- Echinoderme
-
Stachelhäuter 
Schlangenstern
Systematik Reich: Vielzellige Tiere (Metazoa) Abteilung: Gewebetiere (Eumetazoa) Unterabteilung: Bilateria ohne Rang: Neumünder (Deuterostomia) Stamm: Stachelhäuter Wissenschaftlicher Name Echinodermata Stämme - ?Helicoplacoidea †
- ?Arkarua †
- ?Homalozoa †
- Eleutherozoen (Eleutherozoa)
- Asterozoa
- Seesterne (Asteroidea)
- Somasteroidea
- Cryptosyringida
- Seeigel (Echinoidea)
- Seegurken (Holothuroidea)
- Schlangensterne (Ophiuroidea)
- Asterozoa
- Pelmatozoen (Pelmatozoa)
- Seelilien und Haarsterne (Crinoidea)
- Edrioasteroidea†
- Knospenstrahler (Blastoidea) †
- Beutelstrahler (Cystoidea) †
- Eocrinoidea †
- ausgestorben: †
Die Stachelhäuter (Echinodermata) (von griech. ἐχῖνος echinos „Igel“ und δέρμα derma „Haut“) sind ein zu den Deuterostomiern gehörender Tierstamm. Weltweit sind etwa 6.300 rezente Arten der Stachelhäuter bekannt, womit sie die zweitgrößte Tiergruppe innerhalb der Neumundtiere (Deuterostomia) nach den Chordatieren (Chordata) bilden.
Es handelt sich bei ihnen durchweg um Meeresbewohner, die bis auf wenige Tiefseearten reine Bodenbewohner sind - d.h. vorwiegend im Flachmeer leben. Neben ihrer weiten Verbreitung sind sie auch geologisch von Bedeutung, weil ihre abgestorbenen Schalen und Skelettteile wesentlich zur Bildung von Kalkgesteinen beitragen. Bei Vorherrschen einer Skelettart erhalten sie Namen wie Trochiten- oder Crinoidenkalk, die sich unter anderem in den Kalkalpen finden. Mengenmäßig überwiegen aber die Ablagerungen der winzigen, schwebenden Kalkalgen.
Die Echinodermata werden traditionell in die meist gestielten Pelmatozoa sowie die freilebenden Eleutherozoa unterteilt. Zu den Pelmatozoa zählen mehrere als „Seelilien“ zusammengefasste Taxa sowie die sekundär freilebenden Haarsterne (Crinoidea). Die Eleutherozoa umfassen die bekannteren Gruppen der Stachelhäuter: die Seesterne (Asteroidea) und die Schlangensterne (Ophiuroidea), die Seeigel (Echinoidea) sowie die Seewalzen (Holothuroidea) ). In jüngster Zeit (1986) wurde eine weitere Klasse identifiziert, die Seegänseblümchen (Concentricycloidea).
Inhaltsverzeichnis
Allgemeine Merkmale
Die meisten Stachelhäuter erreichen Körpergrößen von weniger als zehn Zentimetern in der Länge oder im Durchmesser. Dabei gibt es allerdings auch sehr große Arten. So können einige Seewalzen Längen von bis zu zwei Metern erreichen und einige Seesterne haben einen Durchmesser von bis zu einem Meter. Die größten Stachelhäuter waren einige ausgestorbene Seelilienarten mit Stammlängen von bis zu 20 Metern. In der Körperform herrschen mit den See- und Schlangensternen sternförmige Tiere vor. Die Seeigel haben meistens eine kugelige Form, die wie bei den Sanddollars auch abgeflacht sein kann, hinzu kommen bei ihnen häufig sehr verschieden gestaltete Stacheln. Eine längliche, manchmal fast schon wurmförmige Gestalt haben die Seewalzen. Die Seelilien wiederum stellen sessile (am Boden angewachsene) Tiere dar, bei denen ein mit vielen Beinen ausgestatteter Körper auf einem langen Stiel sitzt und Nahrungspartikel aus dem Wasser filtert.
Die namensgebenden und so vielfältig ausgeprägten Stacheln sind aus dem Skelett hervorgewachsene, gelenkig bewegliche Hohlformen und sind mit einer Epidermis überzogen. Sie haben mehrere wichtige Funktionen: sie dienen vor allem zum Schutz der Tiere vor Fressfeinden, ferner zum Graben und zur Fixierung am Meeresboden, sowie zur Fortbewegung. Die Stacheln können Gifte enthalten (wie beim Seeigel) und auch Sinnesorgane - zum Beispiel Fotorezeptoren bei den Seesternen. Als Sonderform spezieller Pedicellarien können sie sogar zum Beseitigen von Parasiten oder für den Beutefang verwendet werden.
Aufbau
Obwohl die Stachelhäuter zu den Bilateria und damit zu den bilateralsymmetrischen Tieren gehören, hat sich bei ihnen eine Symmetrieform entwickelt, die auf fünf Symmetrieachsen aufbaut (sekundäre Pentamerie beziehungsweise fünfstrahlige Radiärsymmetrie). Diese ist besonders deutlich bei den fünfarmigen Seesternen, kann jedoch auch bei allen anderen Vertretern der Tiergruppe festgestellt werden. Bei vielen Pelmatozoa und auch bei einigen Seesternen kommt es dabei zu einer Vervielfachung der Fünfstrahligkeit, so dass Seesterne wie Heliaster spec. 50 Arme besitzen können, Seenelken wie Comanthina schlegelii sogar 200. Diese Symmetrieeigenschaften entwickeln sich erst im Laufe der Individualentwicklung (Ontogenese), an deren Anfang eine zweiseitig symmetrische Larve steht. Das Zentrum der Symmetrie bilden fünf Radien, die ein Kanalsystem und Radiärnerven enthalten. Zwischen diesen befinden sich die Interradien.
