- Seitenwurzeln
-
 Teilweise durch Bodenabtrag freigelegter Wurzelstock eines Baumes im Speyerer Auwald
Teilweise durch Bodenabtrag freigelegter Wurzelstock eines Baumes im Speyerer AuwaldDie Wurzel (von althochdeutsch wurzala „das Gewundene“) ist neben Sprossachse und Blatt eines der drei Grundorgane der Kormophyten, zu denen die Samenpflanzen und Farne zählen.
 Die Wurzeln einer hydroponisch gezogenen Pflanze
Die Wurzeln einer hydroponisch gezogenen PflanzeSie dient primär der Aufnahme von Wasser und den darin gelösten Mineralstoffen sowie der Befestigung der Pflanze an ihrem Standort. Vielfach wird die Wurzel durch Metamorphosen auch für andere Aufgaben genutzt, besonders häufig als Speicherorgan für Reservestoffe. Der von den Wurzeln beeinflusste Bereich des Bodens ist die Rhizosphäre.
Ferner sind Wurzeln ein Syntheseort für zahlreiche Pflanzenhormone (z. B. Cytokinin) und sekundäre Pflanzenstoffe (z. B. Alkaloide). Ein Beispiel ist das Nikotin in den Blättern der Tabakpflanze, das in der Wurzel synthetisiert wird.
Die wichtigsten Unterschiede zur Sprossachse sind:
- Die Wurzel verfügt über Wurzelhaare und eine Wurzelhaube (Kalyptra) an der Spitze.
- Die Verzweigungen sind endogenen Ursprungs.
- Die primären Leitbündel sind radial angeordnet.
- Wurzeln tragen keine Blätter.
Inhaltsverzeichnis
Wurzelsysteme
Als Wurzelsystem (Wurzelwerk oder Wurzelgeflecht) wird die Gesamtheit der Wurzeln einer Pflanze bezeichnet. Prinzipiell unterscheidet man verschiedene Arten von Wurzelsystemen:
- „heterogene Wurzelsysteme“ (Allorhizie), mit einer senkrecht nach unten wachsenden Hauptwurzel und seitlich von ihr abzweigenden Seitenwurzeln. Sie treten bei den Dikotylen auf.
- „homogene Wurzelsysteme“ (Homorhizie), die aus zahlreichen gleichrangigen, ähnlich gestalteten Wurzeln bestehen. Hierzu zählen die Farne und die Monokotylen.
Der Übergangsbereich zum Stamm eines Baumes ist in der Forstsprache der Wurzelstock, bei krautigen Pflanzen hingegen meint der Gärtner damit das Wurzelgerüst oder den Wurzelballen (das ganze Wurzelsystem einer Topfpflanze). Das Rhizom (umgangssprachlich „Wurzelstock“) hingegen gehört nicht zum Wurzel-, sondern zum Sprossachsensystem.
Die Ausgestaltung eines Wurzelsystems, das sogenannte Wurzelbild, hängt sehr stark von der Gründigkeit des Bodens ab. Bei Bäumen werden drei Grundtypen unterschieden:
- Pfahlwurzelsystem: eine senkrecht wachsende, verdickte Hauptwurzel dominiert (Eichen, Kiefern, Tannen, Ulmen).
- Herzwurzelsystem: Am Wurzelstock werden mehrere, unterschiedlich starke Wurzeln ausgebildet (Buchen, Birken, Hainbuchen, Lärchen, Linden).
- Horizontalwurzelsystem: Die Hauptwurzel ist nur bei jungen Bäumen vorhanden. Es gibt mehrere waagrecht (plagiotrop) wachsende Seitenwurzeln (Pappeln).
- Eine Mischform ist das Senkerwurzelsystem, wo aus starken Horizontalwurzeln Senkerwurzeln senkrecht in den Boden wachsen (Esche, Fichte in der Altersphase).
Je nachdem, ob die Wurzeln tief in den Erdboden vorstoßen oder sich knapp unter der Erdoberfläche eher vertikal ausdehnen, unterscheidet man:
- Tiefwurzler, die ihre Pfahlwurzel in Richtung Grundwasser treiben (neben den obengenannten Bäumen beispielsweise Königskerze oder Rettich)
- Flachwurzler, die mehr an die Aufnahme des im Boden versickernden Oberflächenwassers angepasst sind.
Das Wurzelsystem kann bei den einzelnen Pflanzen – je nach den Standortbedingungen – ganz unterschiedlich ausgeprägt sein.
Eine weitere Unterscheidung ist die zwischen – oft verholzter – Grobwurzel und der Feinwurzel:
- Die Grobwurzeln bilden das Wurzelgerüst, geben der Pflanze Halt und definieren den durchwurzelten Bodenbereich.
- Die dünnen Feinwurzeln besitzen oft nur eine kurze Lebensdauer und sind für die Wasser- und Nährstoffaufnahme zuständig. Die Obergrenze für Feinwurzeln liegt je nach Definition und Pflanze zwischen 0,8 Millimetern Durchmesser für Ackerpflanzen[1] und zwei Millimetern für Bäume.[2] Die Feinwurzeln sind auch die physiologisch aktivsten Wurzeln.
