- Wasserstress
-
Als Trockenstress oder auch Wasserstress bezeichnet man einen Stress, der durch Wassermangel auf lebende Organismen und insbesondere auf Pflanzen hervorgerufen wird.
Inhaltsverzeichnis
Übersicht
Trockenstress ist relevant vor allem in ariden Klimaten (Wüstengürtel, Subtropen), aber auch in kalten Gebieten (Tundra, alpine Regionen).
Die wichtigen Einflussgrößen, die in ariden Gebieten zu Trockenstress führen, sind: geringe Niederschläge, hohe Temperaturen, geringe Luftfeuchtigkeit und ungünstige Bodeneigenschaften (Wasserrückhaltevermögen). Sie führen dazu, dass die Transpiration der Pflanze größer ist als die Wasseraufnahme. In kalten Gebieten besteht die Gefahr, dass durch das Gefrieren des Bodenwassers die Wassernachlieferung ungenügend ist, was somit zu Trockenstress führt.
Wassermangel und Trockenstress sind die bedeutendsten Limitationen für die landwirtschaftliche Produktion. Die Verluste durch Trockenstress übertreffen bei weitem die, die auf andere abiotische oder auch biotische Faktoren zurückzuführen sind. Die meisten Entwicklungsländer befinden sich in ariden Gebieten und sind deshalb besonders betroffen.
Anpassungsstrategien bei Pflanzen
Hydrostabile Pflanzen unterscheiden sich von hydrolabilen Pflanzen in ihrer Fähigkeit, sich Trockenstress anzupassen. Sie haben ihre Anatomie und ihren Stoffwechsel so weit an das Problem des Wassermangels angepasst, dass sie auf Trockenheit reagieren können und den Wassergehalt in ihrem Gewebe für einen gewissen Zeitraum aufrecht erhalten. Diese hydrostabilen Pflanzen kann man anhand ihrer Strategie der Anpassung wiederum in wassersparende und wasserverbrauchende Pflanzen unterteilen:
Wassersparende Pflanzen vermeiden die Wasseraufnahme durch das Wurzelwerk und lassen es zunächst im Boden, sie teilen es sich regelrecht ein um es länger nutzen zu können. So kann mit weniger Wasserverbrauch eine längere Trockenphase überwunden werden.
Wasserverbrauchende Pflanzen verfolgen die entgegengesetzte Strategie. Bei auftretender Trockenheit entziehen sie dem Boden sämtliches Wasser und speichern es, sodass auf weitere vergebliche Wasseraufnahme während der Trockenphase verzichtet werden kann. Ein Beispiel für diesen Anpassungstyp ist Prosopis spec., der Mesquitebaum. Sein Wurzelwerk reicht in bis zu 100 Metern Tiefe. Sobald das im Boden zur Verfügung stehende Wasser weniger wird, entzieht er sämtlichen Bodenschichten das Restwasser und speichert es in seinem Stamm. Dadurch sichert er sich nicht nur die Wasserversorgung während der Trockenphase, er verschafft sich auch einen Konkurrenz-Vorteil naheliegenden Pflanzen gegenüber, die aus diesem Boden somit kein Wasser mehr aufnehmen können.
Kurzfristige Anpassungen
Bei Trockenstress wird durch die Verminderung des Turgordruckes infolge von Wasserverlust Abscisinsäure (ABA, abscisic acid) in der Wurzel und auch im Blatt gebildet. Abscisinsäure beeinflusst an den Zielzellen und Zielgeweben physiologische und biochemische Reaktionen in einer Weise, welche dem Organismus das Überleben unter Trockenstress erleichtert oder überhaupt erst ermöglicht. Eine der wichtigsten Aufgaben von Abscisinsäure ist die Induktion des Stomataschlusses. Die Regulation der Stomataöffnung geschieht bereits nach einigen Minuten. ABA kann somit auf zwei Arten eingesetzt werden:
1. Als Frühwarnsystem. Ein Wasserdefizit im Boden löst die vermehrte Produktion von ABA in den Wurzeln aus. Durch Assimilationsströme verteilt es sich in der Pflanze, erreicht letztlich auch die Blätter mit ihren Stomata, welche sich daraufhin schließen.
2. Als direkte Reaktion auf Wasserverlust des Blattes. Der sinkende Turgordruck in den Zellen von Blättern veranlasst die lokale Ausschüttung von ABA, was das Schließen der Stomata bewirkt.