Zugleich wandern auch der Mund und der After von den ehemaligen Vorder- und Hinterenden der Tiere auf die Ober- und Unterseite. Bei den frei lebenden Eleutherozoa liegt dabei die Mundöffnung an der Unterseite der Tiere (Oralseite) auf dem Substrat, die Afteröffnung entsprechend auf der Oberseite (Aboralseite). Bei den Seewalzen bildeten sich durch die Streckung wieder ein Vorder- und ein Hinterpol mit Mund und After aus, diese entsprechen jedoch der Ober- und Unterseite anderer Stachelhäuter (die Seewalzen liegen also auf der Seite). Bei den Seelilien sind sowohl der Mund als auch der After vom Substrat abgewendet und liegen nebeneinander.
Durchbrochen wird die Symmetrie durch die Madreporen- oder Siebplatte. Diese liegt auf der Aboralseite neben dem After in einem Interradius. Diese Platte legt als sichtbarer Punkt die Nomenklatur des Körpers fest, der ihr gegenüberliegende Radius wird als Radius A bezeichnet und alle folgenden im Uhrzeigersinn von B bis E benannt. Die Madreporenplatte liegt damit immer im Interradius CD.
Außenhaut und Kalkskelett
 Eine Auswahl unterschiedlich gefärbter Seesterne
Eine Auswahl unterschiedlich gefärbter SeesterneAuch wenn der Name Stachelhäuter anderes vermuten lässt, haben die Tiere kein Außenskelett. Die äußerste Schicht wird vielmehr durch eine dünne und einschichtige Epidermis gebildet, die vor allem aus wenig differenzierten Stützzellen mit jeweils einer Cilie bestehen. Dazwischen liegen Drüsenzellen, Mechanorezeptoren und Pigmentzellen. Die Drüsenzellen produzieren dabei Schleim und Klebesekrete und sie kommen entsprechend besonders häufig im Bereich der Saugfüßchen bei Seeigeln, Seesternen und Seewalzen sowie an den Mundtentakeln der Seewalzen vor. Bei einigen Seeigeln, vor allem den Vertretern der Toxopneustidae, können die Drüsenzellen auch Giftstoffe enthalten, die auch beim Menschen als Neurotoxin wirken und nicht ungefährlich sind.
Die Pigmentzellen sind für die Färbung der Tiere verantwortlich. Sie können auf Licht reagieren, sodass es bei vielen Arten zu unterschiedlichen Tages- und Nachtzeichnungen kommt. Diese Reaktion kann sehr schnell erfolgen, so verfärbt sich der Seeigel Centrostephanus longispinus in nur 50 Minuten von einem graubraun in ein tiefschwarz, wenn er plötzlich Licht ausgesetzt wird. Die Farben selbst kommen durch eine Mischung verschiedener Farbstoffe zustande, darunter die dunklen Melanine, rote Carotinoide und Carotinproteine, die blau, grün oder violett sein können. Hinzu kommen Naphtochinone und bei den Diademseeigeln auch noch physikalische Effekte durch reflektierende Iridophoren.
 Sehr ausgeprägte Stacheln eines Seeigels
Sehr ausgeprägte Stacheln eines SeeigelsDas Skelett liegt unterhalb der Epidermis im Mesoderm und besteht aus Calcit mit einem Magnesiumoxidanteil von drei bis fünfzehn Prozent. Da Magnesium-Ionen bei gleicher Ladung kleiner sind als die im Calcit vorherrschenden Calcium-Ionen, bewirken sie eine Erhöhung der Packungsdichte der Calcit-Kristalle und eine Verstärkung der chemischen Bindungen, was wiederum eine Steigerung der Härte und Festigkeit des Materials zur Folge hat. Dieser Effekt wird insbesondere bei den Zähnen der Stachelhäuter ausgenutzt. Durch seine periphere Lage bestimmt das Skelett maßgeblich die Gestalt der Tiere. Das Kalkskelett entsteht dabei nicht durch einfache Abscheidung von Epidermiszellen sondern wird innerhalb eines Zellverbandes von Skelettbildungszellen (Sclerocyten) angelegt. Das entstehende Gebilde, Stereom genannt, ist stark porös, wodurch eine zugleich leichte aber auch feste Konstruktion entsteht. Das Stereom verwächst zu Platten, die als Ossikel bezeichnet werden. Das Wachstum kann in alle Richtungen erfolgen, sodass nach dem Verlust eines Körperteils dieser wieder ersetzt werden kann. Größere Elemente bilden Gelenke im Skelett aus, die durch Muskulatur beweglich werden.
Auch die Stacheln der Seesterne und Seeigel sind mit dem Skelett gelenkig verbunden und können gegen die darunterliegenden Skelettplatten bewegt werden. Die Stacheln selbst sind spezielle Bildungen des Stereoms, die nur aus einer hohl zusammengerollten Skelettplatte bestehen. Wie alle anderen Skelettelemente sind auch sie von Epidermis überzogen. Eine Sonderbildung des Skeletts bei den Seeigeln und Seesternen stellen die pinzettenartigen Pedicellarien dar, die sich wahrscheinlich aus Stacheln entwickelt haben. Sie bestehen aus mehreren Elementen und können zur Abwehr von Parasiten und sich festsetzende Larven anderer Meerestiere sowie zum Beutefang eingesetzt werden. Weitere Skelettelemente sind der Kieferapparat der Seeigel (Laterne des Aristoteles) aus 40 einzelnen Skelettplatten und der Kalkring der Seewalzen, der die Mundöffnung stabilisiert. Bei den Seewalzen ist die Epidermis außerdem mit mikroskopisch kleinen Kalknadeln (Spiculae) gespickt. Auch der Stiel der Seelilien besteht aus einzelnen Skelettelementen, den scheibenförmigen Trochiten.
