- Müller’sche Körperchen
-
Ameisenbäume 
Cecropia glaziovi
Systematik Abteilung: Bedecktsamer (Magnoliophyta) Klasse: Dreifurchenpollen-
Zweikeimblättrige (Rosopsida)Unterklasse: Rosenähnliche (Rosidae) Ordnung: Rosenartige (Rosales) Familie: Brennnesselgewächse (Urticaceae) Gattung: Ameisenbäume Wissenschaftlicher Name Cecropia Loefl. Ameisenbäume (Cecropia) sind eine 61 Arten umfassende Gattung zweihäusiger Bäume in der Familie der Brennnesselgewächse (Urticaceae). Die Mehrzahl ihrer Arten lebt als Ameisenpflanzen in einer Myrmekophylaxis genannten Symbiose mit Ameisen der Gattung Azteca. Ursprüngliches und hauptsächliches Verbreitungsgebiet der Gattung ist die Neotropis.
Inhaltsverzeichnis
Merkmale
Ameisenbäume sind ausdauernde Bäume. Die meisten Arten erreichen Wuchshöhen zwischen fünf und 15 Metern. Exemplare der Arten C. distachya, C. herthae, C. insignis und C. sciadophylla werden aber deutlich größer und erreichen Höhen von bis zu 40 Metern. Besonders niedrig bleibt zum Beispiel C. ulei, Pflanzen dieser Art werden selten höher als fünf Meter.
Ebenso variabel wie die Wuchshöhe ist auch das Alter, das die Ameisenbäume erreichen können. Können Exemplare von C. hololeuca mehr als einhundert Jahre alt werden, erreichen Bäume der Art C. glaziovii nur ein Alter zwischen 30 und 40 Jahren. Noch kürzer leben Exemplare von C. ulei, die nur wenige Jahre alt werden.
Wuchsformen
 Habitus von Cecropia pachystachya
Habitus von Cecropia pachystachyaAmeisenbäume sind für gewöhnlich nur wenig verzweigt mit einem leuchterähnlichen Astsystem. Bei einigen Arten ist die Verzweigung soweit reduziert, dass der Baum nur aus einer einzelnen Sprossachse besteht, Beispiele für diese Wuchsformen sind C. megastachya und C. ulei. Viele Arten, wie zum Beispiel C. concolor oder C. ficifolia wachsen bis zur ersten Blüte unverzweigt und verzweigen sich erstmals mit der Samenentwicklung, wobei die Äste aus den Achsen der kreuzgegenständig (dekussiert) stehenden Blattstiele auswachsen.
C. garciae und C. hispidissima sind stärker verzweigt. Hier beginnen die Äste auf eine Höhe zwischen einem halben und einem Meter. Die Pflanzen bekommen einen eher strauchähnlichen Habitus. Bei den meisten Arten stehen die Zweige in flachem Winkel zum Stamm (zumindest größer als 45°) und geben der Krone eine schirmartige Form. Einige Arten jedoch wie beispielsweise C. putumayonis oder C. uncubambana entwickeln Zweige in einem spitzeren Winkel, was in einer gedrungeneren Krone resultiert.
Wurzeln
Alle Arten von Ameisenbäumen bilden Adventivwurzeln aus, die sich zu Stelzwurzeln weiterentwickeln. Die Stelzwurzeln werden beim Wachstum nicht durch das Längenwachstum der Sprossachse aus dem Substrat herausgehoben, sondern wachsen überirdisch aus Lentizellen aus.
Die Wurzeln verankern sich nicht sehr tief im Substrat (dem Erdboden). Sogar bei ausgewachsenen Pflanzen reichen sie selten tiefer als drei Meter. Sie breiten sich radial aus und erreichen einen Durchmesser von etwa vier Metern. Einzelne − typischerweise bis zu drei – Wurzeln werden aber bis zu 15 Meter lang. Feine, Haufen bildende Wurzeln finden sich nur in den obersten 10 Zentimetern des Substrats.
Eine klare Unterscheidung der Wurzeln in die von Jeník und Sen vorgeschlagenen Klassen Makrorhizae (sich vielteilende dicke Wurzeln für Längen- und Tiefenwachstum) und Brachyrhizae (sich zweiteilende dünne Wurzeln)[1] ist bei den Ameisenbäumen nicht möglich.
Die Wurzeln von Cecropia können in die dicken Wurzeln benachbarter Bäume wie Miconia poeppigii und Arten der Gattung Clusia einwachsen.[2]
Sprossachse
Die Internodien der Sprossachsen sind bei fast allen Arten hohl und tragen nur am Rand eine dünne Schicht eines weißen, weichen Marks. Bei C. bullata und C. gabrielis hingegen sind sie vollständig mit einem härteren braunen Mark gefüllt, bei C. schreberiana oder C. telealba teilweise ausgefüllt. Allgemein kann gesagt werden, dass die Markschicht bei langsamer wachsenden, montanen Arten eher dicker als bei schnellwachsenden Spezies ist. Oft verdickt sich der Stamm bei jungen Pflanzen nach oben, dies kann graduell oder mehr oder weniger abrupt passieren.
Die Blattansatzstellen (Nodien) sind verdickt. Dort bilden sich im Inneren des Stängels Scheidewände (Septa), so dass die einzelnen Sprossteile zwischen den Nodien (Internodien) räumlich getrennt sind. Das Mark in den Internodien ist allseits von einer dünnen Haut eines extrem harten Sklerenchyms umgeben. Diese Sklerenchymschichten setzen sich horizontal an den Nodien fort und machen die Scheidewände sehr stabil.
Die meisten Arten bilden an den oberen Grenzen der Internodien kleine Vertiefungen, so genannte Prostomata aus. Diese sitzen oft genau mittig, oberhalb der Ansatzstelle eines Blattstiels. Die Wand des Internodiums ist dort besonders dünn. Schimper nannte die Prostomata in seiner Arbeit von 1888 auch Diaphragma.[3] Bei den myrmekophylaktischen Arten sind die Prostomata häufig von den Ameisen durchbissen und bilden ein kleines Loch in der Sprossachse.
