- Brachiopoden
-
Armfüßer 
Onniella meeki
Systematik Unterreich: Vielzellige Tiere (Metazoa) Abteilung: Gewebetiere (Eumetazoa) Unterabteilung: Bilateria ohne Rang: Urmünder (Protostomia) Überstamm: Lophotrochozoen (Lophotrochozoa) Stamm: Armfüßer Wissenschaftlicher Name Brachiopoda Stämme - Linguliformea
- Craniiformea
- Rhynchonelliformea
Der Tierstamm der Armfüßer (Brachiopoda) besteht ausschließlich aus meereslebenden, bilateral-symmetrischen Tieren mit zweiklappigem Gehäuse. Sie ähneln äußerlich den Muscheln (Bivalvia), haben aber anstatt einer linken und rechten Schale (Klappe) eine obere und eine untere, wobei die bauchseitige Schale meist größer ist. Abgesehen davon besitzen die Brachiopoden an beiden Seiten des Mundes armförmige Tentakel. Die größten heute lebenden Arten erreichen Schalenbreiten bis sieben Zentimeter, die größten fossilen Schalen sind etwa 30 Zentimeter breit.
Inhaltsverzeichnis
Merkmale
Äußere Anatomie
Die Armfüßer erreichen im Regelfall Schalengrößen von maximal 5 Zentimetern, fossile Arten konnten allerdings auch bis zu 30 Zentimeter groß werden und vor allem bei den Atremata wie Lingula kann der Stiel ebenfalls bis zu 30 Zentimeter lang werden. Das auffälligste Merkmal der Armfüßer ist die zweiklappige Schale, die in eine Dorsal- und eine Ventralschale unterteilt wird. Obwohl sie äußerlich dadurch den Muscheln ähneln, kann man sie auf Anhieb an der Lage der Symmetrieachse erkennen. Diese geht bei ihnen senkrecht durch die Einzelklappe und teilt diese in jeweils zwei symmetrische Hälften, während sie bei Muscheln entlang des Schalenrandes verläuft. Vor allem bei den fossilen Vertretern können die Schalen sehr unterschiedlich geformt sein. So gibt es neben den in beide Richtungen gewölbten, bikonvexen Schalen, die die Mehrheit der Tiere besitzt, auch konkav-konvexe und abberante Formen. Bei einigen Arten ändert sich die Form im Laufe der Entwicklung von einer konvex-konkaven Form zu einer bikonvexen, in dem Fall spricht man von resupinanten Gehäusen.
Bei den meisten Arten sind beide Klappen stark verkalkt und mit einer organischen Schicht, dem Periostracum, bedeckt. Unterhalb dieser finden sich abwechselnd Schichten aus Calciumcarbonat (Calcit) und Proteinhäutchen. Bei einigen Arten liegt darunter noch eine Schicht aus prismenförmigen Calcitkristallen. Die Schalen können feine Kanälchen aufweisen, durch die Ausstülpungen des Mantels mit Sekretzellen, die so genannten Caeca, reichen. Eine solche Schale wird als „punctate“ Schale bezeichnet. Einige ursprüngliche Arten, wie etwa Lingula, haben dagegen eine wenig verkalkte und biegsame Schale aus Chitinlamellen und Calciumphosphat.
