- Isidien
-
Dieser Artikel erläutert die Lebensform Flechte; zu der Hauterkrankung siehe Lichen ruber planus, Lichen pilaris und Lichen sclerosus. - Krustenflechten: Der Thallus liegt dicht auf dem Untergrund oder durchwächst diesen. Man kann sie nicht vom Substrat ablösen, ohne die Flechte zu beschädigen oder zu zerstören. Diese Wuchsform ermöglicht es, auf exponierten Felsflächen in den sehr hohen Gebirgslagen oder auch auf Betonmauern zu gedeihen. Die Wachstumszone liegt am äußeren Rand der Flechte.
- Laub- oder Blattflechten: Die Flechte ist flächig gestaltet (folios) und liegt mehr oder weniger locker auf dem Substrat auf. Morphologisch sind die Blattflechten sehr vielfältig und besiedeln verschiedene Lebensräume wie etwa auf Moosen, aber auch auf Gestein. Wie bei Pflanzenblättern optimiert der blattartige Wuchs die Lichtausbeute für die Photosynthese des Photobionten. Die Wachstumszone befindet sich auf den „Blatträndern“.
- Strauchflechten: Der Thallus ist strauchförmig und wächst als aufrechter Rasen auf Erde oder Fels oder hängt von Bäumen, Totholz oder Felsen (Bart- oder Bandflechten). Die Wachstumszone liegt am Ende der einzelnen Äste.
- Gallertflechten: Dies sind Flechten mit Cyanobakterien als Partner, die bei Befeuchtung gallertartig aufquellen und meist schwärzlich bis dunkeloliv gefärbt sind[1].
- Apothecien sind gewöhnlich vom Flechtenlager scharf abgegrenzte, rundliche bis scheiben- oder schüsselförmige Gebilde. Darauf beziehungsweise darin liegt eine aus parallel gelagerten Asci und nichtsporenbildenden Hyphenenden bestehende Schicht, das so genannte Hymenium, offen zu Tage.
- Perithecien sind mehr oder weniger kugelige, fast geschlossene Gebilde, in denen sich die Asci befinden und die Ascosporen gebildet werden, die nur durch eine Pore austreten können[4].
- Isidien sind Auswüchse in Form von Stiften, Knöpfen, Blättchen oder kleinen Ästen, die an der Basis eine Sollbruchstelle haben. Durch Wind, Wasser oder leichte Berührungen brechen diese ab und bilden auf einem geeigneten Untergrund eine neue Flechte.
- Sorale sind staubige Aufbrüche der Flechten, aus der Körnchen aus wenigen verflochtenen Pilzfäden und Algen heraustreten (Soredien). Durch Verbreitung dieser Körnchen können ebenfalls neue Flechten gebildet werden[5].
- „Und das Moos, das an gewissen Bäumen wächst, hat Heilkraft in sich. Und solches, das auf fauligen Hälzern wächst, hat fast keine heilkraf, weil die in stinken Säften der Dächer und fauliger Hölzer und in Steinen vorhanden ist, ausbricht und im Moos auswächst, daher ist es fast ohne Nutzen.“[9].
- ↑ Heribert Schöller: Flechten - Geschichte, Biologie, Systematik, Ökologie, Naturschutz und kulturelle Bedeutung. Schweizerbartsche Verlagsbuchhandlung, Frankfurt am Main 1997, ISBN 3-7829-1151-2, S. 21-28
- ↑ Volkmar Wirth: Die Flechten Baden-Württembergs. Verlag Eugen Ulmer, Stuttgart 1995, ISBN 3-8001-3325-3, S. 33
- ↑ Volkmar Wirth: Die Flechten Baden-Württembergs. Verlag Eugen Ulmer, Stuttgart 1995, ISBN 3-8001-3325-3, S. 40
- ↑ Heribert Schöller: Flechten - Geschichte, Biologie, Systematik, Ökologie, Naturschutz und kulturelle Bedeutung. Schweizerbartsche Verlagsbuchhandlung, Frankfurt am Main 1997, ISBN 3-7829-1151-2, S. 35-37
- ↑ Bernhard Marbach, Christian Kainz: Moose, Farne und Flechten. Häufige und auffällige Arten erkennen und bestimmen. BLV Verlagsgesellschaft, München 2002, ISBN 3-405-16323-4, S. 15
- ↑ Bernhard Marbach, Christian Kainz: Moose, Farne und Flechten. Häufige und auffällige Arten erkennen und bestimmen. BLV Verlagsgesellschaft, München 2002, ISBN 3-405-16323-4, S. 17
- ↑ http://www.uni-muenster.de/GeoPalaeontologie/Palaeo/Palbot/nature.html
- ↑ Heribert Schöller: Flechten - Geschichte, Biologie, Systematik, Ökologie, Naturschutz und kulturelle Bedeutung. Schweizerbartsche Verlagsbuchhandlung, Frankfurt am Main 1997, ISBN 3-7829-1151-2, S. 3-10