Primärer Aufbau
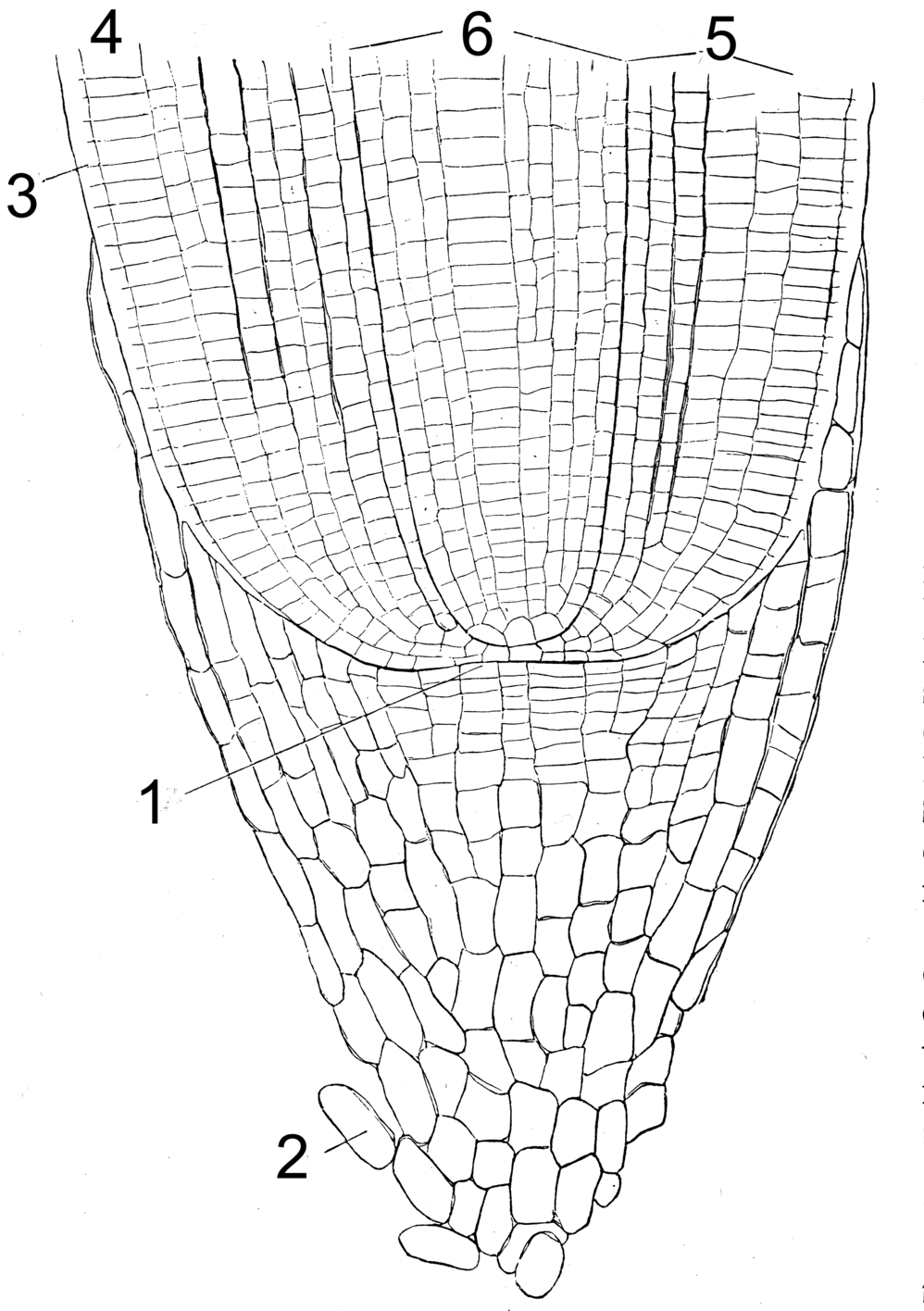
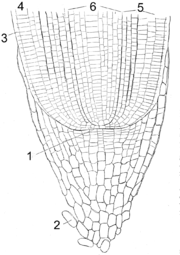 Wurzelspitze. 1 Meristem, 2 Wurzelhaube, 3 Rhizodermis, 4 Dermatogen (bildet Exodermis), 5 Periblem (bildet Rinde), 6 Plerom (bildet Zentralzylinder)
Wurzelspitze. 1 Meristem, 2 Wurzelhaube, 3 Rhizodermis, 4 Dermatogen (bildet Exodermis), 5 Periblem (bildet Rinde), 6 Plerom (bildet Zentralzylinder)Das Wachstum der Wurzel erfolgt mittels Wurzelspitzenmeristem. Bei den Farnen ist dies eine tetragonale, vierschneidige Scheitelzelle, die nach allen vier Richtungen Zellen abgliedert. Bei den anderen höheren Pflanzen wird das Meristem von einer Gruppe von Initialzellen gebildet.
Das Meristem ist durch die Wurzelhaube (Kalyptra) geschützt. Die Wurzelhaube besteht aus parenchymatischen Zellen, deren Mittellamellen verschleimen und so das Vordringen der Wurzel in den Boden erleichtern. Die Zellen werden laufend abgestoßen und vom Vegetationspunkt her neu gebildet. Der Schleim verändert durch das Verkleben mit Tonteilchen auch die Bodenstruktur. Die negative Ladung der Schleime spielen auch eine Rolle bei der Absorption von Nährstoffen und potenziell schädlichen Ionen (Aluminium, Cadmium).[3] Der Schleim wird zwar nur von der Wurzelhaube gebildet, bleibt beim Weiterwachsen der Wurzel aber auch an der Exodermis und den Wurzelhaaren kleben. Dadurch bildet sich bei Gräsern um die jungen Wurzelteile oft eine Wurzelscheide aus verklebten Bodenteilchen.
Vom Meristem nach rückwärts verläuft die Bildung und Ausdifferenzierung der Gewebe. Die nur wenige Millimeter lange Zellstreckungszone folgt auf die meristematische Zone. Hier erlangen die Zellen ihre endgültige Größe. In der folgenden Wurzelhaarzone erfolgt auch die innere Ausdifferenzierung der Gewebe.