Die Länge der Internodien variiert innerhalb einer Pflanze beträchtlich. Junge Bäume bilden in der Regel längere Internodien aus, bei adulten Exemplaren verkürzt sich die Länge der neu gebildeten Internodien auf 0,5 bis zwei Zentimeter. In den distalen Teilen der Äste sind die Internodien für gewöhnlich länger als zwei Zentimeter.
Bei vielen Arten zeigen die Sprossachsen deutliche Narben an den Stellen, wo alte Blätter abgefallen sind. Bei C. annulata, C. engleriana und C. litoralis kann an diesen ringförmig stehenden Narben ähnlich wie an Jahresringen das Alter des Baumes abgelesen werden.
Trichilium
Bei den meisten Ameisenbaum-Arten ist die Unterseite der Blattansätze mit einem oder zwei Haarpolstern (Trichilium) besetzt. Diese bestehen aus sehr verschiedenen Trichomen, die aber immer aus einer einzigen Zelle bestehen. Sie sind zumeist weiß oder bräunlich.
Auf dem Trichilium bilden sich perlenartige kleine Tröpfchen eines latexartigen Saftes, der reich an Proteinen und Fetten ist, die so genannten Müller’schen Körperchen. Ein einziges Trichilium bildet etwa 2500–8000 dieser Körperchen pro Woche, was etwa 10 Gramm entspricht.[4]
Einige Arten bilden gar keine Trichilia aus, dies sind zum Beispiel: C. holoeuca, C. pittieri, C. sciadophylla und C. tacuna, dabei handelt es sich vor allem um montane Arten, die keine Myrmekophylaxis kennen.
Blätter
 Blätter von Cecropia pachystachya
Blätter von Cecropia pachystachyaDie Blätter aller adulten Ameisenbaumarten sind auffällig groß und schildförmig, mit fast kreisrundem Umfang. Die Spreite sitzt exzentrisch, außerhalb des Mittelpunktes, am Blattstiel. Die Nervatur ist fingernervig, und die Spreite ist zwischen den Hauptnerven eingeschnitten, so dass sich handförmige Blätter in verschiedenen Abstufungen (handförmig gelappt, handförmig gespalten, handförmig geteilt, handförmig geschnitten) oder sogar gefingerte Blattformen ergeben. Bei einigen Arten, wie zum Beispiel C. sciadophylla reichen diese Einschnitte bis zum Blattstiel und die einzelnen Blattteile stehen an eigenen kleinen Stielchen. Bei anderen Arten, wie zum Beispiel C. putumayonis oder C. subintegra, ist der untere Teil der Spreite kaum oder gar nicht eingeschnitten und nur der obere Teil gespalten. Die Anzahl der Blattsegmente variiert zwischen den Arten von fünf bis über 20, innerhalb einer Art variiert die Zahl innerhalb engerer Grenzen.
Der Verlauf der Hauptnerven ist innerhalb der Arten der Gattung relativ uniform. Die Narvatur der Seitennerven hingegen variiert stark zwischen den Spezies. Die Epidermis (Blattoberfläche) ist ledrig bis papierartig. Ledrige Blätter sind häufig glatt, wohingegen papierartige eher rau sind.
Der Blattstiel ist etwa so lang wie die Spreite, bei einigen Arten, wie zum Beispiel C. marginalis oder C. virgusa, aber auch nur halb so lang. Blattstiele die länger als 40 Zentimeter werden, sind jedoch selten. Die Spreiten sind an der Basis oft gefaltet und ausgebreitet zur Spitze hin. Bei C. augustifolia und C. montana ist die Blattbasis oft um den Blattstiel umgerollt.
Blattentwicklung
Das Kotyledon (Keimblatt) der Ameisenbäume ist klein und knorpelig. Es ist chlorophylllos und hat keine Nervatur. Die ersten Trophophylle, Blätter die die Pflanze durch Photosynthese ernähren, sind gegenständig. Sie sind breitlanzettlich, genervt und haben einen kurzen Blattstiel. In der weiteren Entwicklung werden die Blattstiele länger und die Blätter zunächst an zwei Stellen eingeschnitten. Nach zweimal eingeschnittenen Blatt mit drei Blattsegmenten entwickeln sich Blätter mit fünf Segmenten, dann siebensegmentierte Blätter und so weiter. Auch die Blattgröße steigt von Generation zu Generation.
Nebenblätter
Die Nebenblätter der Ameisenbäume sind paarig rechts und links der Basis des Blattstiels angewachsen. Ihre Länge variiert von fünf bis zu 50 Zentimetern. Die Spitze ist oft umgebogen oder zu einer Knospe verwachsen.
Blütenstände
Normalerweise stehen in jeder Blattachsel zwei Blütenstände mit einer lateralen Knospe zwischen ihnen. Bei vielen Arten steht dem Blütenstand ein basales Tragblatt gegenüber. Die Tragblätter variieren in der Größe von Art zu Art, werden aber nicht länger als 2,5 Zentimeter.
Der Blütenstand besteht aus einem Blütenstiel, der mehrere Ähren trägt. Zunächst ist jede dieser Ähren komplett in eine Blütenscheide, die Spatha, gehüllt. Bei der Anthese öffnet sich die Spatha zunächst adaxial und fällt dann herunter. C. hololeuca ist die einzige Art, die gar keine Blütenscheiden ausbildet. Vor der Anthese ist die Spatha immer länger als die längste Ähre. Die Gestalt, Oberfläche und Farbe der Spatha ist immer ähnlich den Blütenstielen.
Die Anzahl der Ähren variiert zwischen den Arten. Bei weiblichen Blüten sind es in der Regel vier Ähren. bei C. gabrielis jedoch nur eine einzige und bei C. garciae und C. hispidissima bis zu 20. Bei männlichen Blüten sind es üblicherweise deutlich mehr Ähren, bei C. membranaceae sind es sogar bis zu 100. Die Blüten sind zumeist gelblich.