Ambulacralsystem
Einzigartig ist das innere Kanalsystem (Ambulacralsystem), dessen äußere Fortsätze als Saugfüßchen der Fortbewegung der Tiere oder als Tentakel dem Nahrungserwerb dienen. Es hat sich in der Ontogenese vor allem aus dem Hydrocoel entwickelt und besteht aus einem Ringkanal um den Darm sowie die in die einzelnen Radien führenden Ambulacral- oder Radialkanäle. Auch die Madreporenplatte ist über einen verhärteten Kanal, den Steinkanal, mit dem System verbunden und stellt (außer bei einigen Seewalzen) die Verbindung zum freien Wasser dar.
An den Radialkanälen liegt der Füßchen- und Tentakelapparat der Tiere. Dabei handelt es sich um Fortsätze, die rechts und links vom Radiärkanal angeordnet sind und mit diesem durch einen dünnen Zuleitungskanal in Verbindung stehen. Sie können durch den Kalkpanzer der Tiere nach außen geführt werden und dienen hier dem Nahrungserwerb, dem Gasaustausch, der Fortbewegung, der Exkretion sowie der Osmoregulation. Sie sind bei den verschiedenen Gruppen der Stachelhäuter unterschiedlich aufgebaut. So liegen sie bei den Seelilien und Haarsternen vor allem an den Transportrinnen für die Nahrungsaufnahme, den Pinnulae, und transportieren die Nahrungspartikel zur Mundöffnung. Bei den Seesternen, Seeigeln und Seewalzen bilden sie Füßchen mit einer muskulösen inneren Ampulle, die die Bewegung durch eine Regulation des Flüssigkeitsdrucks steuert.
Weitere Organsysteme
 Seewalze
SeewalzeDer Darm der Stachelhäuter ist sehr einfach aufgebaut. Er zieht bei den Schlangensternen und Seesternen direkt vom Mund zum After, während er bei den Seewalzen und Seelilien im Uhrzeigersinn sowie bei den Seeigeln erst im und dann gegen den Uhrzeigersinn gewunden ist. Er wird durch Mesenterien des Somatocoel in seiner Position gehalten. Im Regelfall lässt sich der Darm in einen Mundbereich mit Schlund (Oesophagus), einen Magen, einen inneren Darmbereich und einen Rektalbereich mit After unterteilen. Durch Zotten des inneren Darmes wird die Oberfläche vergrößert, außerdem existieren Bereiche mit Spezialfunktionen wie etwa die Rektalkiemen der Seewalzen oder ebenfalls der Atmung dienende Siphone bei grabenden Seeigeln.
Das Blutgefäßsystem oder besser Hämalsystem besteht aus einem zentralen Ring (Hämalring) mit fünf Kanälen entlang der Ambulacralkanäle. Ein weiterer Kanal zieht zum Axialorgan, welches die Verbindungsstelle zwischen dem Hämalsystem und dem Ambulacralsystem darstellt und als kontraktives Organ die Flüssigkeitszirkulation regelt. Die Exkretion erfolgt über das Ambulacralsystem.
Das Nervensystem der Stachelhäuter stellt im Wesentlichen ein komplexes Nervennetz unterhalb der Epidermis und um den Darm dar. Um den Schlund liegt ein Nervenring, von dem außerdem Nerven entlang der Radialkanäle führen. Stachelhäuter besitzen nur sehr wenige Sinnesorgane. Vor allem die Epidermis reagiert durch eingelagerte Sinneszellen auf Licht, Temperatur, Bewegungs- und Strömungsrichtung. Auch die Pedicellarien und Stacheln reagieren auf verschiedene Reize. Besonders bei den Seesternen haben sich augenähnliche Photorezeptoren an den Enden der Arme aus Ambulacralfüßchen entwickelt; ähnliche Sinneszellen finden sich aber auch bei Seeigeln und Seewalzen.
Die Gonaden füllen vor allem bei den Seeigeln und Seewalzen die gesamte Leibeshöhle aus, bei den Seelilien und Seesternen liegen sie in den Armen, bei den Seesternen jeweils zwei aufgefächerte Gonaden pro Arm, die zwischen den Armen an jeweils einer Geschlechtsöffnung enden. Seeigel haben zwischen drei und fünf Gonaden, die an den Interradien befestigt sind und nach außen münden. Bei den Seewalzen existiert nur eine Gonade, die sich im Körper verzweigt und in einem Geschlechtsgang mit Geschlechtsöffnung am Hinterende mündet. Da viele ausgestorbene Stachelhäuter ebenfalls nur eine Geschlechtsöffnung hatten, wird dieser Zustand als ursprünglich angesehen.
Fortpflanzung und Entwicklung
Sexuelle Fortpflanzung und Brutpflege
 Haarstern Ptilometra australis)
Haarstern Ptilometra australis)Stachelhäuter werden nach etwa zwei bis drei Jahren geschlechtsreif, abhängig von der Art und den Umweltbedingungen. Die Eier und Spermien werden in das freie Wasser abgegeben, wo die Befruchtung stattfindet. Dies geschieht bei einigen Arten in dichten Populationen zeitlich aufeinander abgestimmt. Eine Paarung mit einer inneren Befruchtung konnte bislang nur bei drei Schlangensternen, drei Seesternen und einer Tiefsee-Seewalze beschrieben werden.