Rhizodermis
Die Rhizodermis ist das einschichtige Abschlussgewebe. Sie besitzt im Gegensatz zur Epidermis des Sprosses keine Cuticula und keine Stomata. In der Wurzelhaarzone bilden die Rhizodermiszellen papillen- bis schlauchartige Ausstülpungen, die Wurzelhaare. Sie dienen der Oberflächenvergrößerung und damit der effektiveren Aufnahme von Wasser und Nährstoffen. Die Zellwände sind dünn und verschleimt (zur leichteren Durchdringung des Bodens), die Zellen besitzen große Vakuolen, Kern und Plasma befinden sich oft an der Haarspitze. Es können alle Zellen der Rhizodermis zu Wurzelhaaren auswachsen, oder nur bestimmte Haarbildner (Trichoblasten). Wurzelhaare und die anderen Rhizodermiszellen haben nur eine Lebensdauer von einigen Tagen und sterben danach ab.
Exodermis
Noch vor dem Absterben der Rhizodermis bildet sich aus dem äußeren Rindenparenchym ein sekundäres Abschlussgewebe, die Exodermis (auch Hypodermis genannt). Dieses ist ein Gewebe aus einer oder mehreren Zelllagen, dessen Zellen ohne Interzellularen miteinander verbunden sind und Suberin Akkrustierungen besitzen. In manchen Wurzeln finden sich außer diesen verkorkten Zellen auch sogenannte Durchlasszellen ohne Suberinakkrustierungen (sogenannte Kurzzellenexodermis). Die Exodermis bildet den äußersten Abschluss der primären Wurzel und verhindert den Wasser- und Nährstoffverlust aus der Wurzel.
Wurzelrinde
Die Wurzelrinde ist ein in der Regel farbloses Parenchym mit großen, schizogenen Interzellularen. Die Rinde ist ein Speichergewebe und dient auch dem Stoffaustausch zwischen der Rhizodermis und dem Xylem. Die Wurzelrinde ist in der Regel von Mykorrhiza-Pilzen besiedelt. Bei Wurzeln ohne sekundäres Dickenwachstum bildet die Rinde auch Festigungsgewebe (Sklerenchym und Kollenchym) aus.
Endodermis
Die Endodermis ist die innerste Schicht der Rinde und umschließt den Zentralzylinder lückenlos als einschichtige Lage lebender Zellen. Die Zellwände sind besonders entwickelt und verhindern den apoplastischen Einstrom von Wasser in den Zentralzylinder. Somit kontrolliert die Endodermis den Durchtritt von Wasser und Nährsalzen. Zugleich ist die Endodermis eine Barriere für die Mykorrhiza-Pilze. Es werden drei Zustände der Endodermis unterschieden:
Primäre Endodermis
Die Primäre Endodermis ist durch den Casparischen Streifen gekennzeichnet, eine streifenartige Zone der Radial- und Horizontalwände, in die ein suberinartiges Polymer („Endodermin“) und Lignin ein- und aufgelagert ist, wodurch die Zellwand wasserundurchlässig wird. Bei Angiospermen mit sekundärem Dickenwachstum ist dies der Endzustand der Endodermis, da sie beim Dickenwachstum abgestoßen wird.
Sekundäre Endodermis
