- Kleinhirnseitenstrangbahn
-
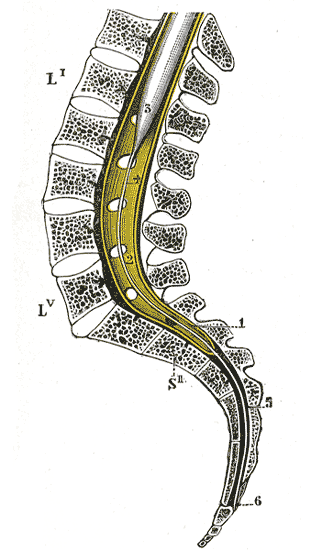
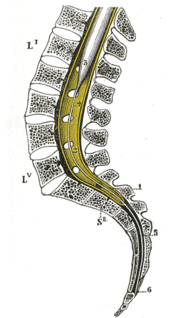 Lage des unteren Rückenmarks im Spinalkanal
Lage des unteren Rückenmarks im SpinalkanalDas Rückenmark (lat. Medulla spinalis, griech. μύελος – myelos) ist der Teil des zentralen Nervensystems (ZNS) der Wirbeltiere, der innerhalb des Wirbelkanals verläuft. Als Teil des ZNS ist das Rückenmark von denselben Häuten umgeben wie das Gehirn.
Inhaltsverzeichnis
Lage, Gestalt und Aufbau
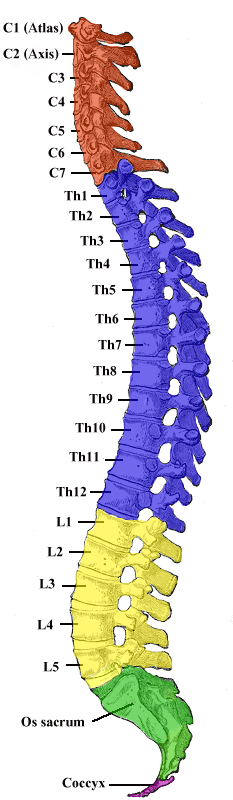
Das Rückenmark des Menschen reicht beim Ungeborenen vom Foramen magnum des Schädels noch bis zum Kreuzbein, beim Säugling bis zu den unteren Lendenwirbeln, beim Erwachsenen endet es auf Höhe des ersten bis zweiten Lendenwirbelkörpers. Dies erklärt sich dadurch, dass die Wirbelsäule im Laufe der menschlichen Entwicklung schneller wächst als das Rückenmark und daher das Längenwachstum des Rückenmarks hinter dem der Wirbelsäule zurückbleibt (sogenannter Ascensus medullae spinalis, „Aufstieg des Rückenmarks“).
Durch dieses Phänomen müssen die Wurzeln der Spinalnerven, da sie trotz des Ascensus dennoch aus „ihrem“ ursprünglichen Zwischenwirbelloch austreten, einen nach kaudal hin immer länger werdenden Weg innerhalb des Wirbelkanals zurücklegen, bevor sie ihn verlassen können. Ab dem 1. Lendenwirbelkörper, nach Ende des Rückenmarks, verlaufen nur noch die Wurzeln der Spinalnerven im Wirbelkanal abwärts. Sie bilden die Cauda equina (von lateinisch cauda „Schwanz“, „Schweif“ und equus „Pferd“; „Pferdeschweif“). Das Rückenmark ist wie das Gehirn von Gehirn-Rückenmarks-Flüssigkeit (Liquor cerebrospinalis) umgeben. Bei der Lumbalpunktion können durch Einstich auf Höhe des 3./4. Lendenwirbelkörpers Liquor entnommen und/oder Medikamente injiziert werden, ohne das Rückenmark zu verletzen.
Der Ascensus tritt bei den anderen Säugetieren ebenfalls auf, hier endet das Rückenmark jedoch auf Höhe der hinteren Lendenwirbel. Bei den anderen Wirbeltieren tritt dieses Phänomen nicht auf.