Bei einigen Haarsternen entwickeln sich die Embryonen in speziellen Bruttaschen (Marsupien), bei diesen gelangen nur die Spermien ins freie Wasser und befruchten die Eier in den Bruttaschen. Auch bei einigen Seeigeln und Seesternen kommt eine Brutpflege vor. So tragen verschiedene Lanzenseeigel die Jungtiere zwischen den Stacheln der Oralseite, Herzseeigel haben speziell ausgebildete Brutkammern. Bei Seesternen können spezielle Kammern im Bereich der Atemkammern an der Aboralseite oder Magentaschen ausgebildet sein, in denen die Jugendentwicklung stattfindet. Bei einigen Seewalzen findet Brutpflege in Bereichen der Kriechsohle, zwischen den Tentakeln oder sogar in der Leibeshöhle statt. Die meisten Stachelhäuter betreiben allerdings keine Brutpflege.
Larvalentwicklung
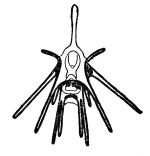 Pluteus eines Seeigels
Pluteus eines SeeigelsDen Beginn der Entwicklung stellt eine Radiärfurchung dar mit annähernd äqualer Teilung. Es entwickelt sich erst eine Coeloblastula, aus der durch Einstülpung eine Gastrula wird. In den Hohlraum wandern Mesodermzellen ein, die später Skelettelemente bilden. Die sekundäre Leibeshöhle (Coelom) bildet sich durch Abschnürung von drei paarigen Blasen (Enterocoelie).
Mit der Metamorphose bilden sich für die verschiedenen Taxa der Stachelhäuter unterschiedliche typische Larvenformen. Dabei handelt es sich bei den Pelmatozoa immer um die dotterreichen Doliolaria- oder Vitellaria-Larven, die nach ihrem Festsetzen ein fressfähiges Pentacrinusstadium bilden, aus der sich später das fertige Tier entwickelt. Die Grundlarvenform aller Eleutherozoa ist die Dipleurula-Larve, die sich bei den Seesternen über eine Bipinnaria-Larve und eine Brachiolaria-Larve zum adulten Tier entwickelt. Bei den Seegurken ist die erste Larvenform die Auricularia-Larve mit einem einheitlichen Wimpernband, aus der sich dann die Doliolaria mit drei bis fünf Wimpernringen entwickelt. Die auffälligsten Larven bilden die Seeigel und die Schlangensterne. Dabei handelt es sich um Pluteus-Larven mit langen Schwebestacheln.
In der Larvalentwicklung kommt es zur bereits angesprochenen Metamorphose, bei der aus den anfänglichen bilateralsymmetrischen Larven die fünfstrahligen Tiere entstehen. Den wesentlichen Teil dabei bildet das ursprünglich in drei paarigen Hohlräumen angelegte Coelom. Dieses entsteht durch eine beidseitige Abschnürung im Urdarmdach, also durch die bei den Deuterostomia übliche Enterocoelie, als Bläschen. Die Bläschen wachsen sich aus und teilen sich in Richtung Hinterpol in drei Abschnitte. Dabei bildet das Protocoel den obersten, das Mesocoel den mittleren und das Metacoel den hintersten Abschnitt. Das linke und das rechte Metacoel lagern sich als Somatocoel übereinander und bilden die eigentliche Leibeshöhle, die den Darm umgibt und die Geschlechtsdrüsen enthält. Aus dem linken Protocoel entwickelt sich das Axocoel, welches über einen Kanal und die Madreporenplatte nach außen mündet. Dabei bildet sich aus dem linken Mesocoel als Hydrocoel ein Ringkanal als Zentrum des Ambulacralsystems, der sich um den Darm legt und mit dem Axocoel über den Steinkanal verbunden ist. Aus dem Ringkanal wachsen Radiärkanäle in die späteren Radien ein. Das rechte Hydrocoel verschwindet vollständig und das rechte Axocoel wird auf eine kleine Dorsalblase reduziert. Bei den meisten Tieren bildet sich entsprechend die Oralseite auf der linken Körperseite. Abänderungen dieser Entwicklung gibt es bei den Seelilien, Haarsternen und den Seewalzen.
Regeneration und asexuelle Fortpflanzung
Sehr viele Stachelhäuter können verlorene Körperteile wieder neu anlegen. Seesterne und Schlangensterne schnüren außerdem aktiv Körperteile ab (Autotomie), wenn sie in Gefahr sind. Die Seegurken stoßen Teile der inneren Organe aus und Seeigel verlieren ständig Pedicellarien oder Stacheln. Das Regenerationsvermögen für alle diese Körperteile ist sehr gut ausgeprägt.
Die Regenerationsfähigkeit erlaubt den Tieren auch eine Form der ungeschlechtlichen Fortpflanzung, die als Fissiparie bekannt ist. Seesterne vermehren sich dabei etwa über den Verlust von Armen, wobei aus jedem Armteil ein neuer Seestern entstehen kann. Bei einigen Arten, etwa den Coscinasterus-Arten, kommt es durch Fissiparie zu einer Vermehrung der Arme, wobei keine neuen Tiere entstehen. Einige Schlangensterne vermehren sich in manchen Populationen offensichtlich über lange Zeiträume ausschließlich fissipar.
Verbreitung und Lebensraum
 Seewalze Stichopus fuscus vor der Küste der Galápagos-Insel Champion
Seewalze Stichopus fuscus vor der Küste der Galápagos-Insel ChampionStachelhäuter sind weltweit in allen Meeren und Ozeanen zu finden, einige Arten leben auch im Brackwasser. Dabei sind sie regional sehr häufig und stellen in vielen Bereichen die häufigsten Bewohner des Meeresbodens dar. Die verschiedenen Arten sind meistens spezifisch an bestimmte Umweltbedingungen angepasst und kommen entsprechend nur regional begrenzt vor. So gibt es unter den Stachelhäutern sehr viele Arten, die ausschließlich in den Tropen zu finden sind, andere leben in den polaren Gewässern der Arktis oder der Antarktis. Unter den tropischen Arten findet man viele, die immer an Korallenriffe gebunden sind, jedoch in allen Riffen im gesamten Indischen Ozean und Pazifischen Ozean vorkommen.