In der Sekundären Endodermis sind alle Wände mit suberinähnlicher Substanz versehen. Einige unverkorkte Zellen, die „Durchlasszellen“, ermöglichen den Stoffaustausch zwischen Rinde und Zentralzylinder. Bei Koniferen ist dies die Endstufe der Entwicklung, ansonsten die Übergangsstufe zur Tertiären Endodermis.
Tertiäre Endodermis
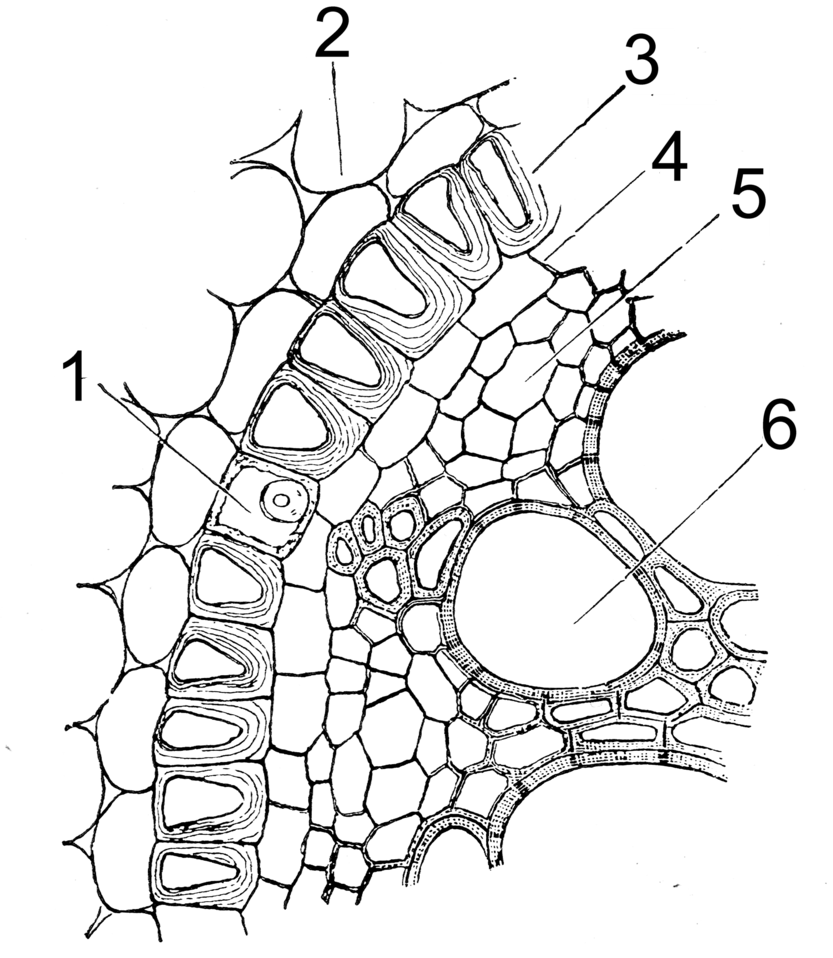
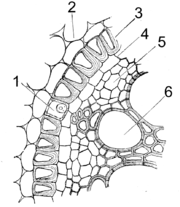 Tertiäre Endodermis (Iris florentina). 1 Durchlasszelle, 2 Rindenparenchym, 3 Endodermis, 4 Perizykel, 5 Phloem, 6 Xylem
Tertiäre Endodermis (Iris florentina). 1 Durchlasszelle, 2 Rindenparenchym, 3 Endodermis, 4 Perizykel, 5 Phloem, 6 XylemAuf die Suberinlamellen werden dicke Zellulosewände aufgelagert, wobei der Protoplast am Leben bleibt. Bei allseitiger Verdickung spricht man von O-Endodermen, bleibt die tangentiale Außenwand unverdickt, von C- oder U-Endodermen. Diese Zelluloseschichten können auch verholzen. Es gibt weiterhin die Duchlasszellen, die sich typischer Weise an das Xylem anschließen. Die Tertiäre Endodermis ist der typische Endzustand bei den Einkeimblättrigen Pflanzen.
Perizykel
Der Perizykel, auch Perikambium genannt, schließt an die Endodermis an und bildet den äußersten Teil des Zentralzylinders. Er ist eine meist einschichtige Scheide aus lückenlos aneindergrenzenden Zellen, die in Längsrichtung gestreckt sind. Die Zellen sind restmeristematisch oder parenchymatisch. Bei Einkeimblättrigen und Koniferen ist der Perizykel oft auch mehrschichtig, bei Ersteren oft sklerenchymatisch. Vom Perizykel gehen die Periderm- und die Seitenwurzelbildung aus, er ist auch wesentlich am Sekundären Dickenwachstum beteiligt.
Leitbündel
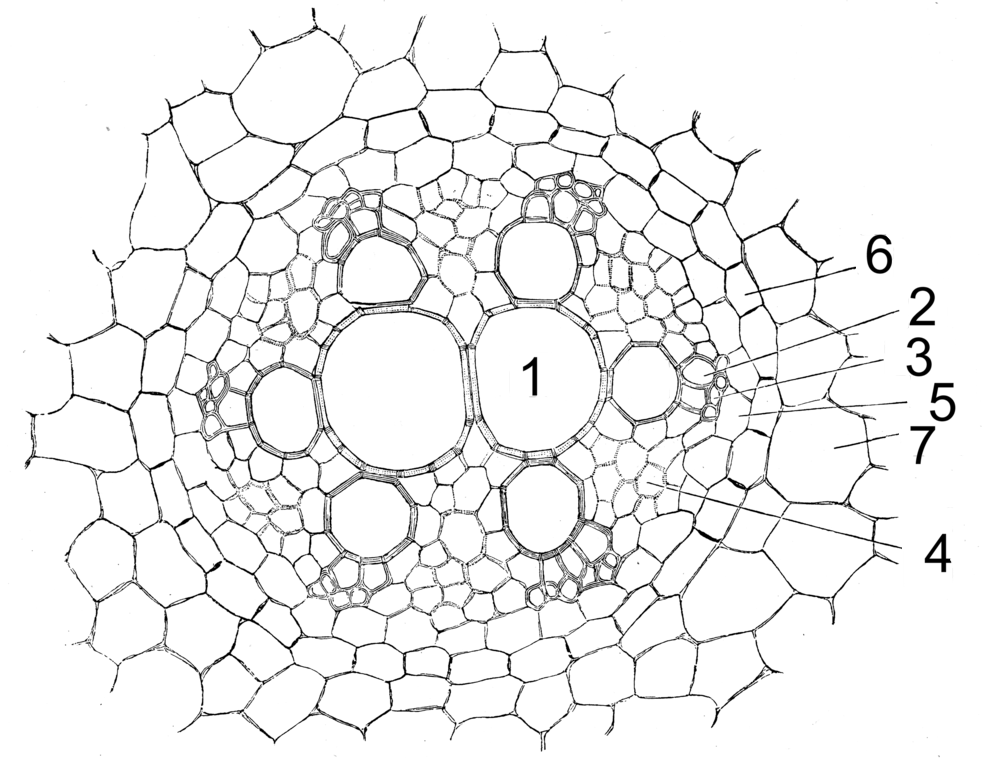
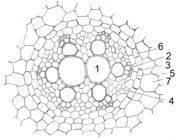 Leitbündel von Allium cepa. 1-3 Xylem (1 Treppengefäß, 2 Schraubentracheiden, 3 Ringtracheiden), 4 Phloem, 5 Perizykel, 6 Endodermis, 7 primäre Rinde
Leitbündel von Allium cepa. 1-3 Xylem (1 Treppengefäß, 2 Schraubentracheiden, 3 Ringtracheiden), 4 Phloem, 5 Perizykel, 6 Endodermis, 7 primäre RindeDer Zentralzylinder ist in der Regel als Aktinostele organisiert, die Teile sind radial angeordnet. In der Mitte befindet sich in der Regel Xylem. Dieses reicht in zwei bis vielen Leisten bis an den Perizykel, dazwischen befindet sich das Phloem. Einkeimblättrige besitzen meist viele Xylemstränge, man spricht von polyarchen Wurzeln. Dikotyle und Koniferen besitzen meist wenige Xylemstränge (oligoarche Wurzeln), man unterscheidet dabei wieder zwei-, drei-, vierstrahlige usw. Wurzeln (di-, tri-, tetrarch). Phloem und Xylem differenzieren sich von außen nach innen (zentripetal), im Gegensatz zum Spross. Die unterschiedliche Anordnung von Xylem und Phloem in Spross und Wurzel erfordert im Übergangsbereich des Hypokotyls eine Drehung und Umorientierung der Leitbündel.