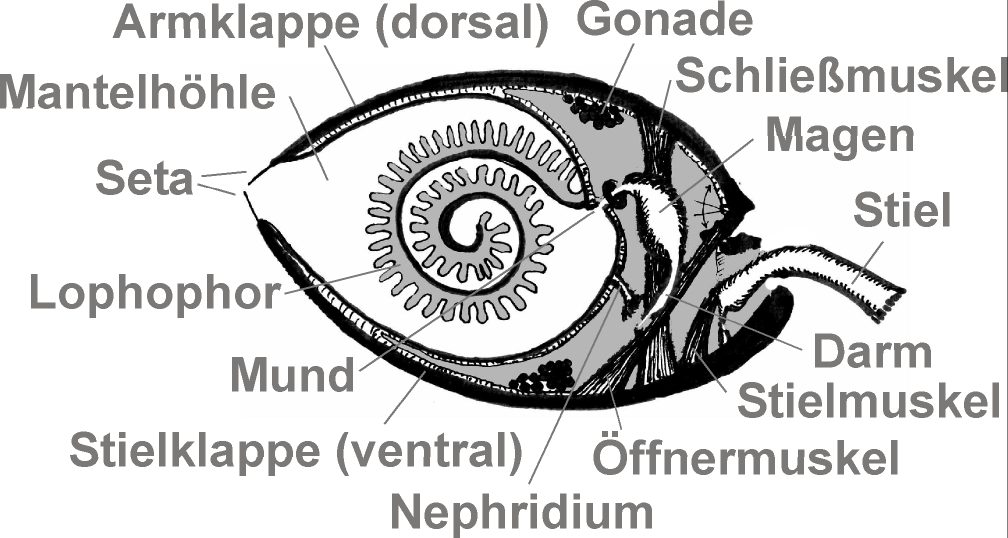
 Aufbau eines Vertreters der Rhynchonelliformea
Aufbau eines Vertreters der RhynchonelliformeaZusammengehalten werden die beiden Schalenhälften durch eine starke Muskulatur, deren wichtigste Teile der „Öffner“ und der „Schließer“ sind. Diese Muskeln arbeiten gegeneinander und ermöglichen die Öffnung bzw. die Schließung der Schalen. Bei den Articulata existiert außerdem ein Scharnier, die beiden Schalen sind also miteinander artikuliert (Name!). Am Schalenaußenrand befinden sich Borsten (Setae). Der Öffnungswinkel der Klappen ist abhängig von der jeweiligen Art und kann zwischen 7° bei Gryphus-Arten und 45° bei Platidia-Arten betragen.
Im Zentrum des Tieres ist der namensgebende Armapparat (Lophophor) befestigt, der häufig (bei den Testicardines) durch ein kalkiges Armskelett an der Dorsal- oder Armschale gestützt wird, dem Lophophorskelett oder Brachidium, und mit dem das Tier Nahrung aus dem Wasser sammelt und atmet. Die Nahrung gelangt über den zentral gelegenen und schlitzförmigen Mund in das Verdauungssystem. Der Körper ist von einem zweilappigen Mantel umgeben, der an der Außenseite die Schalenmaterialien absondert. Der Weichkörper einschließlich der Lophophoren ist vollständig von beiden Gehäuseklappen umschlossen. Die inneren Organe liegen im hinteren Gehäusebereich, der vordere besteht aus einer Mantelhöhle, in die die Lophophoren hineinragen. Bei geschlossenen Klappen wird der Lophophor in die Filterkammer eingelegt und er stellt sich erst wieder auf, wenn die Klappen wieder geöffnet werden.
 Lingula anatina im Aquarium
Lingula anatina im AquariumAus der Ventral- oder Stielklappe, oder zwischen beiden Klappen, tritt am hinteren Ende des Tieres häufig ein muskulöser Stiel mit zäher, chitiniger, Cuticula aus, mit dem das Tier am oder im Substrat festgewachsen ist. Dieser Stiel ist beweglich und erlaubt den Tieren, eine optimale Position im Wasser einzunehmen. Wenige Arten besitzen keinen Stiel und kleben mit der Ventralklappe am Substrat, andere, etwa Ligula spec., sind nicht festgewachsen (sessil) und bewegen sich mit Hilfe des Stieles durch das Substrat (vagil).
Innere Anatomie
Das Nervensystem der Armfüßer ist sehr einfach aufgebaut. Es besitzt Ganglienkonzentrationen zur neuronalen Versorgung des Lophophor und der Klappenmuskulatur. Als Sinnesorgane sind nur Statocysten und einfache Lichtsinneszellen der Larven sowie bei Ligula bekannt. Der Darm ist bei den verschiedenen Taxa der Armfüßer unterschiedlich aufgebaut. Er folgt bei allen Arten einem erweiterten Magen mit einer paarigen Mitteldarmdrüse, verläuft dann jedoch entweder u-förmig zu einem vorn gelegenen After (Crania), auf die rechte Körperseite ausführend (alle Inarticulata) oder er endet blind (bei allen Testicardines).
