Augenfleck (Stigma). Gelegentlich ist in den Chloroplasten auch ein Nucleomorph enthalten.
Grundtypen der Heterotrophie
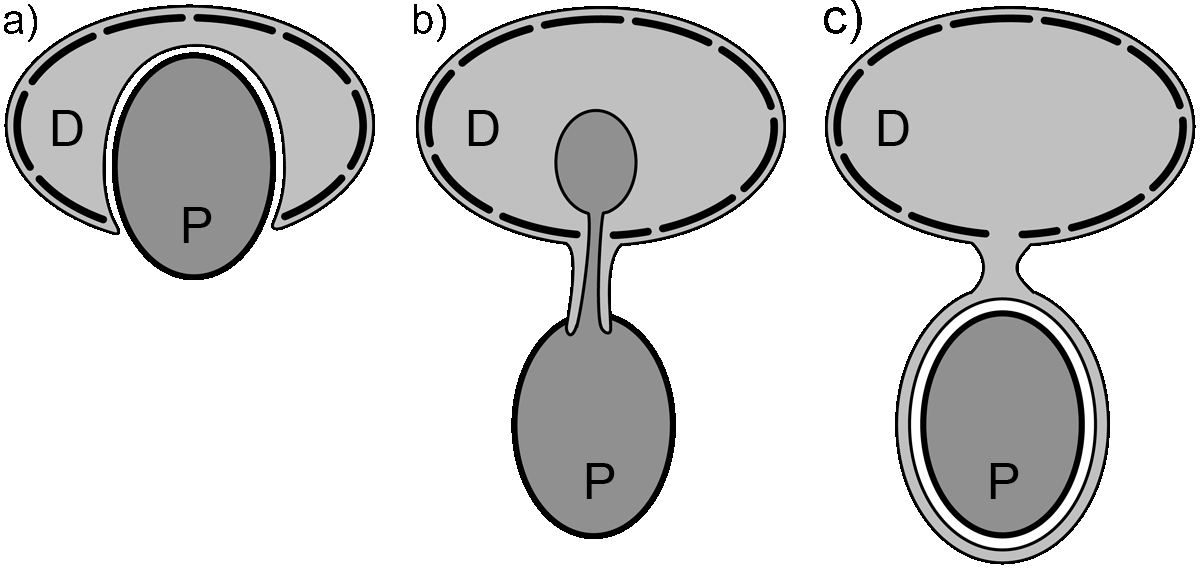 Grundtypen der Heterotrophie bei Dinoflagellaten
Grundtypen der Heterotrophie bei DinoflagellatenDie Fraßmechanismen heterotropher Dinoflagellaten lassen sich mit drei Grundtypen beschreiben.
- Phagozytose: Die Beute wird durch den Sulcus direkt und vollständig aufgenommen und in eine Fraßvakuole eingeschlossen (beispielsweise Noctiluca).
- Myzozytose: Vom Sulcus aus wird ein charakteristischer Plasmastrang ausgestülpt. Dieser Pedunkel durchstößt die Zellhülle der Beute, und saugt den Zellinhalt in eine Fraßvakuole. Die Zellbestandteile der Beute werden dabei nicht sofort verdaut. Chloroplasten beispielsweise können erhalten bleiben und so als Kleptoplastiden im Dinoflagellaten weiter funktionieren (beispielsweise Dinophysis[11]). Die Zellhülle der Beute wird nicht in die Vakuole eingeschlossen.
- Pallium: Vom Sulcus aus wird ein schlauch- oder segelförmiges Pseudopodium ausgestülpt, dass die Beute mit einer dünnen Lage Zellplasma umgibt und so eine Fraßvakuole bildet. Da diese Vakuole außerhalb der Theka gebildet wird, ist die Größe des Pallium und damit die Größe der Beute nicht von der Größe des Dinoflagellaten begrenzt, so dass auch eine um ein Vielfaches größere Beute verdaut werden kann (beispielsweise Protoperidinium[12]). Im Pallium wird die Beute verdaut, und in verflüssigter Form aufgenommen.
Die Grundtypen Myzozytose und Pallium kommen hauptsächlich bei thekaten (gepanzerten) Arten vor, und werden gelegentlich als extrazelluläre Verdauung bezeichnet. Dies stimmt genau betrachtet nicht, denn in jedem Fall wird die Beute in einer Fraßvakuole innerhalb des Zellplasmas verdaut, jedoch können sich diese Fraßvakuolen außerhalb der Theka befinden. Dies kann als Überwindung der Beschränkungen durch die Theka interpretiert werden und eröffnete den heterotrophen Arten ein erweitertes Beutespektrum.
Beutefang
Der untypische Dinoflagellat Noctiluca miliaris besitzt einen kurzen Tentakel, der wie eine Leimrute einsetzt wird. Nahrungspartikel wie Kieselalgen und Detritus bleiben daran hängen, und werden dann vom Tentakel zum Cytostom befördert.