Männliche Blüten
Die männlichen Blüten stehen bei allen Spezies frei, nur bei C. purpurascens sind die oberen Teile der Blütenhüllen verwachsen. Die Länge des Perianths variiert abhängig von seiner Position innerhalb der Ähre, zur Basis und zur Spitze hin werden sie kürzer, seine Form ist röhrig. Die Tepalen sind fast vollständig verwachsen und lassen nur eine schmale schlitzförmige Spaltöffnung offen. Nur bei C. marginalis sind die Tepalen freiblättig. Der obere Teil der Blüte ist oft verdickt und hohl. Im Gegensatz zu den weiblichen Blüten sind die männlichen Blüten zur Spitze hin glatt.
Die männlichen Blüten variieren zwischen den Arten stärker als die weiblichen Blüten. Vor allem das Staubblatt ist oft sehr divergent. Die Staubbeutel stehen zur Achse hin und die Staubfäden sind flach und mehr oder weniger verdickt. Nach der Gestalt des Staubblatts werden die männlichen Ameisenbaumblüten in fünf Gruppen geteilt:
- Cecropia peltata-Typ: Der häufigste Typus. Die Antheren lösen sich von den Staubfäden kleben jedoch an der Blütenöffnung fest und bleiben dort sitzen.
- Cecropia latiloba-Typ: Bei der Anthese lösen sich die Antheren fast vollständig von den Staubfäden. Die Blüte ist weiter geöffnet und die ganzen Staubbeutel können vom Wind davongetragen werden.
- Cecropia sciydophylla-Typ: Die Blüten und Staubbeutel sind in die Länge gezogen und konvex gebogen. Die Antheren lösen sich von den Staubfäden, bleiben aber durch Tracheiden mit der Blüte verbunden.
- Cecropia heterochroma-Typ: Die Staubbeutel zeigen aus der Blüte heraus und bleiben mit den Staubfäden verbunden.
- Cecropia membranaceae-Typ: Die Antheren sind mit den Filamenten verwachsen. Die Blüte ist stark verkürzt.
Die Pollen sind trocken und werden oft in großen Wolken verweht. Cecropia-Pollen in Sedimentgestein wurden zu paläoökologischen Studien herangezogen.[5]
Weibliche Blüten
Die weiblichen Blüten stehen bei den meisten Arten frei, zum Beispiel bei C. peltata oder C. litoralis sind sie jedoch an der Basis verwachsen. Die Blütenhülle ist zu einer Röhre verwachsen. Der obere Teil der Blüten ist verdickt und konvex gebogen. Bei allen Arten ist die Außenseite der Blüte mit spinnenartigen Haaren besetzt. Die Blüte ist fast vollständig verwachsen und es verbleibt nur ein Spalt, der groß genug ist um die Narbe hindurch zu lassen. Die weiblichen Blüten besitzen oberständigen Fruchtknoten aus einem einzigen Fruchtblatt. Die Narbe ist entweder kopfig-pinselig oder gestielt und ungeflügelt.
Früchte und Samen
 Fruchtstände eines Ameisenbaums
Fruchtstände eines AmeisenbaumsDie Früchte sind Achänen, eine Sonderform der Nussfrucht, sie werden von vergrößerten, mehr oder weniger fleischigen Teilen der Blütenhülle umfasst. Sie sind grün und bleiben auch nach vollständiger Reife grün. Ihre Form ist länglich, ellipsoid, eiförmig oder umgekehrt eiförmig. Das Perikarp, das Fruchtgehäuse, ist zumeist glatt und dunkelgrün bis braun. Die Samenschale ist sehr dünn. Der Embryo mit zwei gleichen, flachen Keimblättern liegt eingebettet in Endosperm. Die Samen verfügen über eine hohe Keimfähigkeit, und können lange im Boden überdauern. Die Keimung wird durch Sonnenlicht und Temperaturwechsel ausgelöst.
Verbreitung
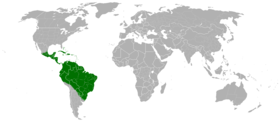 Verbreitungsgebiet der Ameisenbäume (ohne neophytische Vorkommen)
Verbreitungsgebiet der Ameisenbäume (ohne neophytische Vorkommen)Die Gattung ist in fast der gesamten Neotropis, das heißt im tropischen Teil Amerikas verbreitet. In Argentinien kommt nur eine Art (Cecropia pachystachya) im äußersten Nordosten des Landes vor; im Altiplano von Bolivien und Peru fehlt die Gattung; in Uruguay gibt es kleine Bestände. Die Nordgrenze der Verbreitung reicht bis über die Grenze des Bundesstaats Veracruz in Mexiko. Die meisten Ameisenbäume leben im Flachland bis 1000 m, einige montane Arten kommen aber in Nebelwäldern in Höhenlagen bis 2600 m vor.
Die Art Cecropia peltata hat sich als Neophyt auch auf andere Kontinente ausgebreitet. Zum Beispiel gibt es Bestände in Malaysia[6] und in der Elfenbeinküste[7]
Generell ist die Gattung weit verbreitet, dennoch werden einige Arten von der IUCN als bedroht eingestuft, so zum Beispiel C. pastasana oder C. multiflora. Auch einige Arten mit einem sehr kleinen Verbreitungsgebiet, wie C. multisecta, oder mit einer hohen Spezialisierung, wie etwa C. putumayonis und C. utcubambana sind prinzipiell gefährdet.
Ökologie
Ameisenbäume haben einen hohen Lichtbedarf und wachsen als Pionierpflanzen in Sekundärwäldern, auf Lichtungen, an Flussläufen, oder heute entlang von Straßen, die durch den Wald gebaut wurden. Sie wachsen sehr schnell, etwa 2,40 Meter pro Jahr, und können zum Beispiel eine nach einem Hochwasser neu entstandene Sandbank in kurzer Zeit besiedeln. Die hohlen Stämme und Äste sind wahrscheinlich eine Anpassung an das schnelle Höhenwachstum, der Baum investiert in das Höhenwachstum und verzichtet auf einen massiven Stamm, um nicht durch konkurrierende Bäume abgeschattet zu werden.
