- Aufsteigendes retikuläres aktivierendes System
-
Das aufsteigende retikuläre Aktivierungssystem (ARAS) ist in der Neurobiologie ein Teil des Formatio reticularis.
Inhaltsverzeichnis
Der Hirnschrittmacher
Der Hirnforscher Rolf Hassler schrieb 1971 einen Beitrag für den Sammelband Hirnforschung und Psychiatrie im Colloquium Verlag Berlin mit der Überschrift: Regulation der psychischen Aktivität. Darin beschrieb er das Schlaf-Wach-System oder „unspezifische Aktivierungssystem“ der reticulären Formation.
Demnach existieren zwei Aktivierungswirkungen der Formatio reticularis, eine „tonische“ und eine „phasische“. Das tonische System in der Formatio reticularis des Mittelhirns reguliert über den Hypothalamus die Ausscheidung der Neurotransmitter (Nor-)Adrenalin und Serotonin, die zur lange anhaltenden, tonischen Aktivierung und Dämpfung der corticalen Aktivität führen und z. B. den Tag-Nacht-Rhythmus beeinflussen.
Das phasische System für kurzzeitige Aktivierungen einzelner Rindenanteile hat sein Zentrum im medialen Thalamus. Wie eine Hülle liegen die reticulären Strukturen um die spezifischen Kerne der Sinnesorgane, die vom Thalamus in die sensorischen Projektionszentren der Hirnrinde führen. Die unspezifischen, reticulären Strukturen werden hier durch Nebenleitungen sämtlicher Sinnessysteme konvergent informiert und in Gang gesetzt, besonders von Impulsen aus dem Hautsinn und aus dem Gleichgewichtsorgan.
Diese unspezifischen Schalteinheiten des Thalamus haben eine geringere Allgemeinwirkung auf die zugehörige Großhirnhälfte als die reticuläre Formation des Mittelhirns. Sie wirken viel selektiver auf einzelne Rindenfelder und können sogar gleichzeitig andere Rindenfelder abschirmen.
Von den retikulären Thalamuskernen ist bekannt, dass sie nicht direkt zur Großhirnrinde leiten, sondern über Zwischenstationen erst nach 20-40 msec in den Rindenfeldern zur Wirkung kommen. Eine Erklärung für diese Verzögerung findet Hassler in Studien über Faserverläufe, die einen Umweg der Erregungen zu bestimmten Kernen der Stammganglien nachweisen, von dem sie durch zahlreiche Zwischenneurone verzögert wieder zu bestimmten Thalamuskernen zurückkehren, um nun auf schnelleren Leitungswegen die Großhirnfelder zu beeinflussen.
Die unspezifischen Zentralkerne des Thalamus leiten zum Nucleus caudatus und zum Putamen, die beide regulatorisch auf die Motorik wirken.
Aus der Hüllschicht des Thalamus gehen Verbindungen zum Pallidum. Von diesen blassen Stammganglienkernen geht eine Leitung zurück zu den spezifischen Thalamuskernen. Erst nach dieser Leitungsschleife über die Basalganglien erreichen die unspezifischen Erregungen die Hirnrinde und wirken hier über Sternzellen auf die strickleiterförmigen Dendriten der Pyramidenzellen.
Als Ergebnis der unspezifischen Erregungsschleifen über die Basalganglien sieht Hassler die elektrische Grundaktivität der Hirnrinde, die im EEG registrierbar ist und die psychische Aktivität vom Schlaf bis zur angespannten Erregung beeinflusst.
Der Thalamo-corticale Schaltplan
 Thalamo-corticaler Schaltplan
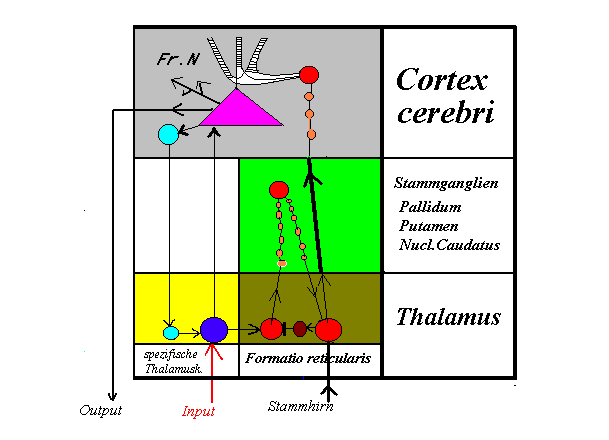
Thalamo-corticaler SchaltplanDiese Skizze ist nach der Beschreibung Hasslers gezeichnet und soll die Verhältnisse zwischen Thalamus und Cortex anschaulich machen. Sie beschränkt sich aus Gründen der Übersichtlichkeit auf eine einzige der milliardenfachen Verbindungen, die vom Thalamus zum Großhirn und von diesem zurück in den Thalamus ziehen.
Die Berechtigung zu solcher Vereinfachung ergibt sich aus der Gleichartigkeit, mit der die verschiedenen Kerngebiete des Thalamus ihre Fasern in alle Teile des Cortex leiten. Auch im Cortex ist die Art der Verschaltung in allen Regionen gleichartig, wenn man das Verhältnis der Pyramidenzellen, Sternzellen usw. im Einzelnen betrachtet.