Wurzelzentrum
Im Wurzelzentrum befinden sich meist weitlumige Leitgefäße des Xylems. Es können sich hier aber auch Speicherparenchym oder Sklerenchymstränge befinden.
Seitenwurzeln
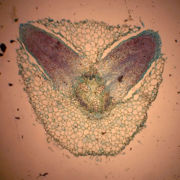 Seitenwurzelbildung bei derErbse
Seitenwurzelbildung bei derErbseDie Seitenwurzeln entstehen endogen, also im Inneren der Wurzel, im Gegensatz zu den Seitensprossen der Sprossachse, die exogen gebildet werden. Meist entstehen sie vor den Xylemprimanen aus dem Perizykel. Daher stehen die Seitenwurzeln in Reihen (Rhizostichen), deren Anzahl mit der Anzahl der Xylemstrahlen im Zentralzylinder übereinstimmt.
Die Zellen des Perizykels werden wieder teilungsfähig und werden zum Perikambium. Sie bilden einen konusförmigen Gewebehöcker, der durch die Rinde hindurchwächst und diese dabei aufreißt. Junge Seitenwurzeln wachsen zunächst senkrecht von der Hauptwurzel weg. Sie reagieren erst später positiv geotrop. Die Endodermis der Hauptwurzel wächst zunächst mit und verbindet sich mit der Endodermis der Seitenwurzel. Die Leitelemente der Seitenwurzel werden gleichfalls an diejenigen der Hauptwurzel angeschlossen.
Sekundäres Dickenwachstum
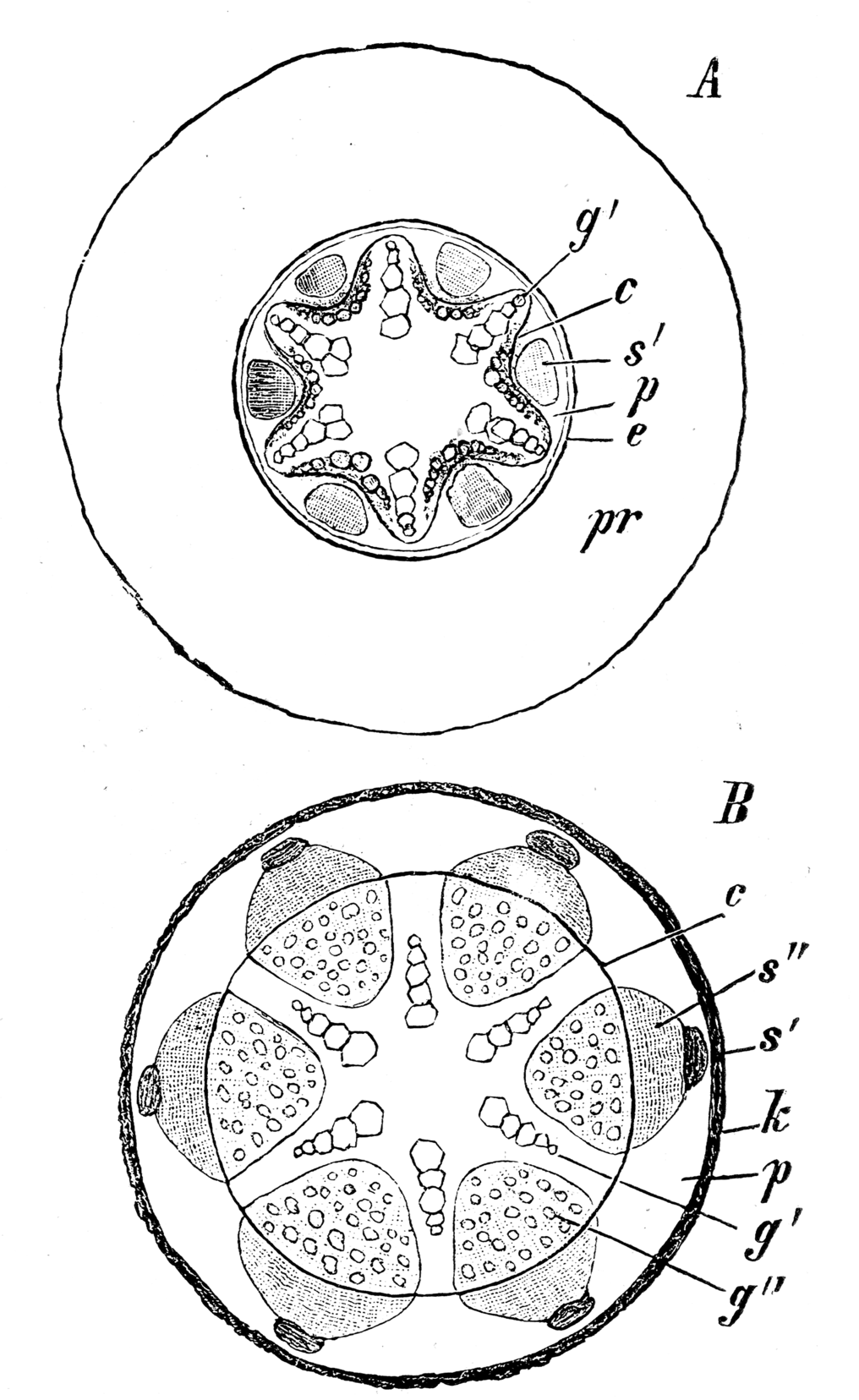
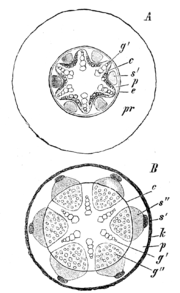 Sekundäres Dickenwachstum. A beginnend, B fortgeschritten. pr primäre Rinde, e Endodermis, c Cambiumring, g' primäres Xylem, s' primäres Phloem, p Perizykel, g" secundäres Holz, s" sekundärer Bast, k Periderm
Sekundäres Dickenwachstum. A beginnend, B fortgeschritten. pr primäre Rinde, e Endodermis, c Cambiumring, g' primäres Xylem, s' primäres Phloem, p Perizykel, g" secundäres Holz, s" sekundärer Bast, k PeridermBei Koniferen und Dikotylen wachsen die Wurzeln wie auch die Sprossachsen sekundär in die Dicke. Das Dickenwachstum setzt gleichzeitig mit dem der Sprossachse ein.