Hüllen
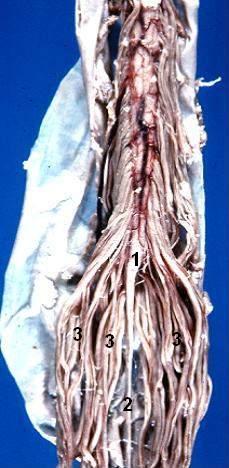
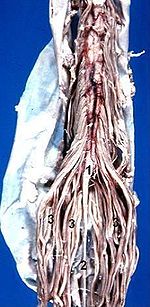 Eröffnete Rückenmarkshäute mit dadurch frei werdendem Blick auf kaudale Anteile des Rückenmarks: (1) Conus medullaris, (2) Filum terminale, (3) Cauda equina
Eröffnete Rückenmarkshäute mit dadurch frei werdendem Blick auf kaudale Anteile des Rückenmarks: (1) Conus medullaris, (2) Filum terminale, (3) Cauda equinaDie Rückenmarkshäute (Meninges medullae spinalis) verhalten sich prinzipiell wie die Hirnhäute. Auch hier unterscheidet man die äußere Dura mater, die mittlere Arachnoidea und die direkt dem Rückenmark anliegende Pia mater. Im Vergleich zur häutigen Umhüllung im Bereich des Gehirns gibt es zwei bauliche Besonderheiten. Zum einen ist die Dura mater nicht mit dem Wirbelkanal verwachsen, so dass zwischen Knochen und Dura mater ein mit Binde- und Fettgewebe ausgefüllter Raum ausgebildet ist, der sogenannte Epiduralraum bzw. Periduralraum. In den Epiduralraum lassen sich Lokalanästhetika applizieren und damit die austretenden Nerven ausschalten (Periduralanästhesie). Eine zweite bauliche Besonderheit sind segmentale Brücken zwischen Pia und Dura mater, das sogenannte Ligamentum denticulatum („gezahntes Band“). Diese in Aufsicht dreieckigen Strukturen dienen der stabilisierenden Aufhängung des Rückenmarks im mit Liquor cerebrospinalis gefüllten Subarachnoidalraum (Raum zwischen Arachnoidea und Pia mater).
Äußere Erscheinung
Das Rückenmark bildet einen langen Strang, der an zwei Stellen deutlich verdickt ist: An der Intumescentia cervicalis und an der Intumescentia lumbosacralis (intumescentia (lat.) = Anschwellung). Die Verdickungen kommen dadurch zustande, dass in diesen Bereichen jene Spinalnerven das Rückenmark verlassen, die die Arme und Beine innervieren, was eine größere Anzahl von Nervenzellen erfordert als beispielsweise die Innervation des Rumpfes. Nach unten hin läuft das Rückenmark spitz als Markkegel (Conus medullaris) aus und setzt sich in den etwa etwa 1 Millimeter dicken Endfaden (Filum terminale) fort, der nur noch aus Glia besteht und beim Menschen an der Hinterfläche des 2. Steißwirbels befestigt ist.
Vorn (bei Tieren unten) verläuft von oben nach unten eine kleine Furche, die Fissura mediana anterior (Tiere: Fissura mediana ventralis). Auf der Rückenseite verläuft der Sulcus medianus posterior. Auf beiden Seiten lassen sich drei Stränge (lat. = funiculi) voneinander abgrenzen: Paramedian der Fissura mediana der Vorderstrang (Funiculus anterior), hinten der Hinterstrang (Funiculus posterior) und zwischen den beiden der Seitenstrang (Funiculus lateralis). Die Hinterwurzeln der Spinalnerven treten jeweils in der Furche (lat. = sulcus) zwischen Hinter- und Seitenstrang (Sulcus lateralis posterior) aus, die Vorderwurzeln zwischen dem Seiten- und dem Vorderstrang (Sulcus lateralis anterior). Die Vorder- und Hinterwurzeln der Spinalnerven treten kontinuierlich aus dem Rückenmark als Fila radicularia aus und vereinigen sich erst in Höhe der Zwischenwirbellöcher (Foramina intervertebralia) der Wirbel zu den Spinalnerven.
Bei Vögeln ist im Bereich der Intumescentia lumbosacralis zwischen die Hinterstränge beider Seiten der sogenannte Glykogenkörper (Corpus gelatinosum) eingelagert. Es handelt sich dabei um eine gallertige, eiförmige Bildung von glykogenreichen Gliazellen, deren funktionelle Bedeutung bislang nicht geklärt ist. Beim Haushuhn ist sie etwa 1 cm lang und 4 mm breit.