- ↑ Heribert Schöller: Flechten - Geschichte, Biologie, Systematik, Ökologie, Naturschutz und kulturelle Bedeutung. Schweizerbartsche Verlagsbuchhandlung, Frankfurt am Main 1997, ISBN 3-7829-1151-2, S. 3-10, S. 189
- Wirth, Volkmar: Flechten erkennen - Luftgüte bestimmen. Verlag Eugen Ulmer, Stuttgart 1997, ISBN 3-8001-3486-1
- Kirschbaum, Ulrich & Düll, Ruprecht: Farbatlas Flechten und Moose. Verlag Eugen Ulmer, Stuttgart 2000, ISBN 3-8001-3517-5
- Jahns, Hans Martin: BLV Bestimmungsbuch: Farne, Moose, Flechten, München 1995, ISBN 3-405-13458-7
- Wirth, Volkmar: Die Flechten Baden-Württembergs. Ulmer, Stuttgart 1987, ISBN 3-8001-3305-9
- Wirth, Volkmar: Flechtenflora. Verlag Eugen Ulmer, Stuttgart, ISBN 3-8252-1062-6
- Reckel, Sylvia, Aöschner, Manfred & Marion Stock: Flechten als Anzeiger der Luftqualität. In: Biologie in unserer Zeit. Bd. 29, Nr. 6, 1999, S. 364-370, ISSN 0045-205X
- Rote Liste der gefährdeten Arten der Schweiz (PDF)
- Umfassende Website zu Flechten (englisch)
- Flechten Belgiens, Luxemburgs und Nordfrankreichs (englisch)
- Checklists of Lichens (englisch)
- Chilenische Flechten (spanisch)
- Datenbank der Nutzbarkeit von Flechten für den Menschen (englisch)
- Fossile Flechten (deutsch)


Als Flechte (Lichen) bezeichnet man eine symbiotische Lebensgemeinschaft zwischen einem Pilz, dem so genannten Mykobionten, und einem oder mehreren Photosynthese betreibenden Partnern. Diese Photobionten, auch Phytobionten genannt, sind Grünalgen (Chlorophyta) oder Cyanobakterien. Die Eigenschaften der Flechten setzen sich deutlich von jenen der Organismen ab, aus denen sie sich zusammensetzen. Erst in der Symbiose bilden sich die typischen Wuchsformen der Flechten heraus und nur in Lebensgemeinschaft mit einem Photobionten bilden die Mykobionten die charakteristischen Flechtensäuren. Die Wissenschaft von den Flechten ist die Flechtenkunde oder Lichenologie.
Weltweit gibt es rund 25.000 Flechtenarten. In Mitteleuropa kommen davon etwa 2.000 vor. Der Anteil endemischer Arten ist bei Flechten viel niedriger als bei Blütenpflanzen. Flechten werden immer nach dem Pilz benannt, der die Flechte bildet, da es meist dieser ist, der ihr die Form und Struktur gibt. Während mehrere Photobionten in einer Flechte vorhanden sein können, findet man immer nur eine Pilzart. Flechten werden daher den Pilzen (Fungi) zugerechnet, unter denen sie als eigene Lebensform eine Sonderstellung einnehmen; sie sind also keine Pflanzen.
Aufbau und Wuchsform
Flechten existieren in einem breiten Spektrum an Farben, das von weiß über leuchtendes Gelb, verschiedene Brauntöne, kräftiges Orange, tiefrot, rosa, olivgrün, blaugrün und grau bis zu tiefschwarz reicht.
Der Pilz bildet fast immer den eigentlichen Vegetationskörper der Flechte, ein Geflecht aus Pilzfäden (Hyphen), das so genannte Lager; darin eingeschlossen befindet sich eine Population der Photobionten. Die meisten Flechten bestehen aus mehreren Schichten.
Nach der Wuchsform und der Auflagefläche des Lagers, auch Pilzthallus genannt, unterscheidet man zwischen:
Die Einteilung in Wuchsformen ist künstlich und entspricht nicht den stammesgeschichtlichen Verwandtschaftsverhältnissen.
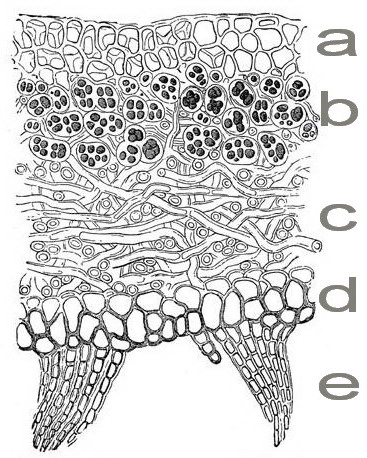
Bei den meisten Laubflechten wird auf der dem Substrat abgewandten Seite die äußere Schicht aus dichter geflochtenen Pilzfäden gebildet, sie wird obere Rinde genannt (a). Darunter liegt die Algenschicht, in der die Algen in einem lockeren Pilzgeflecht lagern (b). Anschließend folgt die Markschicht, die aus lockerem Pilzgeflecht ohne Algen besteht (c). Es schließt sich die untere, dem Substrat zugewandte Rinde an (d), die durch Rhizinen (e), wurzelartige Pilzfäden, die dem Substrat eng anliegen oder es durchdringen, verankert ist. Solche Flechtenkörper, in denen die Photobionten nur in einer Schicht liegen, nennt man heteromer. Wenn der phototrophe Partner dagegen mehr oder weniger regellos zerstreut im Pilzkörper liegt, spricht man von einem homöomeren Thallus.