Der thekate Dinoflagellat Stoeckeria algicida dagegen nutzt ein schlagartig ausgestoßenes Proteinfilament (englisch: tow filament), um die Beute über eine vergleichsweise große Entfernung einzufangen[13]. Ein vergleichbares Filament wird von Protoperidinium benutzt, um sich an Kieselalgen-ketten zu verankern[8].
Fortpflanzung
Die Fortpflanzung erfolgt überwiegend vegetativ. Bei bepanzerten Arten werden die Platten in der Regel schräg zum Gürtel gesprengt, wobei die fehlende Hälfte später nachwächst. Es besteht aber auch die Möglichkeit, dass der Panzer abgeworfen und von den Tochterzellen völlig neu gebildet wird. Unter ungünstigen Lebensbedingungen entstehen dickwandige, überdauerungsfähige Zysten.
Geschlechtliche Fortpflanzung wurde nur bei wenigen Arten nachgewiesen. Hierbei wurden Anisogamie mit zygotischem Kernphasenwechsel als auch Isogamten, die in Gametangien entstehen, freigesetzt werden und miteinander verschmelzen, beschrieben.
Ökologische Bedeutung
Zusammen mit den Kieselalgen sind die Dinoflagellaten die Hauptprimärproduzenten organischer Stoffe im Meer, bilden dort also zusammen mit den Kieselalgen den Hauptteil der Basis der Nahrungspyramide. In Hochgebirgsseen können sie bis zu 50% der Biomasse ausmachen.
Die heterotrophen Dinoflagellaten können mit ihren spezialisierten Fraßmechanismen ein weites Spektrum von Beuteorganismen fressen, das von Nanoplankton kleiner als 10 µm bis zu großen kettenbildenden Kieselalgen reicht. Dadurch stellen die heterotrophen Dinoflagellaten einen wichtigen Teil der mikrobiellen Schleife[14] dar (englisch: microbial loop[15]).
Unter für sie günstigen Bedingungen vermehren sich in tropischen und subtropischen Gewässern bestimmte Arten in extremem Ausmaß, so dass sich die oberen Schichten des Meeres rot bis braun färben. Man nennt diese Algenblüte auch Rote Flut oder Rote Tide (englisch "red tide").
Biostratigraphie
Durch ihre sehr widerstandsfähige, organische Zellwand werden Dinoflagellatenzysten nicht durch Kalklösung zerstört, sondern bleiben auch nach langen Zeiträumen erhalten. Außerdem haben viele Zysten eine charakteristische Form. Das spielt für die Altersdatierung (Biostratigraphie) von Sedimenten eine entscheidende Rolle.
Andere Fossilgruppen wie Foraminiferen besitzen eine zu geringe Artenvielfalt und Dinoflagellatenzysten treten nahezu in allen Gewässern auf, wo sie heute als Klimaindikatoren verwendet werden. Erst 1988 begann man in Deutschland mit der Aufstellung von „Dinoflagellaten-Zonen“, die nun regelmäßig verbessert werden.
Systematik
Durch die teilweise sehr komplexen Lebenszyklen der Dinoflagellaten ist die systematische Gliederung Gegenstand wissenschaftlicher Diskussion. Das Taxon gilt als polyphyletisch. Die hier angeführte Gliederung (Gattungen exemplarisch) folgt Adl et al. 2005[16]
- Dinophyceae
- Gymnodiniphycidae
- Amphidinium
- Gymnodinium
- Ptychodiscus
- Symbiodinium
- Woloszynskia
- Peridiniphycidae
- Alexandrium
- Amphidinopsis
- Amphidoma
- Blepharocysta
- Ceratium
- Crypthecodinium
- Gonyaulax
- Heterocapsa
- Peridiniella
- Peridinium
- Pfiesteria
- Pyrocystis
- Dinophysiphycidae
- Dinophysis
- Triposolenia
- Prorocentrales
- Prorocentrum
- Phytodiniales
- Dinothrix
- Hemidinium
- Gymnodiniphycidae
- Blastodiniales
-
- Crepidoodinium
- Dissodinium
-
- Noctilucales
-
- Kofoidinium
- Noctiluca
-
- Syndiniales
-
- Amoebophrya,
- Duboscquella
- Merodinium
- Syndinium
-
Belege
Soweit nicht unter Einzelnachweisen angegeben, basiert der Artikel auf folgenden Unterlagen:
- C. Körner, G. Neuhaus, U. Sonnewald, J. W. Kadereit, A. Bresinsky, C. Körner: Lehrbuch der Botanik für Hochschulen. Begründet v. E. Strasburger. Spektrum Akademischer Verlag, Heidelberg 362008, ISBN 978-3-8274-1455-7
- Taylor: The Biology of dinoflagellates. Blackwell, 1987, ISBN 0-632-00915-2
- Ettl (Hrsg.): Süßwasserflora von Mitteleuropa: Dinophyceae, Gustav Fischer Verlag, 1990, ISBN 3-334-00247-0
Einzelnachweise
- ↑ C. Körner, G. Neuhaus, U. Sonnewald, J. W. Kadereit, A. Bresinsky, C. Körner: Lehrbuch der Botanik für Hochschulen. Begründet v. E. Strasburger. Spektrum Akademischer Verlag, Heidelberg 362008, ISBN 978-3-8274-1455-7, Seite 703
- ↑ Loeblich, Alfred R. & Loeblich, Laurel A. (1985).Dinoflagellates: Structure of the amphiesma and re-analysis of thecal plate patterns. Hydrobiologia, vol. 123, no. 2: 177-179. (Abstract und Volltext)
- ↑ Taylor: The Biology of dinoflagellates. Blackwell, 1987, ISBN 0-632-00915-2, Seite 160
- ↑ Taylor: The Biology of dinoflagellates. Blackwell, 1987, ISBN 0-632-00915-2, Seite 618
- ↑ Landsberg J.H. (2002). The effects of harmful algal blooms on aquatic organisms. Reviews in Fisheries Science, 10(2): 113–390.