Gliederung
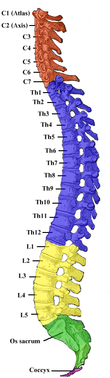
Das Rückenmark wird entsprechend den Austrittsstellen der Spinalnerven in fünf Abschnitte gegliedert, die jeweils in mehrere Segmente unterteilt werden können:
- Hals- oder Zervikalmark (Pars cervicalis): bei Säugetieren 8 Segmente (C1–C8)
- Brust- oder Thorakalmark (Pars thoracica): Segmente entsprechend der Wirbelzahl, beim Menschen 12 (Th1–Th12)
- Lenden- oder Lumbalmark (Pars lumbalis): Segmente entsprechend der Wirbelzahl, beim Menschen 5 (L1–L5)
- Kreuz- oder Sakralmark (Pars sacralis): Segmente entsprechend der Wirbelzahl, beim Menschen 5 (S1–S5)
- Schwanz- oder Kokzygealmark (Pars coccygis): beim Menschen rudimentär ausgebildet, 1 Segment (Co1)
Diese Gliederung richtet sich nicht nach der Lage der Abschnitte im Wirbelkanal, sondern lediglich nach den Austrittstellen der Spinalnerven aus der Wirbelsäule. Ein Segment entspricht dem Rückenmarksabschnitt, der die Fasern für je einen linken und rechten Spinalnerven liefert. Ab dem ersten Thorakalsegment wird der Spinalnerv mit seinem dazugehörigen Segment nach dem darüber liegenden Wirbel benannt, im Zervikalbereich dagegen nach dem darunter liegenden Wirbel, da der erste Spinalnerv nicht unter, sondern über dem ersten Halswirbel (Atlas) austritt. Das Halsmark hat also acht Segmente bei sieben Wirbeln, im Gegensatz zu den restlichen Abschnitten bei denen die Anzahl der Marksegmente mit der Anzahl der Wirbel übereinstimmt. Erklären lässt sich diese scheinbare Diskrepanz damit, dass Teile des Hinterhauptbeines (Os occipitale) entwicklungsgeschichtlich als Wirbel betrachtet werden können.
Feinbau
Das Rückenmark besteht aus grauer Substanz und weißer Substanz. Die graue Substanz besteht überwiegend aus Nervenzellkörpern, die weiße Substanz aus Axonen.
Graue Substanz
Die graue Substanz hat im Rückenmarksquerschnitt die Form eines Schmetterlings. Den vorderen, breiteren Flügelteil nennt man Vorderhorn (Cornu anterius, bei Tieren Cornu ventrale), den hinteren, schmaleren Teil Hinterhorn (Cornu posterius bzw. dorsale). Im Bereich des Thorakal- und Lumbalmarks befindet sich zwischen Vorder- und Hinterhorn das kleinere Seitenhorn (Cornu laterale). Die beiden Schmetterlingsflügel werden durch eine Querverbindung (Commissura grisea) miteinander verbunden. In der Mitte der Commissura grisea verläuft der Canalis centralis, der mit Liquor gefüllt ist und den inneren Liquorraum des Rückenmarks darstellt. Die graue Substanz kann in zehn Schichten (lateinisch laminae) eingeteilt werden. Die Laminae I–VII befinden sich im Hinterhorn, die Laminae VIII und IX im Vorderhorn. Die Lamina X bildet die Commissura grisea. Funktionell können einzelne Nervenkerne unterschieden werden (siehe Bild).
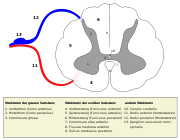 Querschnitt des Rückenmarks
Querschnitt des Rückenmarks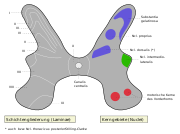 Laminae und Nuclei der grauen Substanz
Laminae und Nuclei der grauen SubstanzZelltypen
Neben den Wurzelzellen, die mit ihren Axonen die Spinalnerven bilden, gibt es in der grauen Substanz eine Vielzahl weiterer Zellen.