Die meisten Stachelhäuter leben vollständig auf dem Meeresboden und auf anderen, ähnlichen Strukturen wie Felsstrukturen oder Korallenriffen. Viele Arten graben sich dabei auch in den Meeresboden ein. Nur wenige Seewalzenarten der Tiefsee sind freischwimmend und leben in den unteren Meeresschichten. Dabei sind alle Formen bis auf die sessilen Seelilien sehr mobil und können aktiv weite Strecken zurücklegen. Besonders die Seeigel und die Schlangensterne nutzen zur Verbreitung auch ihre freischwimmenden Larven, die eine weitere Streuung ermöglichen. Ihre Tiefenverbreitung ist wiederum artabhängig. Einige Schlangensterne leben zu tausenden in den Röhrensystemen von tropischen Schwämmen. Auch einige Seewalzen leben an anderen Organismen, etwa zwischen den Stacheln großer Seeigel in der Antarktis. Eine Seewalzenart befestigt ihren Körper sogar an der Haut von Tiefseefischen und lässt sich auf diese Weise transportieren.
Die Arten der Flachwasserzonen und Meeresküsten kommen dabei selten in größeren Tiefen als 100 Meter vor. Tiefseearten finden sich zwischen 1000 bis 5000 Metern Tiefe. Zu den Tiefseearten gehören auch die Seegänseblümchen, die ausschließlich in untergetauchtem Holz in der Tiefsee gefunden wurden. Aus Gebieten mit 10.000 Metern Tiefe und mehr sind bislang ausschließlich Seewalzenarten bekannt. Diese stellen hier in einigen Bereichen bis zu 90 Prozent aller lebenden Organismen dar.
Lebensweise
Ernährung
Die Ernährungsgewohnheiten innerhalb der Stachelhäuter sind sehr vielfältig. Das Spektrum reicht dabei von den Seelilien, die Nahrungspartikel aus dem sie umgebenden Wasser ausstrudeln bis hin zu aktiven Jägern unter den Seesternen. Die meisten Stachelhäuter fressen vor allem nachts, viele nehmen jedoch auch kontinuierlich Nahrung auf.
Die Seelilien und die Haarsterne gehören zu den Filtrierern, die die Nahrung aus dem sie umgebenden Wasser mit Hilfe von Siebstrukturen ausfiltern. Diese werden durch die Arme der Tiere gebildet, die häufig in großer Anzahl vorhanden sind. Die Nahrungspartikel, vor allem Planktonorganismen bleiben dabei in einem Schleim hängen, der von den Ambulacralfüßchen gebildet und abgegeben wird. Über Rinnen in den Armen fließt der Schleim, aktiv durch Cilien und Füßchen transportiert, zur zentralen Mundöffnung.
Die meisten Seeigel sind Alles- oder Pflanzenfresser und weiden mit ihren scharfen Zähnen Algen und andere Nahrungsstoffe von Steinen des Meeresbodens ab. Andere Arten, wie etwa die zahnlosen Irregularia, graben im Erdboden und greifen kleinere Organismen mit ihren Peducelarien. Besonders die Sanddollars nehmen Schwebstoffe auf, die durch die Wasserbewegung über sie transportiert werden.
 Schlangenstern bei der aktiven Jagd nach Eiern von Korallen
Schlangenstern bei der aktiven Jagd nach Eiern von KorallenSeewalzen nutzen verschiedene Arten der Nahrungsaufnahme. Die meisten Arten schlucken einfach große Mengen Meeresboden und entnehmen diesem organische Bestandteile während sie die Reste wieder ausscheiden. Einige andere Arten suchen mit ihren Mundtentakeln aktiv nach Nahrung, die sie vom Meeresboden aufnehmen können. Eine dritte Gruppe nutzt ihre Mundtentakel als Netz, in dem sich schwimmende Organismen verfangen.
Auch unter den Seesternen gibt es Arten, die Schlamm fressen und die organischen Bestandteile entnehmen. Viele Arten sind jedoch auch aktive Jäger und jagen vor allem Muscheln und andere Seesterne, deren Schalen sie mit ihren kräftigen Armen öffnen. Sie stülpen häufig ihren gesamten Magen in die Beute und verdauen diese außerhalb des Körpers. Schlangensterne fangen meistens aktiv kleine Organismen aus dem Wasser, andere besitzen stark aufgefächerte Arme und filtern ähnlich den Seelilien Nahrung aus dem Wasser.
Feinde und Verteidigung
Obwohl die Stachelhäuter durch ihre Panzerung und ihre Stacheln vor Feinden gut geschützt sind, gibt es doch eine ganze Reihe von Prädatoren, die sich von ihnen ernähren. Zu den Fressfeinden gehören Steinfische, Bodenhaie und andere Fische, außerdem verschiedene Krebse, Meeresschnecken und vor allem größere Seesterne, die kleinere Verwandte und Schlangensterne fressen. Vor allem Seeigel werden außerdem von Meeresvögeln wie den Eiderenten sowie von Seeottern, Polarfüchsen und schließlich auch Menschen gejagt und gefressen.