Ameisenbäume sind getrenntgeschlechtlich. Weibliche Bäume produzieren bis zu 900.000 Samen in einem Jahr. Die Samen bleiben vier bis fünf, oder sogar bis zu neun Jahre keimfähig. Bei Untersuchungen in Suriname fanden sich abhängig von der Art im Schnitt zwischen 20 und 80 Cecropia-Samen pro Quadratmeter Urwaldboden. Wegen der Rodung der Regenwälder durch den Menschen gibt es immer mehr Sekundärwälder und die Ameisenbäume haben sich stark ausgebreitet. [8]
Bestäubung
Die Morphologie der männlichen Blüten und Blütenstände weist darauf hin, dass Ameisenbäume windblütige Pflanzen sind. Dazu können entweder ganze Ähren vom Wind verweht werden oder die Antheren reichen aus der Blüte hinaus und die Pollen werden durch Bewegungen der Blüte herausgeschüttelt. Die Pollen sind trocken und können leicht verweht werden.
Dennoch gibt es auch Hinweise auf Entomogamie, obwohl Transport der Pollen durch Insekten bislang nicht beobachtet werden konnte. In den Blütenständen legen kleine Käfer und Fliegen ihre Eier ab.[9] Schwarzkäfer der Gattungen Epitragus und Ophtalmoborus ernähren sich von den Pollen von C. pachystachya, ob dabei eine Bestäubung stattfindet ist jedoch unklar. Die weiblichen Blüten der Art bilden kleine Mengen an Nektar.[10] Wahrscheinlich dient der Nektar aber eher zum besseren Ankleben der Pollen, als zum Anlocken von Insekten.
Symbiose mit Ameisen
 Cecropia pachystachya
Cecropia pachystachyaInsgesamt 48 der 61 Arten der Ameisenbäume (Siehe Abschnitt Systematik) leben mit Ameisen der Gattung Azteca in einer speziellen Symbiose, der Myrmekophylaxis. Beide Parteien können auch ohne den Symbiosepartner überleben, die Symbiose ist also fakultativ. Die Stängelwand der Cecropia ist über den Blattansätzen an den Prostomata sehr dünn und kann von den Ameisen leicht durchbissen werden. Die Kammern werden dann als Wohnraum verwendet. Etwa 60 % der Ameisenvölker halten Kulturen von Napfschildläusen (Coccidae) in den hohlen Sprossachsen, die sich ausschließlich vom Phloemsaft der Pflanzen ernähren und von den Ameisen „gemolken“ werden. Dabei greifen ältere Völker offenbar stärker auf solche Kulturen zurück als jüngere. Wahrscheinlich bringen die Ameisen ab einer bestimmten Größe ihrer Kolonie die Schildläuse in die Pflanze ein.[11] An der Unterseite der Blattstielbasis auf dem Trichilium werden zudem Futterkörperchen ausgebildet, die Protein- und Fettlieferanten sind und vor allem von den Azteca-Larven genutzt werden. Sie werden nach ihrem Entdecker Johann Friedrich Theodor Müller als Müllersche Körperchen bezeichnet. Sind keine Schildlauskulturen vorhanden, werden diese auch intensiver von adulten Ameisen genutzt.[12]
Drei Arten werden von Knotenameisen (Myrmicinae) der Gattung Crematogaster bewohnt, die auch symbiotisch mit Bäumen der paläotropischen Gattung Macaranga zusammenleben.[13] Die Internodien im unteren Teil der Sprossachse vieler myrmekophylaktischer Arten werden häufig von einer Vielzahl von Gattungen anderer Ameisen bewohnt. Darunter befinden sich wiederum Knotenameisen, zum Beispiel Feuerameisen (Solenopsis), Pheidole, Wasmannia oder Procryptocerus aber auch Urameisen (Ponerinae) wie Pachycondyla oder Schuppenameisen wie Camponotus und Myrmelachista. Auch die Gattung Pseudomyrmex, die mit Akazien als Ameisenpflanzen in Myrmekophylaxis lebt, und noch andere Ameisen finden sich hier.[14]
Die Ameisen verteidigen die Bäume gegen Schädlinge und Fressfeinde, z.B. gegen Blattschneiderameisen der Gattung Atta. Klettern Fressfeinde, bzw. Herbivoren, auf die Pflanze werden diese meist aggressiv von den anwesenden Azteca Ameisen attackiert und vertrieben.
Außerdem befreien die Ameisen die Ameisenbäume von Aufwuchs, wie Epiphyten und Kletterpflanzen. Beides würde die lichthungrigen und leichtgebauten Bäume durch Abschattung und das zusätzlich zu tragende Gewicht belasten.[8] So sind zum Beispiel Riemenblumengewächse auffällig selten auf Ameisenbäumen zu finden, und wenn dann nur sehr lokal, aber dort reichlich.[15]
Ein Nebeneffekt der Myrmekophylaxis ist ein erhöhtes Aufkommen von Spechten an den Ameisenbäumen. Die Vögel können die Ameisen an den Bäumen leicht fressen und beschädigen dabei auch die Stämme. Die Azteca erhalten somit Wohnraum und Nahrung, sind aber einem erhöhten Risiko durch Spechte gegenüber bodenlebenden Ameisen ausgesetzt. Vereinzelt wurden auch Erzwespen der Gattung Conoaxima beobachtet, die den Azteca zusetzen.[16] Der durch die Aggressivität gegenüber den Pflanzenschädlingen erforderliche hohe Energieverbrauch ist für junge Völker außerdem mit einer hohen Sterblichkeit verbunden. Die Bäume müssen Energie für die Müllerschen Körperchen zur Ernährung der Ameisen aufwenden. Darüber hinaus werden sie durch Spechte mehr beschädigt als andere Bäume. Dafür schützen die Ameisen sie aber sowohl vor Schädlingen als auch vor Kletterpflanzen und Epiphyten. Auch profitieren die Bäume durch die stickstoffreichen Ausscheidungen der Ameisen (Myrmekotrophie). Eine Untersuchung der Symbiose mit einem Doebeli-Knowlton-Modell ergab, dass die gegenseitigen Vorteile die Nachteile überwiegen. Junge Bäume profitieren aber stärker von der Verbindung als ältere Exemplare.[17] Neophytische Ameisenbäume müssen generell ohne Azteca-Ameisen auskommen, da diese nur in der Neotropis vorkommen.