Die sogenannten Binnenzellen stellen den Eigenapparat des Rückenmarks dar. Dieser realisiert die Integrationsleistungen des Rückenmarks, zum Beispiel für die Generierung von Reflexen. Zum Eigenapparat gehören des Weiteren Schaltzellen, Assoziationszellen und Kommissurenzellen. Die Fortsätze der Schaltzellen (oder Interneurone) verlassen die graue Substanz nicht, sondern verlaufen ipsilateral (auf der gleichen Körperseite) innerhalb eines Rückenmarkssegments. Sie stellen die Verbindung zwischen Zellen des gleichen Segments her und vermitteln z. B. Eigenreflexe. Wichtige Interneurone sind auch die Renshaw-Zellen, die einen hemmenden Rückkopplungseinfluss auf Motoneurone haben. Die Assoziationszellen (propriospinale Neurone) verbinden verschiedene Rückenmarkssegmente miteinander. Ihre Fortsätze verlassen die graue Substanz ebenfalls nicht, auch sie verlaufen ipsilateral. Die Kommissurenzellen kreuzen mit ihren Axonen auf die andere Körperseite (kontralateral) innerhalb eines Rückenmarkssegments. Sie sorgen dafür, dass Erregungsimpulse auch die andere Rückenmarksseite erreichen.
Strangzellen (Projektionsneurone) gehören nur teilweise zum Eigenapparat des Rückenmarks. Sie verlassen mit ihren Neuriten die graue Substanz und ziehen in die weiße Substanz. Hier ziehen sie in Strängen und übermitteln Informationen an andere Rückenmarkssegmente und das Gehirn.
Assoziations-, Strang- und Kommissurenzellen sind an der Auslösung der Fremdreflexe beteiligt.
Hinterhorn
Das Hinterhorn erhält über die dorsale (sensible) Wurzel des Spinalnervs sensible Informationen aus der Peripherie. Diese Informationen werden in Richtung Gehirn weitergeleitet.
Die sensiblen Wurzelzellen sind pseudounipolare Nervenzellen, deren Zellkörper außerhalb des Rückenmarks im Spinalganglion liegen. Ihre Axone treten über die dorsale (sensible) Wurzel in das Hinterhorn des Rückenmarks. Hier werden sie entweder auf ein zweites Neuron umgeschaltet oder ziehen direkt über spezielle Bahnen der weißen Substanz in das Gehirn.
Vorderhorn
Das Vorderhorn enthält Motoneurone. Die Motoneurone entsenden ihre Axone, als ventrale Spinalnervenwurzel zusammengelagert, zu den Muskeln und Muskelspindeln. Sie sind also für Bewegung und Muskeltonus zuständig.
Seitenhorn
Im Bereich des Brust- und Lendenmarks befindet sich, zwischen dem Vorder- und dem Hinterhorn liegend, das sogenannte Seitenhorn, das von den Zellkörpern vegetativer Neurone gebildet wird und zum Sympathikus gehört. Ihre efferenten Fasern verlassen das Rückenmark über die Vorderwurzel, ziehen zum Grenzstrang und werden zum Teil dort auf das zweite Neuron verschaltet. Der Rest zieht zu den prävertebralen Ganglien, um dort verschaltet zu werden. Sympathisch-afferente Fasern gelangen aus der Peripherie mit der Hinterwurzel zum Seitenhorn. An äquivalenter Stelle liegen im Sakralmark Zellkörper parasympatischer Neurone, allerdings ohne ein erkennbares Seitenhorn zu bilden.
Weiße Substanz
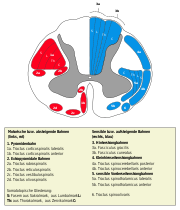 Aufsteigende und absteigende Bahnen der Weißen Substanz
Aufsteigende und absteigende Bahnen der Weißen SubstanzIn der weißen Substanz des Rückenmarks verlaufen aufsteigende (meist sensible) Bahnen, die zum Gehirn ziehen und absteigende (meist motorische) Bahnen, die vom Gehirn kommen. Ursache für die weiße Farbe ist die Myelinisierung der Axone.