Mykobiont, Photobiont und ihre Symbiose

Die Pilze gehören zu 98 Prozent der Abteilung der Schlauchpilze an, nur sehr wenige Arten sind Basidienpilze. Einige, nur steril bekannte Flechtenpilze werden formal den Deuteromycota oder Fungi imperfecti zugerechnet. Über 20 Prozent der heute bekannten Pilze leben in einer Flechtensymbiose.
In 85 Prozent der Fälle ist der Photobiont eine ein- oder wenigzellige Grünalge; bisher sind über 80 Arten aus etwa 30 Gattungen bekannt, die bedeutendste davon ist zweifellos Trebouxia.
Es gibt auch Flechten, bei denen der Partner aus dem Bakterien-Phylum der Cyanobacteria stammt. Dessen einzige Klasse Cyanobacteria enthält über 2.000 Arten in fünf Ordnungen. Mit Ausnahme der Ordnung Oscillatoriales weisen alle auch Vertreter in Flechtensymbiosen auf. Die wichtigste Cyanobakterien-Gattung mit Flechten-Symbionten ist Nostoc.
Manchmal kommen Grünalgen und Cyanobakterien auch zusammen in einer Flechte vor. Während alle Photobionten ohne ihren Pilzpartner leben können, findet man in der Natur die Mykobionten nicht ohne ihre domestizierten Partner; in Kultur können die meisten aber auch ohne Photobionten gehalten werden.
Die Symbiose zwischen Pilzen und Photobionten kann in unterschiedlichen Kontaktformen vorkommen. Die Pilzfäden können nur lose neben dem Partner liegen, man spricht dann von Kontakthyphen, sie können sie fest umschließen (Klammerhyphen) oder sogar in sie eindringen (Haustorium).
Die Vorteile der Symbiose liegen stark auf der Seite des Mykobionten und man beschreibt die Lebensgemeinschaft wahrscheinlich am besten als kontrollierten Parasitismus. Dies zeigt sich auch daran, dass der Pilz das Wachstum und die Zellteilungsrate der Alge kontrolliert. Aufgrund der langen Entwicklungszeit dieser probiotischen Beziehung hat sich daher ein Gleichgewicht zwischen Pilz und Alge eingestellt. Der Vorteil besteht für den Pilz darin, dass er von den Photobionten mit Nährstoffen versorgt wird, welche die Alge durch Photosynthese bildet. Der Pilz wiederum schützt den Partner vor zu rascher Austrocknung, da im Hyphengeflecht die Feuchtigkeit weniger stark schwankt; daneben schirmt er seinen Photobionten vor der Ultraviolettstrahlung ab. An Standorten, wo die Algen auf dem Boden pH-Werten zwischen 3,5 und 6,5 ausgesetzt wären, hilft das Leben im Verband mit dem Mykobionten bei der Aufnahme von Phosphat. Auch durch die gemeinsame Vermehrungsstrategie von Pilz und Alge ergibt sich für beide Symbionten ein Vorteil.
Von Grünalgen werden Zuckeralkohole, etwa Ribit, Erythrit oder Sorbit gebildet, die für den Pilz bekömmlicher als Kohlenhydrate sind. Bei Cyanobakterien als Partner wird hingegen Glucose transportiert. Bei den stickstofffixierenden Cyanobakterien wird auch reduzierter Stickstoff an den Mykobionten geliefert. Stoffströme des Primärstoffwechsels vom Pilz zum Photobionten sind nicht bekannt.
Wasserhaushalt

Flechten besitzen keine Möglichkeit, ihren Wasserhaushalt zu regeln, da sie keine echten Wurzeln zur aktiven Wasseraufnahme und auch keinen Verdunstungsschutz besitzen. Nur über die Oberfläche des Flechtenlagers können sie wie ein Schwamm Wasser in relativ kurzer Zeit aufsaugen, entweder in flüssiger Form oder als Wasserdampf. Bei Trockenheit verlieren sie relativ schnell das für die Aufrechterhaltung des Stoffwechsels nötige Wasser und wechseln in einen photosynthetisch inaktiven „leblosen“ Zustand, in dem der Wassergehalt bei weniger als zehn Prozent des Trockengewichts liegen kann. Es gibt starke Hinweise darauf, dass wie bei den mit ähnlichen Problemen konfrontierten Bärtierchen der Zucker Trehalose eine große Rolle beim Schutz von lebenswichtigen Makromolekülen wie Enzymen, Membranbestandteilen oder der Erbsubstanz DNA selbst spielt.
Anders als lange Zeit angenommen, schützt der Mykobiont den Photobionten nicht vor Austrocknung, sondern verlängert allenfalls die Zeit, die für diesen Prozess zur Verfügung steht. Der nahezu vollständige Feuchtigkeitsverlust ist vielmehr Teil der Überlebensstrategie von Flechten: Nur im ausgetrockneten Zustand sind sie in der Lage, Temperaturextreme oder hohe Lichtintensitäten, insbesondere von ultravioletter Strahlung zu überstehen; künstlich befeuchtete Flechten verlieren unter diesen Umständen dagegen schnell ihre Vitalität. Bei vielen Arten geht mit der Austrocknung eine Verdickung der Rindenschicht einher, die dadurch lichtundurchlässiger wird.