Als funktionale Einheit der Großhirnrinde soll eine der millionenfach in das Grundmuster eingewebten Pyramidenzellen gesehen werden. Diese Zellen sind nur in der Großhirnrinde zu finden und bilden mit den Sternzellen deren spezielle Architektur.
Diese funktionale Einheit besteht nicht allein aus der Pyramidenzelle und ihren corticalen Verbindungen, sondern sie umschließt auch Verbindungen vom Thalamus zum Cortex. Dazwischen liegen aber noch die Stammganglien, die den Erregungen der Formatio reticularis jenen merkwürdigen Umweg aufzwingen, der zum Thalamus zurückkehrt und dann erst mit 20–40 msec Verzögerung in die speziellen Projektionsgebiete der Rinde „feuert“.
Die Funktion des neuronalen Oszillators
Wenn ein Nachrichtentechniker den Schaltplan eines Gerätes verstehen will, beginnt er mit dessen Input, um den Weg der Signale zu verfolgen. Alle Sinnesorgane senden ihre Signale über den Thalamus zum Cortex, hier mit dem roten Input-Pfeil an der dunkelblauen Thalamuszelle eingezeichnet. Die dunkelblaue Zelle sendet das Ergebnis der bisherigen Analyse zu einer der vielen Pyramidenzellen im Cortex.
Gleichzeitig aktiviert diese Thalamuszelle über einen Seitenast die linke, rote Zelle der Formatio reticularis, welche die Erregungen zunächst zu den Stammganglien sendet. Diese Leitung ist nach Hassler sehr langsam, weil sie über etliche Zwischenneurone geht, und sie macht eine Schleife zurück zum Thalamus (rechte rote Zelle), bevor sie von dort aus ihre spezielle Pyramidenzelle im Cortex aktivieren kann.
Hier muss gefragt werden, worin der Sinn dieser rückläufigen Schleife für die Erregungen des reticulären Aktivierungssystem liegen kann. Warum dieser Umweg, warum werden die reticulären Signale nicht auf direktem Weg zum Cortex geleitet? Um eine plausible Antwort auf diese Frage zu finden, ist es gut, dass das Ergebnis der reticulären Aktivität in der Hirnrinde ja bereits durch die vielen Untersuchungen der Hirnwellen (EEG) bekannt ist: Solange der Mensch am Leben ist, wird seine Hirnrinde durch das reticuläre Aktivierungssystem in elektrische Schwingungen versetzt, die vom Schlaf bis zur angespannten Aufmerksamkeit in der Frequenz zunehmen, in der Amplitude abnehmen. Bekannt ist seit den Untersuchungen von William Grey Walter auch, dass diese Wellen durch äußere Reize beeinflusst (getriggert) werden können. Damit wird ja die „Weck-Funktion“ des retikulären Systems erklärt. Starke Reize wecken uns aus dem Schlaf, lassen uns „aufschrecken“, indem sie über die Formatio reticularis die Hirnrinde in einen höheren Aktivierungszustand versetzen.
Wenn solche Reize rhythmisch im Frequenzspektrum der Hirnwellen auftreten, kommt es schnell zu einer Synchronisierung zwischen Reizrhythmus und Hirnwellen. Es muss also in diesem System zwischen Thalamus und Cortex eine Art von Oszillator vorhanden sein, der eine variable, über den sensorischen Input beeinflussbare Schwingung erzeugt.
Wird die Schaltung unter diesem Blickwinkel betrachtet, kann man leicht einen derartigen Schwingungsgenerator damit konstruieren. Es genügt, eine hemmende Zelle in den retikulären Kreislauf einzuschalten, wie sie hier in Form der dunkelroten Schaltzelle zwischen die Startzelle und die Zielzelle der thalamischen Formatio reticularis in das Bild eingezeichnet wurde.
Wenn diese Zelle von der rechten roten Zelle aktiviert wird, hemmt sie die linke rote Zelle und unterbricht den Signalstrom so lange, bis die rechte rote Zelle nicht mehr erregt ist und den Eingang wieder freigibt. So entsteht als Ergebnis an der rechten roten Zelle aus dem kontinuierlichen Input eine rhythmische Folge, die von dort aus in den Cortex gelangt. Ihre Periode entspricht der Zeit, welche die Nervenimpulse für die Schleife benötigen. Über die variable Leitfähigkeit der Zwischenneurone kann die Laufzeit der Schleife sehr fein reguliert und variiert werden. So kann die Entstehung der variablen EEG-Wellen mit dem einfachen Schaltplan grob skizziert werden.
Der Sinn der rhythmischen Erregung erschließt sich über die Vorgänge an den Pyramidenzellen. Sie sollen durch den Input der Sinnesorgane erregt werden und sich mit gleichzeitig erregten Zellen zu Komplexen verbinden. Jede Pyramidenzelle ist aber über unzählige Verbindungen mit allen Teilen des Nervensystems „verdrahtet“, so dass eine einzige Erregung sich wie ein Buschfeuer schnell auf das ganze Gehirn ausbreiten würde, wenn die Zellen immer in erregbarem Zustand wären. Damit wäre jede differenzierte Wahrnehmung oder Handlung unmöglich.