Wichtige aufsteigende Bahnen
Die Hinterstrangbahn, bestehend aus Fasciculus cuneatus und Fasciculus gracilis, leitet ungekreuzt Informationen über die epikritische (= feine Berührungen) und die Tiefensensibilität in die Medulla oblongata. Es erfolgt keine Umschaltung im Hinterhorn, dafür aber in der Medulla oblongata in den entsprechenden Kernen (Nuclei), nämlich im Nucleus gracilis für die untere Körperhälfte und dem Nucleus cuneatus für die obere Körperhälfte. Kurz nach der Umschaltung auf das zweite Neuron findet die Kreuzung der Fasern auf die andere Seite statt. Die Zellkörper der in der Hinterstrangbahn verlaufenden Axone liegen in den Spinalganglien. Sie bilden das lemniskale System.
Der Tractus spinothalamicus (sensibles anterolaterales Bahnsystem oder Vorderseitenstrangbahn) leitet Impulse der protopathischen Sensibilität (grobe Druck-, Temperatur- und Schmerzempfindung) von den Zellen des Hinterhorns bis zum Thalamus; er bildet das Extralemniskale Bahnsystem. Die Fasern kreuzen auf Höhe des Ursprungssegmentes auf die andere Seite.
Der Tractus spinocerebellaris (Kleinhirnseitenstrangbahn) entspringt vor allem aus dem Nucleus dorsalis des Hinterhorns und endet im Kleinhirn. Er verläuft effektiv ungekreuzt und leitet propriozeptive (= Informationen über Lage und Stellung von Gelenken, Muskeln, Sehnen) Informationen.
Wichtige absteigende Bahnen
Die Pyramidenbahn (Tractus corticospinalis) reicht vom motorischen Cortex der Großhirnrinde bis ins Vorderhorn.
Als extrapyramidale Bahnen bezeichnet man alle motorischen Bahnen, die nicht zur Pyramidenbahn gehören. Zu ihnen gehören der Tractus rubrospinalis, der Tractus vestibulospinalis, der Tractus reticulospinalis und der Tractus tectospinalis. Sie alle enden ebenfalls im Vorderhorn.
Neurotransmitter im Rückenmark
Der wichtigste inhibitorische Neurotransmitter im Rückenmark ist Glycin. Interneurone und z. B. die Renshaw-Zellen sind glycinerg, d. h. sie setzen Glycin aus ihren synaptischen Endigungen frei.
Blutversorgung des Rückenmarks
Das Rückenmark wird durch radiär in die weiße und graue Substanz des Rückenmarks einstrahlende Äste von drei Arterien mit sauerstoffreichem Blut versorgt. Die Arteria spinalis anterior versorgt das Vorderhorn, den größten Anteil des Vorderseitenstrangs und die Basis des Hinterhorns. Der restliche Anteil des Rückenmarks wird durch die beiden Arteriae spinales posteriores versorgt. Das venöse sauerstoffarme Blut wird über zwei große Venen abtransportiert, die teilweise parallel zu den Arterien verlaufen: Vena spinalis anterior und Vena spinalis posterior.
Embryologie und Fehlbildungen
Das Rückenmark entsteht beim Wirbeltier-Embryo aus den Neuralfalten des Ektoderms, unmittelbar über der Chorda dorsalis. Die Neuralfalten verschließen sich zum Neuralrohr, einem mit Flüssigkeit gefüllten Kanal, der von einem Epithel umgeben ist. Bei Störungen dieses Verschlusses treten verschiedene Formen der Dysraphie auf. Liegt nur eine Störung im Lendenbereich vor, entsteht eine Spina bifida aperta („offener Rücken“), bei der die Wirbelbögen rückenseitig unvollständig bleiben. Dabei können sich nur die Rückenmarkshäute (Meningozele) oder zusätzlich auch das Rückenmark (Meningomyelozele) durch diesen Defekt nach außen vorwölben. Wenn sich die Neuralrinne an keiner Stelle schließt entsteht eine Myelozele (Rachischisis).
Aus dem Neuralepithel differenzieren sich die Nerven- und Gliazellen. Die Spinalganglien und sympathischen Neurone entstehen aus der sogenannten Neuralleiste. Um das Neuralrohr entstehen aus dem Mesenchym die Urwirbel, aus denen unter anderem die Wirbelsäule entsteht.