Verhältnis zu anderen Tieren
 Das Dreifinger-Faultier zählt zu Fressfeinden der Ameisenbäume (hier auf C. insignis)
Das Dreifinger-Faultier zählt zu Fressfeinden der Ameisenbäume (hier auf C. insignis)Für Ameisenbäume ist das vertikale Wachstum vorrangig. So verzweigt sich die Sprossachse nicht, bis sie eine gewissen Höhe erreicht hat und bildet solange auch keine Blüten und Früchte zur generativen Vermehrung aus.[18] Vor allem junge Pflanzen bilden nur kleine Kronen mit vier bis zwölf Blättern aus; für sie ist viel Sonne sehr wichtig, eine Beschattung durch größere Pflanzen hingegen kritisch. Auch Fraß an den Blättern ist für junge Pflanzen gefährlich, verliert eine junge Pflanze ein Drittel ihrer Blätter, reduziert sich ihr Wachstum um den Faktor drei.[19]
Durch die Myrmekophylaxis sind die Ameisenbäume aber nicht vor allen Fressfeinden geschützt. Die großen auffälligen Blätter werden zum Beispiel besonders gerne von Dreifinger-Faultieren (Bradypus spp.) gefressen. Auch Raupen von Schmetterlingen ernähren sich von den großen Blättern, vor allem Hypercompe icasia, eine Art der Bärenspinner.
Rüsselkäfer der Gattung Pseudolechriops haben sich auf das Leben auf Ameisenbäumen spezialisiert. Sie verwenden lebende oder tote Blattstiele um ihre Eier abzulegen, die Larven wachsen dann in den Blattstielen auf. Einige Arten können die Azteca-Ameisen durch Mimikry imitieren und sind so vor ihnen geschützt.[20]
Insgesamt 33 Vogelarten aus elf Familien fressen die Früchte und Blüten von Cecropia-Arten.[8] 15 Arten von Vögeln ernähren sich von den Müller’schen Körperchen, dies sind zum Beispiel einige Arten der Schillertangaren (Tangara), einige Arten aus der Familie der Waldsänger (Parulidae), aber auch der Tukan-Bartvogel (Semnornis ramphastinus) und andere Vögel.[21]
Die Früchte werden auch von einer Vielzahl neotropischer Fledermäuse gefressen. Im Gegensatz zum Verbiss durch Faultiere ist dies für die Pflanzen jedoch von Vorteil, sie können ihre Samen durch die Tiere ausbreiten (Zoochorie). Dabei wird der fleischige Kelch sowie der äußere Teil der Frucht (Exokarp) und der mittlere Teil (Mesokarp) zum Teil verdaut, viele Samen passieren den Verdauungstrakt der Fledermäuse aber unbeschädigt. Im Boden bleibt die Keimfähigkeit die Samen über ein Jahr lang unbeeinträchtigt.[22] Aber auch Fische scheinen an der Samenausbreitung beteiligt zu sein, indem sie ins Wasser gefallene Früchte fressen.[23]
Ebenfalls unschädlich für die Ameisenbäume sind die Angela-Waldsänger (Dendroica angelae) auf Puerto Rico. Sie bauen ihre Nester aus den großen trockenen Blättern der Gattung.
Systematik
Traditionell wurde die Gattung den Maulbeergewächsen (Moraceae) zugerechnet.[24] Im Jahr 1978 stellte Cornelis Christiaan Berg eine eigene Familie Cecropiaceae auf, die neben den Ameisenbäumen noch fünf weitere Gattungen (zum Beispiel Coussapoa und Pourouma) umfasste.[25] Genetische Untersuchungen geben aber Hinweis, dass die Gattung in die Familie der Brennnesselgewächse (Urticaceae) gehört.[26] Dieser Sichtweise schloss sich auch die Angiosperm Phylogeny Group an.[27]
Die Gattungen Poikilospermum und die Kanonierblumen (Pilea) sind Schwestertaxone. Diese beiden bilden nun wiederum eine Schwesterklade zu den Ameisenbäume. Diese drei Gattungen bilden eine Klade, die morphologisch viel Rückhalt hat, folgendes Kladogramm zeigt das Verwandtschaftsverhältnis noch einmal:[28]
┌────── Cecropia ─┤ ┌─── Poikilospermum └──┤ └─── PileaDer erste Versuch einer Einteilung der Gattung in Sektionen und Untersektionen stammt von Emil Heinrich Snethlage aus dem Jahr 1923.[29] Er teilt die Gattung, wie folgt, in zwei Sektionen und acht Untersektionen:
- I. Sektion Tomentosae
- A. Aequales
- B. Subaequales
- C. Arachnoidae
- II Sektion Atomentosae
- D. Centrales
- E. Angulatae
- F. Elongatae
- G. Abbreviatae
- H. Polystachyae
Mehrere Arten wie C. montana finden jedoch in dieser Einteilung keinen Platz. Cornelis Christiaan Berg schlug deshalb 1990 eine Einteilung in nur zwei größere Gruppen ohne taxonomischen Rang vor.