Die Fähigkeit der Ruhestarre ist besonders in kalten Gebieten sehr wichtig, da gefrorenes Wasser nicht für den Stoffwechsel verfügbar ist. Die Zeit, in der eine Flechte in einem solchen Stadium überleben kann, variiert je nach Art; es ist jedoch der Fall einer Wüstenflechte bekannt, die nach 40 Jahren im ausgetrockneten Zustand durch Befeuchtung „wiederbelebt“ werden konnte.
Erst bei erneuter Wasseraufnahme, über Regen, Tau oder Luftfeuchtigkeit, wird der Stoffwechsel reaktiviert. Bei einem Wassergehalt von 65 bis 90 Prozent des maximalen Speichervermögens erreicht er seine höchste Effizienz.
Weil die Luftfeuchtigkeit im Laufe eines Tages starken Schwankungen unterworfen ist, variiert entsprechend auch die Photosynthese-Rate der Flechten; meist ist sie am frühen Morgen, wenn das Flechtenlager von Tau benetzt wird, am höchsten.
Der vorstehend beschriebene Lebensrhythmus ist auch eine Ursache für das extrem langsame Wachstum mancher Flechten. Krustenflechten wachsen manchmal nur wenige Zehntel Millimeter pro Jahr, Laubflechten meist weniger als einen Zentimeter. Zum langsamen Wachstum trägt jedoch auch die ungleiche Symbiose bei, in welcher der Photobiont, der oft nur zehn Prozent des Flechtenvolumens einnimmt, allein für die Ernährung des Mykobionten aufkommen muss.
Das üppigste Wachstum findet man dagegen vor allem in subtropischen Nebelwäldern und nahe von Meeresküsten, wo eine nur geringen Schwankungen unterworfene Luftfeuchtigkeit für optimale Wachtumsbedingungen sorgt.
Flechtenstoffe
Die primären (intrazellulären) Produkte wie Proteine, Aminosäuren, Polysaccharide, Lipide, Vitamine etc. werden sowohl vom Photo- als auch vom Mykobionten gebildet und sind nicht flechtenspezifisch. Die so genannten Flechtenstoffe sind sekundäre (extrazelluläre) Produkte des Stoffwechsels und werden ausschließlich vom Pilz gebildet und auf den Hyphen deponiert.
Heute sind über 600 Stoffe bekannt, wobei die Hauptgruppen nach ihrer biosynthetischen Herkunft in die Acetyl-Polymalonate (etwa Usninsäure), die Shikimisäuren und die Mevalonsäuren eingeteilt werden. Dies sind auch die wichtigsten Farbpigmente wie etwa die gelbe Vulpinsäure oder das gelb-orange Parietin. Um Flechtensäuren nachzuweisen, nutzt man chemische Reagenzien, die eine Farbreaktion auslösen. Die wichtigsten sind Calcium- oder Natriumhypochlorit („C“), Kaliumhydroxid („K“) und p-Phenylendiamin („P“ oder „Pd“)[2].
Flechtensäuren spielen auch bei der Verwitterung eine wichtige Rolle, da sie Gesteine angreifen und so zur Bodenbildung beitragen. So musste im Sommer 2005 der Mount Rushmore von Flechten gereinigt werden.
Verbreitung und Lebensraum

Viele Flechten wachsen nur sehr langsam, meist nur wenige Millimeter im Jahr, einzelne Arten sogar nur Bruchteile eines Millimeters. Daher können sie nur an Standorten überleben, an denen sie nicht von anderen Pflanzen überwuchert und an der Photosynthese gehindert werden. An feuchten Standorten können sie sich oft nicht gegen Moose durchsetzen. Unter geeigneten Bedingungen, etwa dauerhafter Feuchte und geeigneten Temperaturen, wie im Regenwald oder Nebelwald, wachsen Flechten um einige Zentimeter im Jahr.
Flechten haben meist bescheidene Stoffwechselansprüche und begnügen sich mit geringen Mengen an Mineralstoffen aus Staub, der über die Luft angeweht wird, oder Nährstoffen, die im Regenwasser enthalten sind beziehungsweise aus dem Untergrund gelöst werden.
Viele Arten sind in der Lage, extreme Lebensräume zu erschließen. So können manche Flechten auf blankem Fels wachsen, andere wurden in fast 5.000 Meter Höhe im Himalaja-Gebirge gefunden. Sie kommen in der Wüste ebenso wie in Heidelandschaften, in Mooren ebenso wie in Permafrostgebieten vor und können in Trockenstarre Temperaturen von -47 Grad Celsius bis +80 Grad Celsius überstehen. In der Antarktis lassen sich etwa 200 Flechtenarten antreffen; selbst bei 86 Grad südlicher Breite findet man in den Horlick Mountains noch sechs Flechtenarten. Auch amphibische Arten, die permanent im Wasser leben, gibt es, etwa Verrucaria serpuloides.
Flechten besiedeln unterschiedlichste Standorte wie Baumrinde, Gesteine, Böden und selbst verrostetes Metall. Viele Flechtenarten sind substratspezifisch, das heißt, sie gedeihen nur auf basischem Gestein wie Kalkstein oder Dolomit oder „saurem“ kalkfreiem Silikatgestein wie Quarz, Gneis oder Basalt.