Die Leibeshöhle, das Coelom, ist bei den Armfüßern dreiteilig, wobei die einzelnen Teile vollkommen voneinander abgetrennt sind. So besitzen sie ein röhrenförmiges Protocoel am Innenrand des Lophophor, das als „großer Armsinus“ oder Labialkanal benannt wird. Das Mesocoel stellt ebenfalls einen Kanal am Lophophor dar, dieser liegt allerdings als „kleiner Armsinus“ an der Außenseite und besitzt Seitenzweige in den Tentakeln. Das Metacoel ist paarig und bildet im Körperzentrum der Tiere Mesenterien mit darin enthaltenen Gefäßen sowie zwei Paar seitliche Bänder, das Gastroparietal- und das Ilioparietalband. Außerdem verzweigt es sich im Mantel. Das Gefäßsystem ist geschlossen und besteht aus einem Sinus und mehreren Blutkanälen, das Hämoglobinhaltige Blut wird von einem kontraktilen Herzen, das oberhalb des Magens liegt, durch den Körper gepumpt. Besonders gut versorgt werden die Außenbereiche des Mantels sowie der Lophophor, da hier der Sauerstoff zur Atmung aus dem Wasser aufgenommen wird.
Die Entsorgung von Stoffwechselendprodukten (Exkreten) erfolgt über ein Paar Metanephridien im Ilioparietalband, bei den Rhynchonellidae über zwei Paare im Gastroparietalband. Durch die Nephridien werden auch die Keimzellen ausgeleitet, die Gonaden liegen dabei im Bereich des Metacoels, entweder an den Mesentherien oder in den Mantelkanälen.
Lebensweise
Ernährung
Armfüßer sind typische sessile Filtrierer, sie filtern also ihre Nahrung aus dem vorbeiströmenden Wasser. Dabei bilden sie keine Kolonien, sind also solitär, liegen jedoch sehr häufig nahe beieinander. Die Nahrung der Tiere besteht hauptsächlich aus Dinoflagellaten und Kieselalgen (Bacillariophyta), die sie mit Tentakeln an den Lophophoren heranstrudeln. Der Lophophor hat zu diesem Zweck an der Oberseite einseitig angeordnete Rinnen und Tentakel mit einer reusenartigen Bewimperung. Durch die Bewegung des Lophophors wird entsprechend ein Wasserstrom erzeugt, der seitlich in die geöffneten Klappen eintritt, und die Nahrungspartikel aus dem Wasser gefiltert. Über die Nahrungsrinne wird die Nahrung zur zentral gelegenen Mundöffnung geleitet.
Fortpflanzung und Entwicklung
 Lebende Brachiopoden
Lebende BrachiopodenArmfüßer sind mit Ausnahme der Vertreter der Gattung Argyrotheca getrenntgeschlechtlich und pflanzen sich ausschließlich sexuell fort. Dabei entwickeln sich die Keimzellen in den Gonaden und werden durch die Nephridien in das freie Wasser entlassen, wo die Befruchtung stattfindet. Bei einigen Arten verbleiben die Eizellen im Bereich des Lophophor oder sogar noch in den Nephridien und werden hier von freischwimmenden Spermien befruchtet, die Larvalentwicklung erfolgt bei ihnen anschließend in der Schale (Brutpflege).
Nach der Befruchtung beginnt die Keimesentwicklung durch eine Radiärfurchung bis zum Stadium der Gastrula. Das Mesoderm wird durch eine Enterocoelie des Darmes gebildet, es schnüren sich also Bläschen desselben ab und bilden die drei paarigen Leibeshöhlen (Coelom). Bei den Inarticulata findet die Larvalentwicklung weitestgehend in der Eihülle statt und die Tiere schlüpfen als Larve, die den ausgewachsenen Tieren weitestgehend ähnelt und sich in dieser Form festsetzen kann. Bei den Articulata entwickelt sich eine freischwimmende Larve mit einer morphologischen Zweiteilung, wobei der Vorderteil mit Zilien ausgestattet ist und der Fortbewegung dient.
Nach etwa zwei Tagen heftet sich die Larve mit dem Hinterteil an das Substrat und es beginnt eine Metamorphose, bei der sich aus dem Hinterteil der Stiel und der Mantel bilden. Letzterer stülpt sich über das Vorderende, welches sich zum eigentlichen Körper des Tieres mit dem Lophophor entwickelt. Die Schalen werden abschließend vom Mantel sezerniert, indem sich erst eine Vorschale aus Proteinen bildet (Protegulum). Die eigentliche Schale bildet sich von diesen Protegula ausgehend, die später den zentralen Wirbel bilden. Der Zuwachs kann anhand von Zuwachslinien abgelesen werden, wodurch sich das Alter der Tiere an der Schale abschätzen lässt. Das Lebensalter kann von wenigen Monaten bei Kleinformen bis zu mehreren Jahren reichen, eine ähnliche Altersspanne wird auch für fossile Formen angenommen.