Den Hauptschutz der Stachelhäuter stellen die bereits erwähnten Stacheln der Seeigel sowie ihr hartes Skelett dar. Hinzu kommen bei einigen Seeigeln Gifte, die sie über ihre Pedicellarien abgeben können. Viele Seewalzenarten stoßen bei Gefahr ein Gewirr von klebrigen Fäden aus, die Cuvierschen Schläuche, andere produzieren mit den Holothurinen starke Gifte.
Fortbewegung
.jpg) Seeigel während der Fortbewegung mithilfe seiner Ambulacralfüßchen
Seeigel während der Fortbewegung mithilfe seiner Ambulacralfüßchen Unterseite eines Seesterns mit deutlich erkennbaren Ambulacralfüßchen
Unterseite eines Seesterns mit deutlich erkennbaren AmbulacralfüßchenSeesterne und Seeigel bewegen sich vor allem durch ihre Ambulacralfüßchen sowie durch die Stacheln fort. Sie sind dadurch in der Lage, in jede Richtung zu wandern. Diese Füßchen werden auch als Saugnäpfchen eingesetzt, wodurch die Tiere auch an senkrechten Strukturen klettern können. Die Schlangensterne nutzen ihre Arme zur Fortbewegung, indem sie sie schlängelnd bewegen und in die Bewegungsrichtung schlagen. Auch die Seewalzen nutzen die Ambulacralfüßchen zur Fortbewegung. Dabei wandern sie meistens in Richtung des Vorderendes und unterstützen die Bewegung durch Kontraktionen ihres Körpers. Die Synaptidae verankern sich außerdem mit ihren klebrigen Tentakeln und ziehen sich dadurch vorwärts.
Die Seelilien bewegen sich in der Regel gar nicht fort, obwohl es bei ihnen auch Arten gibt, die sich von ihrem Fixpunkt lösen und an anderer Stelle erneut festsetzen können. Die Haarsterne schwimmen, indem sie ihre Arme auf- und abwärts schlagen, doch auch sie sind meistens ortstreu. Eine schwimmende Fortbewegung kennt man außerdem bei einigen Schlangensternen und vor allem bei speziell abgeflachten Seewalzen der Tiefsee. Diese haben meist einen gelatinösen Körper und besitzen kein Skelett mehr.
Sehr viele Arten der Stachelhäuter graben im Boden und bewegen sich entsprechend auch grabend fort. Dabei nutzen die Seeigel ihre Stacheln, Pedicellarien und Ambulacralfüßchen zum Graben, einige Arten haben außerdem die Möglichkeit, sich durch ihre Zähne und Stacheln in Felsgestein zu bohren. Die Seewalzen graben sich mit Hilfe ihrer Tentakel sowie kontrahierenden Bewegungen ein. Seeigel und Schlangensterne nutzen ihre Arme zum Graben.
Die meisten Stachelhäuter haben eine Lebendposition, bei der die Mundöffnung zum Boden weist. Werden sie umgedreht, so versuchen sie durch Streckungen und Verdrehungen, die ursprüngliche Position wieder einzunehmen.
Aggregation
Bei vielen Stachelhäutern kommt es zur Bildung von großen Ansammlungen, vor allem bei verschiedenen Seeigeln und Schlangensternen. Dieses Phänomen ist bereits fossil überliefert, da viele fossile Formen in großen Fossillagerstätten aus einer oder wenigen Arten gefunden werden. Diese Ansammlungen lassen sich vor allem auf verschiedene Umweltbedingungen zurückführen, die den Tieren optimale Lebensbedingungen bieten. Dabei spielt vor allem die Verteilung der Nahrung eine große Rolle, so finden sich große Ansammlungen von Schlangensternen häufig in Bereichen mit hohem Planktonvorkommen.
Ökologische Rolle und Gefahr
 Schlüsselloch-Seeigel
Schlüsselloch-SeeigelStachelhäuter leben auf der einen Seite von organischem Bestandsabfall auf dem Meeresboden und von Pflanzenteilen, auf der anderen stellen sie effektive Räuber kleinerer Tiere dar. Damit sorgen sie für den Abbau von organischem Material und für die Regulation der Kleintierpopulationen. In Korallenriffen kontrollieren verschiedene Seesterne das Wachstum von Algenmatten und verhindern damit ein Zuwachsen der Korallenstöcke. Seewalzen sorgen für eine Veränderung des Meeresbodens ähnlich wie dies Regenwürmer auf dem Land tun. Sie fressen große Mengen Substrat und scheiden dieses nach der Darmpassage wieder aus. Dadurch wird das Substrat von organischen Stoffen befreit und zugleich chemisch und physikalisch verändert. Durch Grabtätigkeit durchmischen verschiedene grabende Seesterne, Sanddollars und Herz-Seeigel den Boden in Tiefen bis etwa 30 Zentimeter, Bohrseeigel bohren sogar Löcher in feste Gesteine.
Die großen Mengen von Larven der Stachelhäuter stellen außerdem eine reichhaltige Futterquelle für andere Tierarten dar. Auch die ausgewachsenen Stachelhäuter werden von verschiedenen Tieren gefressen, etwa von Seeottern, Meeresvögeln und einigen Fischen. Einige Stachelhäuter, vor allem Seewalzen, stellen den Lebensraum für andere Tiere dar. So leben in den Körpern verschiedene Würmer, aber auch Krabben und Meeresschnecken. Besonders bekannt sind verschiedene Grundeln, die im Darm von Seewalzen leben und den After als Eingangsloch ihrer „Höhlen“ nutzen. Die meisten dieser Bewohner sind Kommensalen, einige leben jedoch auch parasitisch.