Wenn durch eine rhythmische Impulsfolge das Membranpotential aller Pyramidenzellen gemeinsam angehoben und abgesenkt wird, so entstehen in ihnen erregbare Momente und unerregbare Momente in stetigem Wechsel. Dadurch kann kein Aktionspotential überdauern oder sich grenzenlos ausbreiten, jede Erregung wird sofort wieder durch einen Löschvorgang vernichtet, um einer neuen Erregung Platz zu machen.
Die Möglichkeit zum Heben und Senken des Membranpotentiale ergibt sich bei den Pyramidenzellen über die strickleiterförmigen Kontakte mit den Sternzellen, welche von der Impulsfolge der Formatio reticularis rhythmisch erregt werden.Diese stickleiterförmigen (John Eccles nennt sie „patronengürtelförmig“) synaptischen Kontakte sind bei der Geburt noch nicht vorhanden, sie bilden sich besonders stark in den ersten Lebensjahren aus und scheinen damit zur Reifung des Großhirns im Zusammenhang mit Lernprozessen nötig zu sein. Vorstellbar ist, dass die rhythmischen Erregungen aus dem ARAS durch die strickleiterförmige Übertragung auf die Pyramidenzellen in der Stärke und im zeitlichen Verhalten über Neurotransmitter sehr fein reguliert werden können.
Mit diesen Überlegungen wird der Sinn der EEG-Wellen erkennbar: Die thalamische Formatio reticularis hebt und senkt in variablem Rhythmus das Membranpotential der kortikalen Pyramidenzellen. Sie wird dazu stimuliert durch die Stärke der Inputreize. Immer wenn das Membranpotential der Pyramidenzellen die „Zündschwelle“ erreicht, dann können die von den Sinnesorganen ankommenden spezifischen Sinnesreize gemeinsam erregt werden und spezifische Nervennetze hervorrufen, die wegen der selbstähnlichen Durchdringung aller Sinnesorgane aus vielen Teilkomplexen ein Ganzes entstehen lassen; eine ganzheitlich verbundene Struktur, die circa zehn Mal pro Sekunde neu gebildet wird.
In dem Schaltplan sind Hasslers Beschreibung entsprechend auch rückläufige Verbindungen von der Pyramidenzelle in die spezifischen Kerne des Thalamus gezeichnet. Damit lässt sich verstehen, dass das Gehirn nicht auf Eingangssignale angewiesen ist, um aktiv zu sein. Selbst wenn ein Mensch von allen Wahrnehmungen isoliert lebt, hat er Vorstellungen und Gedanken, mitunter sogar Halluzinationen, weil sein Cortex Aktivität wünscht und erzeugt. Im Schlaf oder in der Narkose werden die Hirnwellen langsamer, womit der Abstand zur „Zündschwelle“ für die Pyramidenzellen so groß wird, dass nur sehr starke „Weckreize“ zur Aktivierung der Wahrnehmung führen.
Der beschriebene Schrittmachermechanismus zeigt sich also von vielen Seiten sinnvoll. Er macht aber auch Störfälle denkbar, die in zwei pathologischen Varianten auftreten können:
- Der „Hirnschrittmacher“ kann die Pyramidenzellen durch Fehlsteuerung zu stark erregen, so dass deren „Zündschwelle“ auch ohne spezifische Sinnesreize von vielen Neuronen überschritten wird. Ein epileptischer Anfall wäre die Folge.
- Das gegensätzliche Extrem ist so denkbar, dass die rhythmische Erregung zu schwach ist und die „Zündschwelle“ kaum noch erreicht wird. Eine Hemmung oder Verlangsamung der Bewegungen, die sich bei vielen Störungen des retikulären Nervensystems unter anderem als Parkinson-Krankheit feststellen lässt, muss dann die Folge sein.
Künstliche Hirnschrittmacher
Seit wenigen Jahren werden vorwiegend bei der Parkinson-Krankheit therapeutische Versuche mit elektronischer Stimulation in den Basalganglien und im Nucleus subthalamicus durchgeführt, die zum Teil gute Erfolge bringen.
2007 haben amerikanische Neurochirurgen einem 38jährigen Mann, der nach einem Unfall sechs Jahre lang im Koma lag, einen Schrittmacher im Thalamus implantiert, mit dessen Wirkung der Patient sofort die sechs Jahre lang geschlossenen Augen öffnen und einige Wörter sprechen konnte.
Weblinks
- Wikibook Gehirn und Sprache
Literatur
- Hassler, Rolf: Regulation der psychischen Aktivität (in „Hirnforschung und Psychiatrie“ Colloquium Verlag Berlin 1971)
- Eccles, J.C.: Wie das Selbst sein Gehirn steuert, Berlin 1994
- Walter, William Grey.: The Living Brain, (1953), Penguin, London, 1967
Wikimedia Foundation.