Flechten, die als Epiphyt auf Bäumen wachsen, sind keine Parasiten; sie entnehmen der Pflanze keine Nährstoffe oder Wasser, lediglich die Photosynthese wird durch die Abdeckung etwas behindert. Sie zeigen eindeutige Vorlieben für bestimmte Bedingungen wie saure Rinden von Fichten, Birken oder Erlen oder basenreiche Rinden von Nussbaum, Spitzahorn oder Holunder. Diese Merkmale sind oft wertvolle Bestimmungshilfen. Eine Reihe von Flechten dient selbst als Substrat für andere Flechten. Oft bilden sich typische Abfolgen, in denen verschiedene Flechtenarten in einer charakteristischen Reihenfolge übereinander geschichtet vorliegen.

Auf Fels sind Flechten wichtige Pionierorganismen, die entweder dem Gestein aufsitzen oder sogar in den Stein eindringen. Bei endolithischen Flechten ist das Lager im Inneren des Gesteins entwickelt, und äußerlich nur an einer Verfärbung des Gesteins erkennbar. Bei Vertretern der Gattung Verrucaria auf Kalkstein sind etwa nur die Perithecien genannten Fruchtkörper als schwarze Vertiefungen sichtbar. Nach dem Absterben des Lagers ist der Fels von kleinen Gruben übersät. Erst nach dem Anritzen des Steins erscheint die grüne Algenschicht. Trotz der Unauffälligkeit spielen diese Arten eine bedeutende Rolle bei der chemisch-physikalischen Verwitterung und Bodenbildung, umso mehr, da sie die Felsen oft flächendeckend überziehen.
Da Flechten naturgemäß keinen Unterschied zwischen Substraten in natürlicher und „künstlicher“ Umgebung machen, finden sie sich schließlich auch oft auf Mauern, Dächern, Zäunen oder Grabsteinen. Letztere können zur Datierung des Flechtenwachstums eingesetzt werden.
Der extremste „Lebensraum“, in dem Flechten bisher ihre Überlebensfähigkeit unter Beweis stellen konnten, ist ohne Zweifel der Weltraum. Durch im Mai 2005 durchgeführte Experimente an den Flechten Landkartenflechte (Rhizocarpon geographicum) und der Zierlichen Gelbflechte (Xanthoria elegans) konnte gezeigt werden, dass diese Arten zumindest für einen Zeitraum von etwa zwei Wochen in der Lage sind, die lebensfeindlichen Bedingungen außerhalb der Erdatmosphäre wie starke Temperaturschwankungen und hohe UV-Strahlungsintensität zu überstehen.
Vergesellschaftung

Wie bei Blütenpflanzen treten auch Flechtenarten miteinander bzw. mit Pflanzenarten vergesellschaftet auf. Die Benennung solcher Gesellschaften folgt Regeln, die in einem Code der pflanzensoziologischen Nomenklatur festgehalten sind[3].
Ein Beispiel einer von Flechten mitbestimmten Assoziation ist das Cladonio-Pinetum - der Flechten-Kiefernwald. Es handelt sich um die forstwirtschaftlich zwar uninteressanteste, naturschutzfachlich aber besonders wertvolle und seltene Ausprägung von Kiefernwald nährstoffärmster Standorte. Durch diffuse flächendeckende Einträge von Schad- und Nährstoffen (Eutrophierung) ist diese Gesellschaft in Mitteleuropa stark bedroht. Bei der durch die Nährstoffe begünstigten Sukzession wird sie vor allem durch Drahtschmielen-Kiefernwald verdrängt.
Fortpflanzung
Die Photobionten vermehren sich, solange sie in Flechtengemeinschaft leben, nur vegetativ, bilden also keine Gameten. Der Pilzpartner kann sich hingegen wie andere Pilze auch sexuell fortpflanzen.
Alle Sporen sind nur wenige tausendstel Millimeter groß. Die geschlechtlichen Sporen werden je nach der Zugehörigkeit des Mykobionten zu den Schlauch- oder Basidienpilzen in so genannten „Schläuchen“ (Asci) oder an so genannten „Ständern“ (Basidien) gebildet und dementsprechend als Asco- oder Basidiosporen bezeichnet.

Bei den Schlauchpilz-Flechten werden die Ascosporen in Fruchtkörpern gebildet, die sich nach ihrem Aufbau in zwei größere Gruppen einteilen lassen, Apothecien und Perithecien:
Der Pilz kann sich aber auch asexuell durch Pyknosporen fortpflanzen, die in Pyknidien gebildet werden. Dies sind kugelige bis birnenförmige Behälter, die in das Lager eingebettet sind. In diesen werden von speziellen Hyphen Pyknosporen abgegliedert. Pyknidien sind meist als sehr kleine schwärzliche Punkte auf dem Lager erkennbar.
Die Sporen verbreiten sich durch die Luft und können, wenn sie höhere Luftschichten erreichen, über weite Strecken, mitunter auch weltweit, verfrachtet werden. So erfolgt beispielsweise auch die Besiedelung isolierter Denkmäler oder Grabsteine, auch wenn das nächste Flechtenvorkommen weit entfernt ist.