Vor allem, wenn die Tiere überhand nehmen, kann ihr Fraßverhalten allerdings zu einem Ungleichgewicht führen. So führen beispielsweise die Massenbestände von Seesternen in einigen Kelpwaldregionen zu einem Absterben der großen Tangpflanzen, andere Arten ernähren sich von Korallenpolypen und führen bei Massenauftreten zum Absterben von ganzen Riffen.
Evolution
 Fossile Seelilienkronen
Fossile SeelilienkronenErste sichere Funde von Stachelhäutern sind bereits seit dem frühen Kambrium von über 540 Millionen Jahren bekannt. Allerdings gibt es auch Fossilien aus dem späten Präkambrium (Arkarua, Tribrachidium), die häufig dieser Tiergruppe zugeordnet werden. Innerhalb einer ersten Radiation im Kambrium entstanden mehrere heute nicht mehr existierende Großgruppen. Die Helicoplacoidea, spiralig aufgebaute Tiere, starben bereits im frühen Kambrium wieder aus. Erfolgreicher sind die asymmetrischen Carpoidea, die in mehreren wahrscheinlich nicht näher verwandten Taxa existierten. Von den vier Taxa überdauerten zwei das Kambrium, auch diese verschwinden jedoch im Laufe des Paläozoikums. Nicht eingeordnet wurden bislang die Vertreter der Gattungen Peridionites und Cymbionites, die während des mittleren Kambriums lebten.
Mit den wenig erfolgreichen Gruppen des Kambriums entstanden auch die bereits fünfstrahligen Eocrinoidea und Edrioasteroidea, aus denen sich die modernen Vertreter der Stachelhäuter ableiten lassen. Diese stellen die Basis für eine weitere Radiation der Stachelhäuter im unteren Ordovizium dar. Die Eocrinoidea zeichneten sich durch eine aufwärts gerichtete Mundöffnung sowie den Erwerb von Körperanhängen in Form von Brachiolen (selten echten Armen) aus. Innerhalb dieser Gruppe entwickelten sich die Cystoidea und die Blastoidea sowie die Parablastoidea und die Paracrinoidea. Ob die heute noch existierenden Seelilien und Haarsterne ebenfalls auf diese Gruppe zurückgehen ist nicht bekannt.
Die Edrioasteroidea besaßen keine Verankerungsorgane und auch keine Brachiolen, bei ihnen kam es allerdings zu einer Umorientierung der Mundöffnung auf die Unterseite der Tiere während der After auf die Oberseite wanderte. Diese Tiere bildeten die Wurzel der Eleutherozoa, zu denen auch die heute noch lebenden Seesterne, Schlangensterne, Seeigel und Seewalzen gehören.
Systematik
In der klassischen Systematik der Stachelhäuter hat sich in den 1980er Jahren eine Einteilung etabliert, die von der ursprünglichen Kategorisierung der Tiere in die vorwiegend festsitzenden Pelmatozoa und die frei beweglichen Eleutherozoa abweicht. Das Hauptargument gegen eine solche Einteilung stellt der fehlende Bezug zu den ausgestorbenen Gruppen dar, bei denen beide Lebensweisen in den verschiedenen Gruppen sowohl bei den Eleutherozoa als auch den Pelmatozoa zu finden sind. Die folgende Systematik teilt die fossilen und rezenten Stachelhäuter entsprechend in mehrere Unterstämme auf:
- Stamm Stachelhäuter (Echinodermata)
- Unterstamm Homalozoa † - Mittleres Kambrium bis Mittleres Devon (570–365 Millionen Jahre)
- Klasse Stylophora † - Mittleres Kambrium bis Oberes Ordovizium (540–460 Millionen Jahre)
- Klasse Homostelea † - Mittleres Kambrium (540 Millionen Jahre)
- Klasse Homoiostelea † - Oberes Kambrium bis Unteres Devon (510–400 Millionen Jahre)
- Klasse Ctenocystoidea † - Mittleres Kambrium (540 Millionen Jahre)
- Unterstamm Blastozoa † - Kambrium bis Perm (570–280 Millionen Jahre)
- Klasse Eocrinoidea † - Unteres Kambrium bis Silur (570–430 Millionen Jahre)
- Klasse Knospenstrahler (Blastoidea) † - Silur bis Perm (430–280 Millionen Jahre)
- Klasse Paracrinoidea † - Mittleres Ordovizium (460 Millionen Jahre)
- Klasse Parablastoidea † - Unteres bis Mittleres Ordovizium (500–400 Millionen Jahre)
- Klasse Rhombifera † - Unteres Ordovizium bis Oberes Devon (500–350 Millionen Jahre)
- Klasse Diploporita † - Unteres Ordovizium bis Unteres Devon (500–400 Millionen Jahre)
- Unterstamm Crinozoa - seit dem Unteren Ordovizium (500 Millionen Jahre bis heute)
- Klasse Seelilien und Haarsterne (Crinoidea) - seit dem Unteren Ordovizium (500 Millionen Jahre bis heute)
- Unterstamm Asterozoa - seit dem Unteren Ordovizium (500 Millionen Jahre bis heute)
- Klasse Somasteroidea † - Unteres Ordovizium bis Oberes Devon (500–350 Millionen Jahre)
- Klasse Seesterne (Asteroidea) - seit dem Mittleren Ordovizium (460 Millionen Jahre bis heute)
- Klasse Schlangensterne (Ophiuroidea) - seit dem Mittleren Ordovizium (460 Millionen Jahre bis heute)
- Klasse Seegänseblümchen (Concentricycloidea) - nur rezent bekannt
- Unterstamm Echinozoa - seit dem Unteren Kambrium (570 Millionen Jahre bis heute)
- Klasse Cyclocystoidea † - Mittleres Ordovizium bis Mittleres Devon (460–375 Millionen Jahre)
- Klasse Edrioasteroidea † - Unteres Kambrium bis Oberes Karbon (570–340 Millionen Jahre)
- Klasse Helicoplacoidea - Unteres Kambrium (570 Millionen Jahre)
- Klasse Ophiocistioidea † - Unteres Ordovizium bis Oberes Silur (500–395 Millionen Jahre)
- Klasse Seewalzen (Holothuroidea) - seit dem Ordovizium (460 Millionen Jahre bis heute)
- Klasse Seeigel (Echinoidea) - seit dem Ordovizium (460 Millionen Jahre bis heute)
- Unterstamm Homalozoa † - Mittleres Kambrium bis Mittleres Devon (570–365 Millionen Jahre)
In der phylogenetischen Systematik werden die Seelilien und Haarsternen (Crinoidea) mit etwa 620 Arten allen anderen Gruppen (Eleutherozoa) als Schwestergruppe gegenüber gestellt. Die Seesterne (Asteroida) mit etwa 1500 Arten sowie die erst 1986 entdeckten Seegänseblümchen (Concentricyclida) mit bislang 2 bekannten Arten stehen innerhalb der Eleutherozoa dem gemeinsamen Taxon aus Seeigel (Echinoida) mit etwa 950 Arten, Seewalzen (Holothuroida) mit etwa 1200 Arten und den Schlangensternen (Ophiuroida) mit etwa 2000 Arten gegenüber.