Das Parenchym zwischen Xylem und Phloemsträngen wird meristematisch und bildet das neu entstehende Kambium. Die über dem Xylem liegenden Teile des Perizykels werden gleichfalls meristematisch. Dadurch entsteht ein geschlossener, im Querschnitt sternförmiger Kambiummantel.
Dieses Kambium gliedert nun nach innen Xylem- und nach außen Phloemelemente ab. Der zunächst stärkere Xylemzuwachs führt zu einer Abrundung des Kambiums, das schließlich zylinderförmig wird. Während des Wachstums werden ähnlich wie in der Sprossachse primäre Markstrahlen und sekundäre Strahlen gebildet. Die ersten Strahlen entstehen über den primären Xylempolen.
Exodermis und Rinde folgen dem Sekundären Dickenwachstum nicht. Das Gewebe reißt auf, die Zellen sterben ab. Die Endodermis macht am Beginn das Dickenwachstum durch Dilatation noch mit, reißt später aber ebenfalls. Als sekundäres Abschlussgewebe wird ein Periderm angelegt, das durch den Perizykel, der jetzt vollends zum Perikambium geworden ist, gebildet wird. Bei Holzgewächsen bildet sich eine Borke ähnlich der der Sprossachse.
Mehrere Jahre alte Wurzeln unterscheiden sich kaum mehr von Sprossachsen. Unterschiede finden sich nur mehr in der radialen Anordnung der primären Leitelemente in der Mitte der Wurzel. Das Holz der Wurzel ist meist weitlumiger und ähnelt daher dem Frühholz des Stammes. Aus diesem Grund sind auch die Jahresringe weniger stark ausgeprägt.
Funktionen
Die Hauptfunktionen der Wurzel sind die Wasser- und Mineralstoffaufnahme aus dem Boden sowie die Befestigung der Pflanze im Boden. Andere Funktionen gehen meist mit deutlichen Umgestaltungen parallel und werden weiter unten besprochen.
Wasseraufnahme
Die Wasseraufnahme erfolgt über die Wurzelhaare und die feinen Seitenwurzeln. Ältere Wurzeln sind verkorkt und dienen nur mehr der Wasserleitung.
Wurzeln entwickeln meist ein negatives Wasserpotential von nur einigen Zehntel Megapascal (MPa). Hygrophyten können meist maximal −1 MPa erreichen, Mesophyten −4 MPa und Xerophyten −6 MPa. Waldbäume erreichen nicht mehr als −2 bis −4 MPa.
Das Wurzelsystem kann Wasser aus dem Boden nur so lange aufnehmen, als das Wasserpotential der Feinwurzeln niedriger ist als des umgebenden Bodens. Die Wasseraufnahme Wa während einer Zeiteinheit ist dabei proportional der Austauschfläche A (aktive Wurzelfläche) und dem Potentialunterschied ψ zwischen Wurzel und Boden. Sie ist umgekehrt proportional den Transferwiderständen r für Wasser im Boden (Nachleitwiderstand) und beim Übergang vom Boden in die Pflanze (Permeationswiderstand):[4]
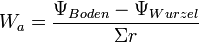
Das Wasser gelangt vom Boden über das Rindenparenchym bis an die Endodermis. Der Transport kann in diesem Bereich sowohl innerhalb der Zellen (symplastisch) oder auch im Zellzwischenraum (im Apoplasten) erfolgen. Durch den Casparischen Streifen wird in der Endodermis der apoplastische Transport blockiert. Das Wasser gelangt nur über den Symplasten weiter in den Zentralkörper und hier in die Xylem-Gefäße, von wo es durch Ferntransport in die Blätter gelangt.
Mineralstoffaufnahme
Pflanzen nehmen mit wenigen Ausnahmen die für das Wachstum benötigten Mineralstoffe über die Wurzeln auf. Zu den Ausnahmen zählen die Wasserpflanzen, die Wasser und Nährstoffe über die gesamte Pflanzenoberfläche aufnehmen. In manchen Bereichen spielt auch die Nährstoffaufnahme über die Blätter eine gewisse Rolle. Dafür siehe dort.