Auf welche Weise sich die Flechtensynthese vollzieht, also wie die Gemeinschaft aus Myko- und Photobiont entsteht, ist noch nicht vollständig aufgeklärt. Der Pilzpartner muss zunächst einen geeigneten frei lebenden Algen- oder Bakterienpartner aufspüren und dann über diesen die Kontrolle übernehmen. Beides geschieht anscheinend erst, wenn sowohl Pilz als auch Alge oder Bakterium in einem „ausgehungerten“, dringend auf Nährstoffe angewiesenen Zustand sind. Auch im Labor ist es nur dann möglich, aus den beiden Einzelorganismen die Flechte zu bilden. Die typische Wuchsform der jeweiligen Flechtenart entsteht erst, nachdem der Pilzpartner seine Dominanz über den Photobionten etabliert hat.
Viele Flechten sind auf das Zusammentreffen solch günstiger Umstände nicht angewiesen und haben spezielle vegetative Vermehrungsorgane ausgebildet, mit denen Pilz und Alge gleichzeitig verbreitet werden können:
Alter

Flechten zählen zu den langlebigsten Lebewesen überhaupt und können ein Alter von mehreren hundert Jahren, in Einzelfällen sogar von über 4.500 Jahren erreichen, wie etwa bei einer Landkartenflechte (Rhizocarpon geographicum) aus Grönland. Durch ihre, nach einer Initialzeit, konstante Wachstumsrate können sie zur Altersbestimmung von bloßgelegten Steinen (Gletscherrückgang oder neu errichtete Bauwerke) genutzt werden. Die meisten Untersuchungen beziehen sich auf die gelben Sippen der Gattung Rhizocarpon, wobei der Durchmesser der Flechtenlager direkt zum Alter des Untergrunds in Bezug gesetzt wird. Diese Altersdatierung anhand von Flechten wird auch als Lichenometrie bezeichnet und wurde 1957 vom österreichischen Botaniker Roland Beschel eingeführt. 1965 bestimmte Gerhard Follmann etwa das Alter der durchschnittlich knapp 500 Jahre alten Monumentalfiguren auf der Osterinsel anhand des Flechtenbewuchses. Die Methode ist jedoch wegen des nicht immer gleichmäßigen Wachstums nicht unumstritten und wird nur dort genutzt, wo etwa die Radiocarbonmethode nicht angewandt werden kann[6].
Flechten und Tiere

Besonders im hohen Norden, wo die Vegetation spärlich ist, sind Flechten während der Wintermonate für Rentiere mit etwa 90 Prozent Hauptbestandteil der Nahrung. Meist handelt es sich um Rentierflechten (Cladonia), die sie mit ihren Hufen auch unter einer Schneedecke freilegen und mit Hilfe des Enzyms Lichenase verwerten können. Auch Elche nutzen diese Nahrungsquelle.
Für viele Larven von Schmetterlingen dienen Flechten als Nahrungsgrundlage, wie etwa für Vertreter der Gattung der Flechtenbärchen (Eilema), deren Raupen sich ausschließlich von Flechten ernähren.
Im Übrigen sind es vor allem wirbellose Tiere wie Schnecken, Insekten und Milben, zu deren Ernährung Flechten in unterschiedlichem Ausmaß beitragen. Dazu zählen auch Staubläuse (Psocoptera), manchmal auch Flechtlinge genannt, zu denen etwa die Bücherlaus (Liposcelis simulans) gehört. Erwähnenswert ist auch die Larve von Mycobates parmeliae, die sich mit ihrer leuchtend orangen Färbung an ihren Lebensraum in der Gewöhnlichen Gelbflechte angepasst hat.
Die Flechtenvegetation bietet vielen Tieren nebst Nahrung auch Lebensraum und Tarnung vor Fressfeinden. Milben und Insekten leben in großer Zahl zwischen Flechtenlagern; auch für die ebenfalls austrocknungsresistenten Bärtierchen sind Flechten ein wichtiger Lebensraum. Die Raupen verschiedener Nachtfalter tarnen sich mit Flechtenstückchen, andere ahmen einen flechtenbewachsenen Zweig nach (Mimikry).
Viele Vögel verwenden Flechten, vor allem blatt- und strauchförmige Arten, für den Nestbau, wie etwa der Wanderregenpfeifer (Pluvialis dominica), der sein Bodennest aus etwa 250 Thalli der Totengebeinsflechte und anderen Vertretern der Gattung Cladonia und Cetraria baut.
Bioindikatoren

Flechten gelten als Zeigerorganismen für bestimmte Umweltbedingungen, insbesondere die Luftqualität. Dies liegt daran, dass das Zusammenleben zwischen Pilz und Alge leicht gestört werden kann. Die in Luft und Regen enthaltenen Nähr- und Schadstoffe werden nahezu ungefiltert aufgenommen, da Flechten keine speziellen Organe zur Wasseraufnahme aus dem Boden besitzen und über den gesamten Thallus Feuchtigkeit aufnehmen. Daher reagieren sie besonders empfindlich auf Luftverschmutzung. Die ersten Berichte über eine massive Verarmung der Flechtenvegetation im Bereich industrialisierter Städte stammen aus der zweiten Hälfte des 19. Jahrhunderts, lange bevor Waldsterben und saurer Regen ins Blickfeld der Öffentlichkeit gerieten. Als Hauptursache konnte der erhöhte Schwefeldioxid-Gehalt der Luft identifiziert werden. Inzwischen haben Schwefelfilter in Industrieanlagen und Katalysatoren in Kraftfahrzeugen dazu beigetragen, die Luftgüte zu verbessern, so dass heute Flechten wieder häufiger in Großstädten aufzufinden sind.