- I. Cecropia peltata-Gruppe
- II. Cecropia telenitida-Gruppe
Nach der letzten monographischen Bearbeitung der Gattung umfasst sie 61 Arten.[30] Die Arten sind:
Arten der Gattung CecropiaArt Myrmekophylaxis C. alibicans Trécul ja, aber nicht im ganzen Verbreitungsgebiet C. andina Cuatrec nein C. angulata I.W.Bailey ja C. angustifolia Trécul ja, bis auf eine Unterart ohne M. im Süden Ecuadors C. annulata C.C.Berg & P.Franco ja C. bullata C.C.Berg & P.Franco nein C. chlorostachya C.C.Berg & P.Franco wahrscheinlich nicht C. concolor Willd. ja C. distachya Huber ja C. elongata Rusby ja C. engleriana Snethl. ja C. ficifolia Snethl. ja C. gabrielis Cuatrec. nein C. garciae Standl. ja C. glaziovii Snethl. ja C. goudotiana Trécul ja C. granvilleana C.C.Berg ja C. herthae Diels ja C. heterochroma C.C.Berg & P.Franco ja C. hispidissima Cuatrec. ja, aber mit einer anderen Ameisengattung C. hololeuca Miq. nein C. idroboi Cuatrec. ja C. insignis Liebm. ja C. kavanayensis Cuatrec. ja C. latiloba Miq. ja C. litoralis Snethl. ja C. longipes Pittier ja C. marginalis Cuatrec. ja C. maxima Snethl. ja C. megastachya Cuatrec. ja, aber mit einer anderen Ameisengattung C. membranacea Trécul ja, manchmal aber auch mit einer anderen Ameisengattung C. metensis Cuatrec. ja C. montana Sneth. ja C. multisecta P.Franco & C.C.Berg ja C. mutisiana Mildbr. ex Cuatrec. ja C. obtusa Trécul ja C. obtusifolia Bertol. ja C. pachystachya Trécul ja C. palmata Willd. ja C. pastasana Diels ja C. peltata L. ja, aber nicht im ganzen Verbreitungsgebiet C. pittieri B.L.Rob. nein C. plicata Cuatrec. ja, meistens C. polystachya Trécul ja C. purpurascens C.C.Berg ja C. putumayonis Cuatrec. ja C. reticulata Cuatrec. ja C. sararensis Cuatrec. ja C. saxatilis Snethl. ja C. schreberiana Miq. nein C. sciadophylla C.Mart. nein C. silvae C.C.Berg nein C. strigosa Trécul ja C. subintegra Cuatrec. ja C. tacuna C.C.Berg & P.Franco nein C. telealba Cuatrec. nein C. telenitida Cuatrec. ja, aber nicht im ganzen Verbreitungsgebiet C. ulei Snethl. ja C. utcubambana Cuatrec. ja C. velutinella Diels wahrscheinlich nicht C. virgusa Cuatrec. ja Etymologie
Der deutsche Trivialname leitet sich von der Myrmekophylaxis und somit vom engen Verhältnis der Gattung zu Ameisen ab.
Die Benennungsgeschichte der lateinischen Gattungsbezeichnung ist unklar. Vielfach wurde davon ausgegangen, dass sie vom lateinischen Namen Cecrops, altgriechisch Κέκροψ Kékrops stammt, ohne dass sich ein Bezug zum Ameisenbaum ergibt. Kekrops war ein Autochthon, das heißt ein Sohn der Gaia. Er war ein Mischwesen aus Mensch und Schlange.[31]
Eine plausiblere Theorie bezieht sich auf Kekrops II., den Sohn des Erechtheus und frühem König von Attika. Das erste Exemplar der Echten Feige (Ficus carica), einem Maulbeergewächs, soll in Attika gestanden haben. Auch die Gattung der Ameisenbäume wurde früher zu den Maulbeergewächsen gezählt, so dass eine Anlehnung daran sinnvoll erschien.[32]
Inhaltsstoffe und Wirkungen
Keiner der Bestandteile von Ameisenbäumen ist für den Menschen giftig, die Blätter sind jedoch zum Teil scharfkantig und können die Haut leicht einschneiden. Die Azteca-Ameisen an den Bäumen beißen Menschen. Ihre Bisse führen zu juckenden Quaddeln.
Die Inhaltsstoffe von Ameisenbäumen sind noch unzureichend erforscht, am besten bekannt sind Wirkstoffe aus Cecropia peltata, die Blätter enthalten Ergomitrin, ein Mutterkornalkaloid, Oxytocin, Serotonin sowie Acetylcholin und Prostaglandine. Viele dieser Stoffe können Wehen auslösen. So geben Bauern in Honduras Tieren während der Geburt Blätter von Cecropia peltata, um die Niederkunft zu beschleunigen, oder um das Lösen der Nachgeburt zu fördern.[33]
Die Blätter enthalten aber auch reizende Stoffe wie 1,8-Cineol, α-Pinen, Borneol, Eugenol, Ölsäure oder Vanillin. Einige der Inhaltsstoffe wie 1,8-Cineol, Serotonin, α-Pinen und β-Pinen können beim Menschen Spastiken auslösen.[34]
Ein Extrakt aus den Blättern wirkt gegen Gonorrhoe.[35] Traditionell werden Ameisenbäume vor allem in Mexiko, aber auch in anderen Teilen Süd- und Mittelamerikas gegen eine Vielzahl von Erkrankungen verwendet.[36] Cecropia pachystachya hat eine sedierende und positiv inotrope (herzstärkende) Wirkung und wird in Argentinien zur Behandlung von Asthma eingesetzt.[37]
Verwendung
Das Holz von Ameisenbäumen ist sehr leicht und biegsam. C. peltata hat zum Beispiel eine Relative Dichte von 0,29 in Bezug auf Wasser, dies ist nur leicht mehr als bei den meisten Balsahölzern. Dennoch ist es in der Verwendung eingeschränkt und wird nur lokal genutzt.[38] Es wird vor allem zur Herstellung von Musikinstrumenten und Werkzeugstielen verwendet. Zum Beispiel werden in Nariño traditionelle Klanghölzer aus Ameisenbaumholz gefertigt, aber auch Flöten oder Gitarren werden aus dem Holz hergestellt.
Darüber hinaus wird das Holz zur Fertigung von Streichhölzern und billigen Kisten verwendet. Seltener werden die halbierten, hohlen Stämme als Wasserleitungen genutzt. Es wurde versucht das Holz zur Papierherstellung heranzuziehen und es gibt einige Anlagen die Holzstoff aus Cecropia herstellen. Wegen seines hohen Harz- und Milchsaft-Gehaltes ist das Holz dazu aber nur schlecht geeignet.[39] In Puerto Rico wird das geschredderte Holz mit Zement vermischt als Baustoff oder so als Dämmstoff verwendet.[40]
Der Hauptnutzen von Ameisenbäumen liegt in der Aufforstung. Sie stellen geringe Ansprüche an den Boden und wachsen sehr schnell. So können von Bodenerosion bedrohte Gebiete befestigt werden. Der Boden wird zunächst zurückgehalten und neue Biomasse eingebracht, sodass sich auch andere Arten wieder ansiedeln können.[41]
Fasern der Rinde können zu Seilen gedreht werden. Aus solchen Seilen werden unter Anderem Bogensehnen und Hängematten gefertigt. Die Fruchtstände sind essbar und werden in Bolivien unter dem Namen „bananitas“ gehandelt. Das Mark der Blattstiele wird in Napo in Ecuador für die Herstellung traditionellen Kopfschmucks verwendet.