Somit erlaubt ABA eine präzise und reversible Steuerung des Wasserhaushaltes einer Pflanze oder sogar eines einzelnen Blattes. Unter Lichtbedingungen führt der Schluss der Stomata zu einer Abnahme der internen CO2-Konzentration und damit zu einer starken Hemmung der Photosynthese. Dies führt in C3-Pflanzen zu einer gesteigerten Aktivität der Photorespiration und damit zur Bildung von reaktiven Sauerstoffverbindungen (H2O2).
Das Schließen der Stomata im Detail
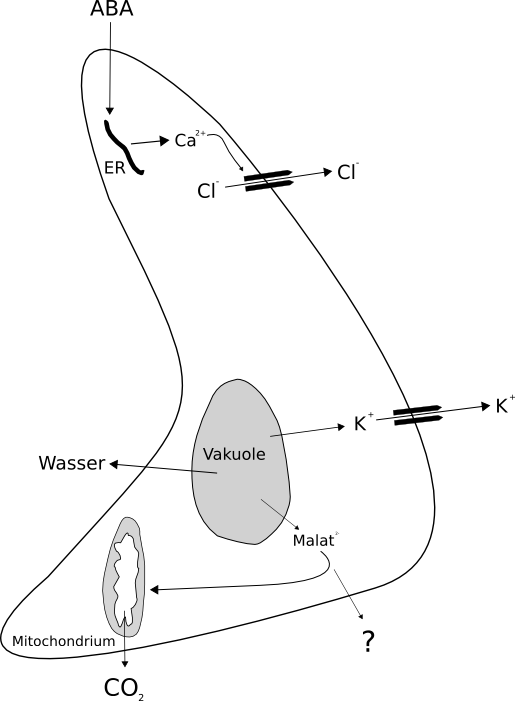
 Schematische Darstellung der Wirkung auf Schließzellen bei Wasserstress
Schematische Darstellung der Wirkung auf Schließzellen bei WasserstressBei Trockenstress wird Abscisinsäure in der Wurzel und auch im Blatt gebildet. Sie wird durch den Transpirationsstrom an die Schließzellen herangeführt. In diesen Zellen induziert sie die Freisetzung von Calcium-Ionen. Der gesteigerte Calciumspiegel bewirkt eine Hemmung der Protonen-ATPase, wodurch es zu einem Absinken des transmembranen H+-Gradienten kommt, was zu einer Depolarisation führt. Außerdem binden Calcium-Ionen an Chloridkanäle. Dadurch strömen Chloridionen passiv aus der Zelle aus, was eine weitere Depolarisation zur Folge hat. Die exakten Einzelschritte sind allerdings noch Gegenstand der Forschung.
Diese Depolarisation hat zwei Konsequenzen. Zum einen schließen sich die Kaliumkanäle, die nur bei Hyperpolarisation geöffnet sind. Außerdem sind die nach außen gerichteten Kaliumkanäle besonders aktiv, was zu einem starken Ausstrom von Kaliumionen führt. Die Chlorid- und Malatanionen strömen daran gekoppelt nach, wodurch die Zelle osmotisch Wasser verliert und das Stoma sich schließt. Die Ionen werden in benachbarten Zellen gespeichert.
Die Regulation der Stomataöffnung geschieht bereits nach einigen Minuten. Somit erlaubt sie eine präzise und reversible Steuerung des Wasserhaushaltes einer Pflanze oder sogar eines einzelnen Blattes. Die molekularen Wirkungsweisen von Abscisinsäure sind noch nicht detailliert bekannt. Jedoch wurden in welkenden Tomaten-Mutanten nur etwa 10% der üblichen Abscisinsäure-Konzentration gefunden. Nach exogener Zugabe funktionierten die Stomata jedoch wieder korrekt.
Langfristige Anpassungen
Durch geringfügigen Wassermangel über einen längeren Zeitraum wird das proportionale Wachstum von Wurzel und Spross beeinflusst. Steht einer Pflanze weniger Wasser zur Verfügung, wächst der Spross langsamer oder stoppt ganz sein Wachstum. Die eigentlich dafür verwendeten Assimilate werden nun konzentriert der Wurzel zur Verfügung gestellt, sie wächst weiter und weiter und kann somit in tieferen Bereichen neues Wasser erschließen, während die oberen Bodenschichten langsam austrocknen. Dadurch zeichnen sich Pflanzen in feuchten Böden durch ein flaches Wurzelsystem, in trockenen Böden durch ein sehr tiefes Wurzelsystem aus. Das ist allerdings nur bei vegetativen Pflanzen der Fall. Reproduzierende Pflanzen speichern die übrigen Assimilate eher in Früchten und hemmen das Wachstum der gesamten Pflanze, somit auch der Wurzel. Außerdem bewirkt ABA eine Erhöhung der hydraulischen Wasserleitfähigkeit der Wurzel.