--- Stachelhäuter (Echinodermata) |-- Crinoidea | |-- "Seelilien": 4 Taxa, Systematik ungeklärt. | '-- Haarsterne (Comatulida) | '-- Eleutherozoa |-- N.N. | |-- Seesterne (Asteroida) | '-- Seegänseblümchen (Concentricycloida) | '-- Cryptosyringida |-- Schlangensterne (Ophiuroida) '-- Echinozoa |-- Seeigel (Echinoidea) '-- Seewalzen (Holothuroida)Stachelhäuter und Menschen; Wirtschaft
Die meisten Menschen kennen die Stachelhäuter eher von der unangenehmen Seite: wenn man sich in Küstennähe - z.B. bei Felsen oder einem Riff - viele giftige Stacheln eines Seeigels eintritt. Wegen ihres feinen, ringförmigen Aufbaus brechen sie beim Entfernen meist ab und brauchen Geduld (Herauseitern) oder den Arzt. Für die Küche einiger Länder sind Stachelhäuter jedoch besondere Delikatessen - und für Kinder ihre Skelette ein beliebtes Sammelobjekt.
 Orangefarbene Eier des Seeigels
Orangefarbene Eier des SeeigelsDie wirtschaftliche Bedeutung der Stachelhäuter ist vornehmlich lokal. So werden pro Jahr etwa 50.000 Tonnen Seeigel gefangen, von denen besonders in Japan, Peru und in Frankreich die Gonaden bzw. die Eier gegessen werden. In Japan sind die Eier als Uni bekannt und stellen eine Zutat der Sushi-Küche dar. Dabei wird der Geschmack als weich und schmelzend beschrieben. Die Qualität ist abhängig von der Farbe, die von hellgelb bis leuchtend orange reichen kann.
Auch Seewalzen stellen in einigen Ländern Südostasiens eine Delikatesse dar. Am bekanntesten sind sie als Trepang oder bêche-de-mer in China und Indonesien. Die Seewalzen werden dafür von den (häufig giftigen) Eingeweiden befreit und getrocknet. Besonders beliebt sind die Ananas-Seewalze (Thelenota ananas) (susuhan) und die rote Halodeima edulis. Die starken Giftstoffe der Seewalzen, bekannt als Holothurine, sind leicht psychoaktiv, Nutzungen der entheogenen Wirkungen sind jedoch nicht bekannt. Allerdings konnte festgestellt werden, dass Holothurine hemmend auf die Wachstumsrate verschiedener Tumorzellen wirkt, wodurch es für die Krebsmedizin interessant wird.
Seesterne gelangen gemahlen als Kalkzusatz in industrielles Tierfutter, jährlich werden für diesen Zweck etwa 4000 Tonnen der Tiere verwendet. Vor allem in Gebieten, in denen kommerziell Muscheln, vor allem Austern, gezüchtet werden, können Massenauftreten von Seesternen einen großen wirtschaftlichen Schaden anrichten.
Literatur
- D. T. Anderson: Invertebrate Zoology. Oxford University Press, Oxford 2001. ISBN 0-19-551368-1
- R. S. K. Barnes, P. Calow, P. J. W. Olive, D. W. Golding, J. I. Spicer: The invertebrates - a synthesis. Blackwell, Oxford 2001, S.151. ISBN 0-632-04761-5
- R. C. Brusca, G. J. Brusca: Invertebrates. Sinauer, Sunderland Mass 2003. ISBN 0-87893-097-3
- H. Fechter, E. Thenius: Die Stachelhäuter. In: Grzimeks Tierleben. Kindler, Zürich 1962.
- A. Goldschmid: Echinodermata, Stachelhäuter. In: W. Westheide, R. Rieger: Spezielle Zoologie. Teil 1. Einzeller und Wirbellose Tiere. Spektrum, Heidelberg 1996, 2003. ISBN 3-8274-0998-5
- J. Moore: An Introduction to the Invertebrates. Cambridge University Press, Cambridge 2001. ISBN 0-521-77914-6
- E. E. Ruppert, R. S. Fox, R. P. Barnes: Invertebrate Zoology. A functional evolutionary approach. Brooks/Cole, Belmont 2004. ISBN 0-03-025982-7
Weblinks







.jpg)



Wikimedia Foundation.