Traditionell werden die frischen Blätter von Ameisenbäumen verbrannt und die Asche, mit gerösteten und pulverisierten Coca-Blättern gemischt. Dieser Priem wird zwischen Wange und Zahnfleisch unter die Zunge gelegt.[42] Hierfür werden die Blätter von C. ficifolia, C. palmata, C. peltata und C. sciadophylla verwendet.[43]
Literatur
- Cornelis Christiaan Berg, Pilar Franco Rosselli: Cecropia (Cecropiaceae). In: Flora Neotropica Monographs. 94, 2005, ISBN 978-0-89327-461-0.
- Sandra Burger: Vergleichende Bestandsaufnahme von Ameisenpflanzen entlang eines Transekts im tropischen Tieflandregenwald Costa Ricas. Diplomarbeit an der Universität Wien, Wien September 2003 (pdf ; Stand: 17. Juli 2007).
- Servando Carvajal, Luz María Gonzáles-Villarreal: La familia Cecropiaceae en el estado de Jalisco, México. 1. Auflage. Universidad de Guadalajara, Guadalajara 2005, ISBN 978-970-27-0683-0 (pdf).
Einzelnachweise
- ↑ J. Jeník, D. H. Sen: Morphology of root systems in trees: a proposal for terminology. In: Tenth International Botanical Congress, Edinburgh. Abstracts. 1964, S. 393–394.
- ↑ Jindrich Pavlis, Jan Jeník: Roots of pioneer trees in the Amazonian rain forest. In: Trees - Structure and Function. 4, Nr. 18, 5. August 2000, S. 442–455 (doi:10.1007/s004680000049) (Stand: 2. August 2007).
- ↑ Andreas Franz Wilhelm Schimper: Wechselbeziehungen zwischen Pflanzen und Ameisen im tropischen Amerika. In: Botanische Mitteilungen aus den Tropen. 1, G. Fischer, Jena 1888, S. 25 (txt ; Stand: 17. Juli 2007).
- ↑ Fred R. Rickson: Ultrastructural Differentiation of the Mullerian Body Glycogen Plastid of Cecropia peltata L.. In: American Journal of Botany. 63, Nr. 9, Oktober 1976, S. 1272–1279 (doi:10.2307/2441743).
- ↑ Kam-Biu Liu, Paul A. Colinvaux: A 5200-Year History of Amazon Rain Forest. In: Journal of Biogeography. 15, Nr. 2, März 1988, S. 231–248 (doi:10.2307/2845412).
- ↑ F. E. Putz, N. M. Holbrook: Further observations on the dissolution of mutualism between Cecropia and its ants: the Malaysian case. In: Oikos. 53, 1988, S. 121–125.
- ↑ L. Ake Assi: Cecropia peltata Linne (Moracees): ses origines, introduction et expansion dans l'est de la Cote d'Ivoire. In: Serie A. Sciences Naturelles. 42, Institut Fondamental d'Afrique Noire, 1980, S. 96–102.
- ↑ a b c John Kricher: A Neotropical Companion. Princeton University Press, 1999, ISBN 0691009740, S. 71–73.
- ↑ J. C. Andrade: Observações preliminares sobre a eco-etologia de quatro coleópteros Chrysomelidae, Tenebrionidae, Curculionidae) que dependem da embaúba (Cecropia lyratiloba var. nana – Cecropiaceae), na restinga do Recreio dos Bandeirantes, Rio de Janeiro. In: Rev. Bras. Entomol.. 28, 1984, S. 99–108.
- ↑ C. de Andrade: Epizootia natural causada por Cordyceps unilateralis (Hypocreales, Euascomycetes) em adultos de Camponotus sp. (Hymenoptera, Formicidae) na região de Manaus, Amazonas, Brasil. In: Acta Amazonica. 10, 1980, S. 671–677.
- ↑ A. Benjamin, M. Giambruno, M. Lyke, A. B. Pyle, R. Warner: A lesson in ABC’s of La Selva (Ants, Bugs, Cecropia). Abgerufen am 21. Juni 2007.
- ↑ Dennis J. O'Dowd: Pearl Bodies as Ant Food: An Ecological Role for Some Leaf Emergences of Tropical Plants. In: Biotropica. 14, Nr. 1, März 1982, S. 40–49 (doi:10.2307/2387758).
- ↑ Jerry F. Downhower: The Distribution of Ants on Cecropia Leaves. In: Biotropica. 7, Nr. 1, April 1975, S. 59–62 (doi:10.2307/2989801).
- ↑ The Cecropia-Azteca association in Costa Rica. John T. Longino. Abgerufen am 22. August 2007.
- ↑ P. Jolivet: Relative protection of Cecropia trees against leaf-cutting ants in tropical America. In: R. K. Vander Meer, K. Jaffe, A. Cedeno (Hrsg.): Applied myrmecology: a world perspective. Westview Press, Boulder 1990, S. 251–254.
- ↑ W. M. Wheeler: Studies of neotropical ant-plants and their ants. In: Bulletin of the Museum of Comparative Zoology. 90, Harvard 1942, S. 1–262.
- ↑ The Costs and Benefits of the Cecropia-Azteca-Coccidae Symbiosis. David Logue. Abgerufen am 21. Juni 2007.
- ↑ Daniel H. Janzen: Dissolution of mutualism between Cecropia and its Azteca ants. In: Biotropica. 5, 1973, S. 15–28.
- ↑ Jerry F. Downhower: The Distribution of Ants on Cecropia Leaves. In: Biotropica. 7, Nr. 1, April 1975, S. 59–62.
- ↑ Henry A. Hespenheide, Louis M. Lapierre: A review of Pseudolechriops Champion (Coleoptera: Curculionidae: Conoderinae). In: Zootaxa. 1384, 2006, ISSN 1175-5326, S. 1–39 (http://www.mapress.com/zootaxa/2006f/zt01384p039.pdf ; Stand: 31. Juli 2007).