Die Entnahme aus dem Boden erfolgt durch drei Prozesse:
- Durch Absorption von Nährionen aus der Bodenlösung: diese Ionen befinden sich bereits frei in Lösung und sind sofort für die Pflanze verfügbar. Die Konzentrationen sind jedoch meist sehr gering: Nitrat häufig 5 bis 10 mM, Phosphat meist unter 4 µM.
- Durch Austauschabsorption von sorbierten Nährstoffionen: Diese Ionen sind relativ locker an Ton und Huminteilchen gebunden. Durch die Abgabe von Wasserstoffionen und Hydrogenkarbonat, den Dissoziationsprodukten des Atmungs-Kohlendioxids in Wasser, fördert die Pflanze den Ionenaustausch an diesen Teilchen. Dadurch gehen die Nährstoffionen in Lösung und können damit aufgenommen werden.
- Durch Mobilisierung von chemisch gebundenen Nährstoffvorräten: Dies geschieht durch Ausscheiden von organischen Säuren und von Chelat-Bildnern. Die Wasserstoffionen der dissoziierten Säuren lösen Nährstoffe aus Mineralen. Die Chelat-Bildner sind organische Säuren (z. B. Äpfelsäure, Citronensäure) und Phenole (z. B. Kaffeesäure), die besonders mit den wichtigen Mikronährstoffen wie Eisen Metallchelate bilden und sie so vor neuerlicher Festlegung schützen.
Die Nährionen gelangen mit dem Wasser zunächst in den apoplasmatischen Raum der Wurzel. Die Ionenaufnahme in das Cytoplasma erfolgt großteils im Rindenparenchym, da der apoplastische Transport am Casparischen Streifen der Endodermis endet. Da die Konzentration der Nährstoffe meist in der Pflanzenzelle höher ist als in der Bodenlösung, sind für die Aufnahme der Ionen in die Pflanzenzelle aktive Transportprozesse nötig. Diese werden durch den Aufbau eines Protonengradienten durch Membran-ATPasen ermöglicht. Die Ionen werden über spezifische Ionophore und Tunnelproteine aufgenommen. Durch diese Mechanismen erreicht die Pflanze ein Anreicherungsvermögen gegenüber der Bodenlösung und ein Wahlvermögen, d. h. sie kann bestimmte, in geringer Konzentration vorliegende Ionen gegenüber anderen, häufigeren Ionen, bevorzugen.
Die Ionen werden symplastisch (von Zelle zu Zelle) weitertransportiert. In die Leitgefäße des Xylems, die Tracheen und Tracheiden, werden die Ionen passiv aufgrund des Konzentrationsgefälles abgegeben. Zusätzlich werden sie von den angrenzenden Parenchymzellen auch aktiv in die Gefäße abgeschieden. Von hier an erfolgt dann der Ferntransport.
Befestigung
Eine weitere Grundfunktion der Wurzeln ist die Verankerung der Pflanze im Boden. Der Zugbeanspruchung der Wurzel entsprechend, sind die festen Elemente (Xylem) in der Mitte konzentriert, wodurch sich ein besonders zugfester, der Kabelbauweise entsprechender, anatomischer Bau ergibt.
Metamorphosen
Pflanzen begegnen den verschiedenen ökologischen Bedingungen ihrer Umwelt mit entsprechenden Umbildungen und Abwandlungen des Grundaufbaus ihrer Organe (Metamorphosen). Die Metamorphosen der Wurzel werden durch die speziellen Aufgaben, welche die Wurzel wahrnehmen muss, bestimmt.
 Speicherwurzeln des Scharbockskrauts
Speicherwurzeln des ScharbockskrautsSpeichermetamorphosen
Viele Pflanzen speichern in ihren Wurzeln Reservestoffe, besonders Pflanzen mit ausgeprägter Pfahlwurzel.
Besondere Speicherorgane sind Wurzelknollen. Sie treten bei vielen Orchideen, z. B. beim Helmknabenkraut (Orchis militaris), bei den Dahlien (Dahlia) oder beim Scharbockskraut (Ranunculus ficaria) auf.
Bei Rüben gehört zumindest meistens ein Teil der Wurzelregion an, der andere der Sprossachse (Hypokotyl). Reine Wurzelrüben besitzen z. B. die Karotte und die Zuckerrübe.
Wurzeln können zu sukkulenten, also wasserspeichernden Organen umgewandelt werden. Diese Wurzelsukkulenz kann z. B. bei Grünlilien schwach ausgeprägt gefunden werden.
Befestigung
 Mangroven bilden Stelzwurzeln
Mangroven bilden StelzwurzelnKletterpflanzen und Epiphyten nutzen Wurzeln zur Anheftung an den Untergrund. Dies kann einerseits wie bei der Vanille durch Wurzelranken geschehen, andererseits wie beim Efeu durch Haftwurzeln.
Schnell wachsende, häufig schlecht verankerte Bäume bilden Brettwurzeln durch exzessives sekundäres Dickenwachstum der Oberseite von unmittelbar unter der Erdoberfläche horizontal wachsenden Wurzeln. Dies tritt häufig bei Bäumen des tropischen Regenwaldes auf.
Maispflanzen bilden Stützwurzeln an den untersten Knoten des Sprosses, da sie über kein sekundäres Dickenwachstum verfügen. Die Stützwurzeln verhindern so ein Umfallen, da der untere Sprossteil alleine zu schwach wäre.
Viele Mangrovenbäume bilden Stelzwurzeln, welche sie über das mittlere Hochwasserniveau heben.
Atemwurzeln
 Atemwurzeln einer Sumpfzypresse
Atemwurzeln einer SumpfzypresseIn Sumpf oder Schlick wachsende Pflanzen bilden negativ gravitrop wachsende Atemwurzeln, Pneumatophore, die das Wurzelsystem über das Interzellularsystem des Rindengewebes mit Sauerstoff versorgen. Beispiele sind wiederum die Mangrovenbäume. Atemknie werden aber auch von Zypressengewächsen gebildet.