Laubfall
Bei dieser Strategie perennierender Pflanzen wird Wasserverlust minimiert, indem während Perioden mit schlechter Wasserverfügbarkeit (Dürre oder Winter) keine Transpirationsfläche vorhanden ist. Der Laubfall kann bei gewissen Pflanzen (aride Zone) nur partiell sein oder komplett wie bei vielen Pflanzen der gemäßigten Zone (Winterdormanz). Diese bilden Reservespeicher in Wurzeln oder in oberirdischen Pflanzenorganen, welche ein schnelles Austreiben bei besseren Bedingungen erlauben. Ein Beispiel für laubabwerfende Pflanzen aus ariden Zonen ist der Affenbrotbaum. Er treibt vollkommen neu erst nach dem nächsten Regen wieder aus. Der gesamte Blattverlust wird Blattabscission genannt.
Trockenstressvermeidung
Dürremeidende Pflanzen schließen ihren Lebenszyklus vor einer bevorstehenden Dürre ab. Die meiste Zeit des Jahres überdauert die Pflanze in Form von Speicherorganen unterhalb der Erde (Kryptophyten) oder in Form von Samen, die erst bei dem nächsten Regen keimen. Dieses Phänomen kann besonders in Wüsten und Steppen beobachtet werden. Nach einem starken Regen keimen die im Boden befindlichen Samen, wachsen innerhalb weniger Wochen heran, blühen, bilden Früchte und Samen aus und sterben schon nach 6 Wochen wieder ab. Die Samen überdauern wieder im Boden bis zur nächsten Regenzeit.
Dürretoleranz
Manche mehrjährige Wüstenpflanzen haben ein extrem tiefes Wurzelwerk. Der Affenbrotbaum zum Beispiel bildet bis zu 80 m tiefe Pfahlwurzeln, mit denen er an Grundwasser herankommt. Andere haben ein extrem ausgeweitetes Wurzelwerk, dessen Wurzeln bei manchen Arten bis zu 30 m lang werden können. Daher stehen Schirmakazien sehr weit voneinander entfernt. Hier konkurriert das Wurzelwerk um den seltenen Niederschlag. Durch extreme Verkleinerung der Blattoberfläche oder Umwandlung zu Dornen und schmalen Wuchs wird die Oberfläche, die der direkten Sonneneinstrahlung ausgesetzt ist, verringert und somit die Transpirationsrate gesenkt. Optisch lässt sich dies am besten an einem großen, schmalen Kaktus erkennen. Photosynthese findet nicht länger in Blättern statt, sondern im Spross der Pflanze. Die Blätter können auch verdickt, gerollt oder behaart sein. Andere Pflanzen haben ihren Spross komplett zurückgebildet. Durch diesen gedrungenen, rosettenförmigen Wuchs wird die Luftzirkulation verringert und eine Art Mikroklima entsteht. Somit wird ebenfalls Verdunstung vermieden. Den Formen und Variationen sind keine Grenzen gesetzt.
Die Sukkulenten (z.B. Kakteen) sind besonders gut an Wasserknappheit angepasst. Sie haben in ihren Geweben große Wasserreserven und haben keine Blätter. Sie betreiben Photosynthese mit dem Stamm und optimieren somit das Verhältnis des Volumens zur Transpirationsfläche. Das Wasser wird bei den meisten Sukkulenten in einem fächerartig gebildeten Stamm gespeichert, durch die Wasseraufnahme dehnt er sich aus. Verbraucht die Pflanze das Wasser, ziehen sich die Fächer wieder zusammen. Zwischen diesen Längsrippen sammelt sich morgendlicher Tau, läuft sie herab und wird direkt zu den Wurzeln geleitet, wo die Pflanze das Wasser aufnehmen kann.Zur Minimierung von Transpiration haben sie den CAM-Mechanismus.Siehe auch
Wikimedia Foundation.