- ↑ Kazuya Naoki, Efraín Toapanta: Müllerian Body Feeding by Andean Birds: New Mutualistic Relationship or Evolutionary Time Lag?. In: Biotropica. 33, Nr. 1, 2001, S. 204–207.
- ↑ Tatyana A. Lobova, Scott A. Mori, Frédéric Blanchard, Heather Peckham, Pierre Charles-Dominique: Cecropia as a food resource for bats in French Guiana and the significance of fruit structure in seed dispersal and longevity. In: American Journal of Botany. 90, 2003, S. 388–403 (http://www.amjbot.org/cgi/content/abstract/90/3/388).
- ↑ G. Gottsberger: Seed dispersal by fish in the inundated regions of Humaita, Amazonia. In: Biotropica. 10, 1978, S. 170–183.
- ↑ W. Burger (Hrsg.): Flora Costaricensis, Family #52, Moraceae. Fieldiana, Botany 40. 1977, S. 94–215.
- ↑ Cornelis Christiaan Berg: Cecropiaceae, a new family of Urticales. In: Taxon. 27, 1978, S. 39–44 (doi:10.2307/1220477).
- ↑ W. S. Judd, R. W. Sanders, M. J. Donoghue: Angiosperm family pairs: preliminary phylogenetic analyses. In: Harvard papers in Botany. 5, 1994, S. 1–51.
- ↑ The Angiosperm Phylogeny Group: An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II. In: Botanical Journal of the Linnean Society. 141, Nr. 4, 2003, S. 407 (doi:10.1046/j.1095-8339.2003.t01-1-00158.x) (Stand: 2. August 2007).
- ↑ W. S. Judd, R. W. Sanders, M. J. Donoghue: Angiosperm family pairs: preliminary phylogenetic analyses. In: Harvard papers in Botany. 5, 1994, S. 27–28.
- ↑ Emil Heinrich Snethlage: Beiträge zur Kenntnis der Gattung Cecropia und ihrer Beziehungen zu den übrigen Conocephaloideen. Friedrich-Wilhelms-Universität Berlin, Berlin 1923.
- ↑ Cornelis Christiaan Berg, Pilar Franco Rosselli: Cecropia (Cecropiaceae). In: Flora Neotropica Monographs. 94, 1990, ISBN 0893274615.
- ↑ Helmut Genaust (Hrsg.): Etymologisches Wörterbuch der botanischen Pflanzennamen. 3. Auflage. Nikol, Hamburg 2005, ISBN 3937872167, S. 136f.
- ↑ V. Hehn: Kulturpflanzen und Haustiere in ihrem Übergang von Asien nach Griechenland und Italien sowie in das übrige Europa. 8. Auflage. Bornträger, Berlin 1911 (mit historisch linguistischen Studien von O. Schrader und botanischen Beiträgen von F. Pax).
- ↑ J. Paquet: Manual de Dendrologia de Algunas Especies de Honduras. Programa Forestal ACDI-COHDEFOR, 1981.
- ↑ Eintrag in der Nutrient Network Library. Abgerufen am 21.06.2007.
- ↑ A. Caceres, H. Menendez, E. Mendez, E. Cohobon, B. E. Samayoa, E. Jauregui, E. Peralta, G. Carrillo: Antigonorrhoeal activity of plants used in Guatemala for the treatment of sexually transmitted diseases. In: Journal of Ethnopharmacology. 48, 1995, S. 85–88.
- ↑ Treating Livestock with Medicinal Plants: Cecropia spp.. Abgerufen am 21.06.2007.
- ↑ Alicia E. Consolinia, María Inés Ragonea, Graciela N. Miglioria, Paula Confortib, María G. Volonté: Cardiotonic and sedative effects of Cecropia pachystachya Mart. (ambay) on isolated rat hearts and conscious mice. In: Journal of Ethnopharmacology. 106, Nr. 1, 15. Juni 2006, S. 90–96 (doi:10.1016/j.jep.2005.12.006).
- ↑ Justiniano Velazquez: Contribución al conocimiento de las especies del género Cecropia L. Moraceae-"Yagrumbos" de Venezuela. In: Acta Botánica Venezolana. 6, 1971, S. 25–64.
- ↑ E. L. Keller, R. M. Kingsbury, D. J. Fahey: Neutral sulfite semichemical pulping of guaba (Inga vera), yagrumo hembra (Cecropia peltata) and eucalyptus (Eucalyptus robusta) from Puerto Rico. In: USDA Forest Service, Report 2127. Forest Products Laboratory, Madison, WI 1958, S. 7.
- ↑ W. S. Chalmers: Observations on some Caribbean forests. In: Caribbean Forester. 19, Nr. 12, 1958, S. 30–42.
- ↑ Susan R. Silander, Ariel E. Lugo: Cecropia peltata L. Yagrumo Hembra, Trumpet-Tree. In: Silvics of North America. 2, United States Department of Agriculture Forest Service, Washington, DC 1990, S. 496–497 (http://www.na.fs.fed.us/spfo/pubs/silvics_manual/volume_2/silvics_v2.pdf ; Stand: 6. August 1007).
- ↑ Richard Evans Schutes: A new method of coca preperation in the Columbean Amazon. In: Botanical Museum Leaflets. 17, Harvard University, 1957, S. 241–264.
- ↑ Christian Rätsch, Jonathan Ott: Coca und Kokain, Ethnobotanik, Kunst und Chemie. AT Verlag, Aarau, Schweiz 2003, ISBN 3855027072.
Weblinks
- Longino, J.T. (2005): The Cecropia-Azteca association in Costa Rica. Artikel über die Myrmekophylaxis Crecropia <-> Azteca (Englisch)
- Cecropia in Flora de Nicaragua (Spanisch)
- Cecropia in Peru (PDF) (Englisch)
- Eintrag in der Flora der Anden von Ecuador (Spanisch)
- Eingescannte Herbarbelege aus dem Field Museum (Chicago) (Englisch)
- Schöne Detailbilder auf ruhr-uni-bochum.de (Deutsch)






Wikimedia Foundation.