Luftwurzeln
Einige, vor allem epiphytische Pflanzen haben die Möglichkeit auch über die Luftwurzeln Wasser aufzunehmen, da sie die Wasserreservoirs des Bodens nicht erreichen. Die Luftwurzeln besitzen hierfür ein spezielles Gewebe, das Velamen radicum. Dieses Velamen liegt außerhalb der Exodermis und enthält viele frühzeitig abgestorbene Zellen und daher viele Lufträume. Diese Zellen saugen Regenwasser kapillar auf und leiten es über Durchlasszellen der Exodermis in den Wurzelkörper.
Manche Luftwurzeln besitzen in den Rindenzellen photosynthetisch aktive Chloroplasten. Bei einigen epiphytischen Orchideen wurden die Blätter reduziert und die bandförmig verbreiterten Luftwurzeln haben die Aufgabe zur Photosynthese übernommen.
Haustorien
 Zottiger Klappertopf (links) und Kleiner Klappertopf (rechts) sind Hemiparasiten aufgrund ihrer Wurzelhaustorien.
Zottiger Klappertopf (links) und Kleiner Klappertopf (rechts) sind Hemiparasiten aufgrund ihrer Wurzelhaustorien.Pflanzliche Parasiten zapfen ihre Wirtspflanzen über Wurzel-Haustorien an. Die Weißbeerige Mistel bildet im Bast des Wirtsbaums Rindenwurzeln, von denen aus sie Senker in das Splintholz des Wirtes treibt, wo sie über Kurztracheen direkten Anschluss an das Xylem des Wirtes errichtet. Die Schuppenwurz (Lathraea) zapft das Xylem von Baumwurzeln an, während Sommerwurz-Arten (Orobanche) das Phloem der Wirtswurzeln anzapfen und sogar die über der Anzapfungsstelle liegenden Wurzelteile zum Absterben bringen können. Weitere Schmarotzer mit Wurzelhaustorien sind Augentrost, Klappertopf, Wachtelweizen und Läusekraut.
Wurzeldornen
Bei bestimmten Palmen sind die Enden einiger Luftwurzeln zu Wurzeldornen als Schutz der Stammbasis umgebildet.
Zugwurzeln
Zugwurzeln ziehen Erdsprosse – also Knollen, Zwiebeln oder Rhizome – durch Wurzelkontraktionen tiefer in den Boden. Die Rindenzellen dieser Wurzeln sind längs (axial) gestreckt, die Fasern der Zellwände verlaufen ebenfalls längs. Bei einer Erhöhung des Turgors verkürzen und verdicken sich daher die Zellen. Durch die Verdickung werden die Wurzeln fixiert, sodass durch die Verkürzung die Erdsprosse nach unten gezogen werden. Diese Funktion tritt bei vielen Geophyten auf.
Symbiosen
Oft sind die Feinwurzeln in Form einer Symbiose mit Pilzen (Mykorrhiza) oder Bakterien kombiniert, um die Nährstoffaufnahme zu verbessern.
Bei der Symbiose mit Stickstoff-fixierenden Bakterien (z. B. Aktinomyzeten) kommt es zur Bildung von Wurzelknöllchen, lokalen Wucherungen des Rindengewebes In diesen Wucherungen bestehen vergrößerte, polyploide Parenchymzellen, in denen die Symbionten als Knöllchenbakterien, bzw. Bacteroide, in speziellen Vakuolen leben. Diese Symbiose ermöglicht es beispielsweise den Erlen, an Stellen mit wenig Nährstoffen zu leben.
Die Symbiose mit Hyphengeflecht von Bodenpilzen (Mykorrhiza) ist dagegen noch weiter verbreitet. Da die Hyphen des Pilzes eine enorme Absorptionsfähigkeit haben, bilden Wurzeln mit Hyphenkontakt keine Wurzelhaare aus. Einigen Pflanzen, wie dem Fichtenspargel ermöglichen Mykorrhiza auf eigene Photosynthese zu verzichten, womit wieder ein Holoparasitismus erreicht ist.
Fußnoten
Literatur
- W. Braune, A. Leman, H. Taubert: Pflanzenanatomisches Praktikum I. Zur Einführung in die Anatomie der Vegetationsorgane der Samenpflanzen. 6. Auflage, Gustav Fischer, Jena 1991, S. 221–242. ISBN 3-334-60352-0
- Walter Larcher: Ökophysiologie der Pflanzen. 5. Auflage, Ulmer, Stuttgart 1994, ISBN 3-8252-8074-8
- Horst Marschner: Mineral Nutrition of Higher Plants. 2. Auflage, Academic Press, London etc. 1995, ISBN 0-12-473543-6
- Margaret E. McCully: Roots in Soil: Unearthing the Complexities of Roots and Their Rhizospheres. Annual Review in Plant Physiology and Plant Molecular Biology 1999, 50: S. 695–718.
- Peter Schütt, Hans Joachim Schuck, Bernd Stimm: Lexikon der Baum- und Straucharten. ecomed, Landsberg/Lech 1992 (Nachdruck 2002, ISBN 3-933203-53-8)
- P. Sitte, E. W. Weiler, J. W. Kadereit, A. Bresinsky, C. Körner: Strasburger. Lehrbuch der Botanik für Hochschulen. 35. Auflage, Spektrum Akademischer Verlag, Heidelberg 2002. ISBN 3-8274-1010-X
Weblinks






Wikimedia Foundation.