Zunächst verlassen Proneurone (die Vorläufer der Neurone) das Neuroepithel und bilden eine Mantelschicht (Mantelzone), aus der die graue Substanz entsteht. Die meisten Axone dieser Proneurone wachsen in Richtung Oberfläche und bilden so die Randschicht (Marginalzone), die später zur weißen Substanz wird.
Innerhalb der Mantelschicht entwickeln sich durch Zellteilungen und Zellmigration zwei Verdickungen, die Flügelplatte (Alarplatte) und die Grundplatte (Basalplatte), die nach und nach das spätere Erscheinungsbild eines Schmetterlings aufweisen. Die Randschicht wird durch das weitere Einwachsen auf- und absteigender Axone, deren Myelinisierung und der Vergrößerung der Flügel- und Grundplatten in 3 Stränge gegliedert: Hinterstrang, Seitenstrang und Vorderstrang.
Blutversorgung
Drei Arterien versorgen das Rückenmark: Vorn die Arteria spinalis anterior, hinten zwei Arteriae spinales posteriores. Bei Tieren mit horizontaler Körperstellung wird stattdessen von Arteria spinalis ventralis und dorsalis gesprochen.
Diese drei Arterien entspringen im Halsbereich beidseits aus den seitlich in der Wirbelsäule verlaufenden Arteria vertebralis. Im Thorax- und Lendenbereich erhalten sie segmentale Zuflüsse aus den Interkostalgefäßen (Arteriae intercostales dorsales) und den Lendengefäßen (Arteriae lumbales).
Im Bereich zwischen dem 4. und dem 6. Brustwirbel ist die Ausbildung von solchen Zuflüssen gering, sodass bei einem Gefäßverschluss (Rückenmarksinfarkt) dieser Abschnitt besonders gefährdet ist.
Phylogenese
In der hier beschriebenen Form ist das Rückenmark bei allen Vertretern der Schädeltiere (Gnathostomata) aufgebaut, wobei es in den unterschiedlichen Taxa jeweils geringe Unterschiede im Detail gibt.
Bei den basalen Chordatieren, zu denen die Schädellosen (Acrania) sowie die Manteltiere (Tunicata) gehören, gibt es kein differenziertes Rückenmark. Hier existiert stattdessen nur das Neuralrohr, welches dem späteren Rückenmark homolog ist. An seinem Vorderende besitzt es eine Verdickung, die als Hirnbläschen bezeichnet wird, mit dem Gehirn der Wirbeltiere allerdings nicht viel gemein hat. Das Neuralrohr selbst ist aus teilweise hoch differenzierten Neuronen aufgebaut, über Verschaltungen und Funktionen derselben ist allerdings wenig bekannt. Die Hauptaxone gehen von den so genannten Kollossalzellen aus, die große Zwischenneurone darstellen und deren Axone ventromediane Riesenfasern bilden. Segmental befinden sich dorsale Nervenwurzeln, die den späteren Spinalganglien entsprechen, aufgrund der versetzten Myomere allerdings versetzt zueinander sind. Während bei den Acraniern dieser Aufbau bis zum adulten Tier besteht, findet er sich bei den Tunicaten nur im Larvenzustand.
Bei den Schleimaalen (Myxinoida) und den Neunaugen (Petromyzontida), die als Kieferlose (Agnatha) die ursprünglichsten Taxa der Schädeltiere (Craniota) darstellen, existiert bereits ein Rückenmark, bei dem das Neuralrohr wie bei den späteren Wirbeltieren nur das Lumen darstellt. Bei den Schleimaalen ist es bandförmig aufgebaut und besitzt verschiedene Motoneurone und Interneurone ohne eine segmentale Anordnung, die Wurzeln der Spinalnerven vereinigen sich erst in der somatischen Muskulatur und eine Reihe von Organen werden direkt von Spinalnerven innerviert. Das Rückenmark der Neunaugen ähnelt dagegen bereits deutlicher dem der späteren Wirbeltiere. Es weist eine Trennung in weiße und graue Substanz auf und die Zellkörper der Riesenaxone liegen vornehmlich im Hirnstamm. Andererseits ist die Verschaltung der Neurone in der Wirbelsäule so komplex, dass die Tiere auch nach einer Durchtrennung derselben vor dem Hirn, etwa durch Entfernung des Kopfes, noch koordiniert schwimmen können. Die dorsalen und ventralen Wurzeln der Spinalnerven vereinigen sich bei den Tieren nahe dem Rückenmark, treten jedoch nicht segmental in gleicher Höhe aus dem Rückenmark hervor.