Mit dem „Passiven Monitoring“ werden anhand von Verbreitung und Häufigkeit Rückschlüsse über die Luftgüte getroffen. Beim „Aktiven Monitoring“ werden mehrere Thalli einer bestimmten Art, meist der Blasenflechte, an einem belasteten Standort ausgesetzt und die Reaktionen wie Vitalitätsverlust, Verfärbung des Thallus oder gar das Absterben der Organismen beobachtet. Die Bioindikation mit Flechten ist allerdings auf lange Zeiten ausgelegt. In Deutschland sind passives und aktives Monitoring durch die VDI-Richtlinie 3799 standardisiert.
In Gebieten mit intensiver Landwirtschaft reagieren Düngemittel, die Stickstoffverbindungen enthalten, mit dem Regen schwach basisch. Dies führt vor allem zum Verschwinden der Flechtenarten, die saure Standorte bevorzugen.
Daneben sind Flechten Akkumulationsindikatoren für Schwermetalle, da sie die toxischen Partikel im Gewebe anreichern, was schließlich auch zum Absterben der Flechte führen kann.
Schließlich speichern Flechten auch radioaktive Substanzen. So lassen sie sich insbesondere zur Überwachung des radioaktiven Niederschlags nach atmosphärischen Kernwaffentests heranziehen. Nach dem Reaktorunglück von Tschernobyl gelangten große Mengen radioaktiver Isotope nach Finnland und wurden dort von Rentierflechten (Cladonia) aufgenommen. In Rentieren, die sich hauptsächlich von diesen Flechten ernähren, reicherten sie sich weiter an und gelangten schließlich über die Milch der Tiere und den daraus hergestellten Käse in die menschliche Nahrung.
Entwicklungsgeschichte
Älteste paläontologische Hinweise auf eine Symbiose aus Pilz und Alge belegen Fossilien aus Südchina, die rund 600 Millionen Jahre alt sind und somit aus der erdgeschichtlichen Epoche des Ediacarium stammen. Sie enthalten noch im Wasser lebende Flechten. Bis dahin galten Fossilien aus dem frühen Devon vor etwa 400 Millionen Jahren[7] als die ältesten Flechtenfossilien. Neuerdings wird Prototaxites aus dem Devon als mögliche "Riesenflechte" diskutiert. Ob diese an Land lebende Art von der in China gefunden Spezies abstammt, ist nicht geklärt, da Flechten mehrfach unabhängig entstanden sind. Taxonomisch gesehen handelt es sich bei Flechten um eine so genannte polyphyletische Gruppe der Pilze, das heißt die einzelnen Arten gehen nicht auf eine Flechten-Stammart zurück. Für ein hohes phylogenetisches Alter sprechen auch Überlegungen, dass diese Organisationsform vor den Gefäßpflanzen das Land besiedelte, da nur genügsame, wechselfeuchte Organismen erste Schritte auf blankem Fels unternehmen konnten.
Flechten und der Mensch
Geschichte der Flechtenkunde

Der griechische Botaniker Theophrastos, ein Schüler von Aristoteles, beschreibt erstmals in seinem Werk „Geschichte der Pflanzen“ zwei Flechtenarten, eine Bartflechte (Usnea) und eine Flechte auf Küstenfelsen (Rocella). Zu dieser Zeit erkannte man sie noch nicht als eigenständige Organismen, sondern hielt sie für Auswüchse von Bäumen oder Algen (Seetang).
Erst im 17. Jahrhundert entwickelte sich erneut Interesse und der Name „Lichen“ wurde nun gebräuchlich. Dieser leitet sich vom griechischen λειχην (leichän, lat. Lichen) ab und bedeutet „Warze“, was auf die Gestalt der Fruchtkörper hinweist. Die Anzahl der bis dahin bekannten Arten erhöhte sich auf nur 28. Der französische Arzt und Botaniker Joseph Pitton de Tournefort gliederte in einem neuen System die Flechten als eigene Abteilung „Lichen“ von den Moosen ab. Obwohl 1753 schon über 170 Arten bekannt waren, beschrieb Carl von Linné nur 80 Arten und bezeichnete sie als „armseligstes Bauernvolk“ der Vegetation.
Mit der Publikation der Schrift Methodus, qua omnes detectos lichenes ad genera redigere tentavit begründete Erik Acharius 1803 die wissenschaftliche Lichenologie. Er erstellte ein System, das auf dem Bau der Fruchtkörper beruht und verfasste eine Zusammenstellung aller zu seiner Zeit bekannten 906 Flechtenarten.
Der Arzt und Mykologe Heinrich Anton de Bary erkannte 1866 erstmals die Symbiose bei einer bestimmten Art von Gallertflechte. Die Vermutung, dass Flechten Doppelorganismen von Alge und Pilz sind, wurde 1869 vom Schweizer Botaniker Simon Schwendener aufgegriffen, der sie auf die übrigen Flechtenarten anwendete[8].