- ↑ Swift A., Swift T. (1993). Ciguatera. J. Toxicol. Clin. Toxicol. 31 (1): 1–29. (Abstract)
- ↑ Moeller P.D., Beauchesne K.R., Huncik K.M., Davis W.C., Christopher S.J., Riggs-Gelasco P., Gelasco A.K. (2007). Metal complexes and free radical toxins produced by Pfiesteria piscicida. Environ. Sci. Technol. 41 (4): 1166–72. (Abstract und Volltext)
- ↑ a b c Schnepf, Eberhard and Elbrächter, Malte (1992). Nutritional strategies in dinoflagellates. Europ. J. Phycology, vol. 28: 3 – 24.
- ↑ a b Hansen, Per J. and Calado, António J. (1999). 'Phagotrophic mechanisms and prey selection in free- living dinoflagellates'. Journal of Eukaryotic Microbiology, vol. 46, no. 4: 382 – 389. (Abstract und Volltext)
- ↑ Gordon, A. and Dyer, B. (2005). Relative contribution of exotoxin and micropredation to icthyotoxicity of two strains of Pfiesteria shumwayae (Dinophyceae) Harmful algae, vol. 4, no. 2: 423 – 431.
- ↑ Kiyotaka Takishitaa, Kazuhiko Koikeb, Tadashi Maruyamaa and Takehiko Ogatab (2002). Molecular Evidence for Plastid Robbery (Kleptoplastidy) in Dinophysis, a Dinoflagellate causing Diarrhetic Shellfish Poisoning. Protist, Vol. 153, 293–302. (Abstract)
- ↑ Gaines, G. & Taylor, F.J.R. (1984).Extracellular digestion in marine dinoflagellates. Journal of Plankton Research, Vol. 6, No. 6: 1057-1061
- ↑ Hae Jin Jeong, Jae Seong Kim, Jong Hyeok Kim, Seong Taek Kim, Kyeong Ah Seong, Tae Hoon Kim, Jae Yoon Song, Soo Kyeum Kim (2005). Feeding and grazing impact of the newly described heterotrophic dinoflagellate Stoeckeria algicida on the harmful alga Heterosigma akashiwo. Mar Ecol Prog Ser, Vol. 295: 69–78.
- ↑ Naustvoll, Lars J. (2001). The role of heterotrophic dinoflagellates in marine pelagic food webs. Dissertation, University of Oslo.
- ↑ Azam, S. D.; Fenchel, T.; Field, J. G.; Gray, J. S.; Meyer- Reil, L. A.; Thingstad, F. (1983). The ecological role of water- column microbes in the sea. Marine Ecology Progress Series, vol. 10: 257 – 263. (Volltext)
- ↑ Sina M. Adl, Alastair G. B. Simpson, Mark A. Farmer, Robert A. Andersen, O. Roger Anderson, John A. Barta, Samual S. Bowser, Guy Bragerolle, Robert A. Fensome, Suzanne Fredericq, Timothy Y. James, Sergei Karpov, Paul Kugrens, John Krug, Christopher E. Lane, Louise A. Lewis, Jean Lodge, Denis H. Lynn, David G. Mann, Richard M. McCourt, Leonel Mendoza, Øjvind Moestrup, Sharon E. Mozley-Standridge, Thoams A. Nerad, Carol A. Shearer, Alexey V. Smirnov, Frederick W. Spiegel, Max F. J. R. Taylor: The New Higher Level Classification of Eukaryotes with Emphasis on the Taxonomy of Protists. The Journal of Eukaryotic Microbiology 52 (5), 2005; Seiten 433 (Abstract und Volltext)
Weblinks


Wikimedia Foundation.