Die Knorpelfische (Chondrichthyes), zu denen die Haie und Rochen gehören, besitzen ein Rückenmark mit segmentalen Spinalnerven, wie es für die Schädeltiere typisch ist. Bei ihnen sind jedoch einige Besonderheiten vorhanden. Auch bei ihnen ist das Rückenmark sehr stark autonom und ein Hai kann auch dann noch koordiniert schwimmen, wenn das Rückenmark am Hals durchtrennt wird. Anders als bei allen anderen Wirbeltieren nimmt die Anzahl der sensorischen und motorischen Wurzelganglien während des Wachstums lebenslang zu. Neben weiteren Unterschieden im Feinbau, wie etwa einem separaten absteigenden Strang für die Kerne des Hirnnerven VIII besitzen die Knorpelfische ein kaudales neurosekretorisches System, welches eine wichtige Rolle bei der Osmoregulation sowie bei der Fortpflanzung spielt.
Bei allen folgenden Taxa, von den Strahlenflossern bis zu den Amphibien, den verschiedenen Reptilientaxa sowie den Vögeln und den Säugetieren entspricht das Rückenmark in Aufbau und Funktion dem dargestellten Grundmuster und unterscheidet sich nur in sehr speziellen Details voneinander.
Schädigungen des Rückenmarks
Schädigungen des Rückenmarks können durch
- Druck (Tumor, Bandscheibenvorfall, Wobbler-Syndrom, Cauda-equina-Syndrom),
- Durchtrennung (Spinalisation mit nachfolgender Querschnittlähmung),
- Entzündungen (Myelitis, Multiple Sklerose),
- Durchblutungsstörungen (Fibrokartilaginöse Embolie),
- degenerative Prozesse (Degenerative Myelopathien der Hunde, Cauda-equina-Syndrom der Hunde), Syringomyelie oder eine
- angeborene Missbildung wie die Chiari-Malformation Typ II bzw. III
ausgelöst werden.
Untersuchung des Rückenmarks
Das Rückenmark selbst ist der klinischen Untersuchung durch seine Lage im knöchernen Wirbelkanal nicht direkt zugänglich. Reflexuntersuchungen und die Verteilung von Lähmungen (Paralysen) oder Gefühlsstörungen geben dem Neurologen jedoch wichtige Hinweise auf die Art und Lokalisation der Schädigung. Neurophysiologische Untersuchungsmethoden wie motorisch und somatosensibel evozierte Potenziale erlauben eine funktionelle Überprüfung einzelner Bahnsysteme des Rückenmarks wie den Tractus corticospinalis und die Hinterstrangbahnen. Untersuchungen von Reflexbahnen über Motoneurone im Rückenmark sind möglich mit Hilfe des H-Reflexes und der F-Welle.
Strukturelle Veränderungen lassen sich dagegen mit den bildgebenden Verfahren vor allem der Myelografie, der Computertomografie und der Magnetresonanztomografie erkennen. Fehlbildungen im Blutgefäßsystem des Rückenmarks lassen sich mit der Angiografie darstellen. Hinweise auf Entzündungen kann auch die Lumbalpunktion und Untersuchung der Rückenmarksflüssigkeit liefern.
Weblinks
Literatur
- F.-V. Salomon, H. Geyer, U. Gille: Anatomie für die Tiermedizin. Enke-Verlag, Stuttgart 2004, ISBN 3-8304-1007-7
- Th. H. Schiebler (Hrsg.): Anatomie. Springer-Verlag, 9. Aufl., 2005, ISBN 3-540-21966-8
- W. Westheide, R. Rieger: Spezielle Zoologe; Teil 2: Wirbel- und Schädeltiere. Gustav Fischer Verlag, Stuttgart, Jena, 2004, ISBN 3-8274-0307-3



Wikimedia Foundation.