Heute hat sich die Lichenologie zu einer eigenen Disziplin entwickelt, die zwischen Mykologie und Botanik angesiedelt ist.
Verwendung

Die älteste Verwendung von Flechten ist jene als Nahrungsmittel. Oft wird diskutiert, ob es sich beim biblischen Manna um die Wüstenflechte Sphaerothallia esculenta gehandelt haben könnte.
Bestimmte Flechten, z.B. Cetraria islandica und Lecanora esculenta, wurden vor allem in Notzeiten gekocht oder als Mehlzusatz verwendet. Manche Teilnehmer schwieriger Expeditionen, etwa bei John Franklins Suche nach der Nordwest-Passage, haben nur dank Flechten überlebt. In Kanada waren manche Flechten als „tripes de roche“ (Felskutteln, rock tripe) bekannt. In der indischen Region Bellary wird aus einer Parmelia-Art das Currygericht „rathapu“ zubereitet. In Japan gilt die Nabelflechte Iwatake (Umbilicaria esculenta) als Delikatesse und findet als Suppe oder Salat Verwendung. In Nordamerika werden Bryoria-Arten als Nahrung zubereitet.
Seit dem Altertum werden Flechten auch als Heilmittel genutzt, etwa von dem griechischen Botaniker Theophrast. Die mittelalterliche Mystikerin Hildegard von Bingen schrieb:
Entsprechend der Signaturenlehre wurde früher die Echte Lungenflechte (Lobaria pulmonaria) gegen Lungenleiden eingesetzt und findet noch heute in der Homöopathie Verwendung. Im 17. und 18. Jahrhundert fand die Echte Lungenflechte, die auf Kiefern wuchs, in einem Kloster an der Ussolka in Sibirien als Bitterstoff (an Stelle des Hopfens) beim Bier brauen Verwendung. Mit der in Afrika endemischen Art Parmelia hottentotta wird Honigbier gewürzt.
Im Mittelalter wurden Flechten, die auf freiliegenden Totenschädeln wuchsen, als „Muscus cranii humani“ oder „Muscus ex cranio humano“ gegen Epilepsie verwendet.
Flechten enthalten oft eine große Vielfalt an Inhaltsstoffen, die sie für die pharmazeutische Industrie interessant machen. So wird das Isländische Moos (Cetraria islandica) Hustenmitteln beigegeben. Das Antibiotikum Usninsäure wurde im Baumbart (Usnea) entdeckt. Neuerdings sind gewisse Polysaccharide (sarcoma-180) in der Krebsbehandlung von Interesse.
Einige Arten werden zur Alkoholproduktion verwendet (z.B. Bryoria spp., Cladonia spp., Cetraria islandica).
Die durch Vulpinsäure giftige Wolfsflechte (Letharia vulpina) wurde früher zum Vergiften von Fuchs- und Wolfsködern genutzt.
Lange Zeit wurde aus den an Küstenfelsen vorkommenden Flechten der Gattung Roccella und der Art Pertusaria corallina die purpurfarbene Orseille, ein wertvoller Farbstoff, gewonnen. Lackmus ist ebenfalls ein Flechtenfarbstoff, der aus Roccella-Arten gewonnen wird. Auch andere Flechtenarten, etwa Evernia- oder Parmelia-Arten, können zum Färben von Wolle und Stoffen verwendet werden, was hauptsächlich in Skandinavien und Schottland praktiziert wurde. Vor allem angenehme Gelb- und Brauntöne können erzielt werden. Der schwedische Naturwissenschaftler Carl von Linné erwähnt in seinem Plantae tinctoriae sechs Färberflechten.
Das Baummoos (Pseudevernia furfuracea) und das Eichenmoos (Evernia prunastri) werden in der Parfümindustrie genutzt.
Die Alpen-Rentierflechte (Cladonia stellaris) wird schließlich in größeren Mengen aus Skandinavien importiert und findet als Modellbäumchen in Architekturmodellen oder in Kranzschmuck Verwendung.
Beim Wilde-Mändle-Tanz des Alpenraums, der heute noch alle fünf Jahre in Oberstdorf aufgeführt wird, sind die Darsteller am ganzen Körper mit langen, zottelig herabhängenden Bartflechten, die auf das leinene Gewand aufgenäht sind, geschmückt. Lediglich die Augenpartie bleibt frei. Sie tanzen zu urtümlicher, rhythmischer Musik.
Flechten in der Literatur

Ein mit Flechten behangener Baumriese war wohl die Vorlage für die „Ents“ genannten Baumwesen in J. R. R. Tolkiens Roman Der Herr der Ringe.
John Wyndhams Science-Fiction-Roman Ärger mit der Unsterblichkeit (Trouble with Lichen) erwähnt die Gewinnung des altershemmenden Wirkstoffs Antigeron aus Flechten.
Naturgemäß werden Flechten auch in Berichten über den Norden Europas an vielen Stellen erwähnt, etwa bei Alfred Andersch in seinen Reiseerzählungen „Hohe Breitengrade“ und „Nordische Wanderungen“.
Quellen und weiterführende Informationen
Einzelnachweise
Literatur
Weblinks
Wikimedia Foundation.