Stammesgeschichte
Phylogenie
Entstehung der Armfüßer
 Platystrophia ponderosa
Platystrophia ponderosaDie Armfüßer sind fossil bereits seit dem frühen Kambrium vor etwa 530 Millionen Jahren bekannt. Dabei sind bis heute Fossilien von mehr als 30.000 ausgestorbenen Arten gefunden worden, die in mehr als 4000 Gattungen eingeordnet werden. Besonders artenreich war das Taxon im Devon (siehe hierzu Abschnitt „Entfaltung der Armfüßer“).
Man geht davon aus, dass die Armfüßer einen gemeinsamen Vorfahren haben und entsprechend monophyletisch sind. Zu möglichen fossilen Vertretern dieser Vorfahren gibt es allerdings keine überzeugenden Kandidaten, folglich ist die evolutionäre Entstehung des Armfüßer-Bauplanes ein umstrittenes Problem. Auf paläontologischer Basis wurden Beziehungen zu den frühkambrischen Halkieriida hergestellt, einer ausgestorbenen Tiergruppe, welche zwei Schalen auf der Rückenseite trägt. Die Gegner dieser Hypothese bezweifeln unter anderem, dass aus zwei rückenseitig gelegenen Schalen allmählich die Rücken- und Bauchklappe der Armfüßer geworden sein könnte.
Im Vergleich mit heutigen Tierstämmen wurde aufgrund des charakteristischen Lophophor-Organs eine nahe Verwandtschaft mit den Hufeisenwürmern und Moostierchen vermutet, diese beiden Gruppen wurden mit den Armfüßern zu einem Taxon Lophophorata oder Tentaculata vereinigt. Mittels molekularbiologischer Methoden konnte bisher nur die Vermutung eines Schwestergruppenverhältnisses zu den Hufeisenwürmern bekräftigt werden. Im Rahmen der ebenfalls molekularbiologisch begründeten Lophotrochozoa-Hypothese wurde eine Abstammung der Armfüßer und Hufeisenwürmer von ringelwurmartigen Vorfahren (Annelida) in Betracht gezogen, eine Hypothese, die bereits 1871 von E.S.Morse auf morphologischer Basis vertreten worden war. Als morphologische Indizien, die für eine Beziehung der Armfüßer zu den Ringelwürmern sprechen, werden unter anderem identisch aufgebaute Borsten genannt.
 Neospirifer condor aus dem Karbon
Neospirifer condor aus dem KarbonEntfaltung der Armfüßer
Im Kambrium lebten vor allem inarticulate Brachiopoden. Dies änderte sich im Ordovizium, als articulate Formen eine erste große Radiation durchmachten und die inarticulaten in der Artenzahl überflügelten. Eine weitere Zunahme der Artenzahlen erfolgte, mit einem leichten Einbruch im Silur, bis zum Devon, wo die höchsten Artenzahlen in insgesamt sieben Ordnungen bekannt sind. Nach dem Devon kam es zu einem erneuten Einschnitt, nach dem sich vor allem die Strophomeniden stark vermehrten. Der größte Einschnitt in die Artenvielfalt erfolgte zum Abschluss des Perm vor 251 Millionen Jahren. Bei diesem größten bekannten Massenaussterben verschwanden etwa 95 Prozent der marinen Invertebraten wie etwa die Großforaminiferen, die paläozoische Korallen, die Trilobiten und die Eurypteriden, während die Armfüßer ebenso wie die Bryozoen, die Crinoiden und die Ammoniten stark dezimiert wurden. Bei den Armfüßern wird angenommen, dass das Aufkommen neuer Fressfeinde, der hochangepassten Seesterne nach der Perm-Trias-Grenze das Aussterben noch verstärkt haben könnte.
In der Trias existierten nur noch sehr wenige Armfüßer-Gruppen, die größtenteils bis heute noch existieren. Das Massenaussterben am Ende der Kreidezeit vor etwa 65 Millionen Jahren, bei dem neben den Großtieren wie Dinosaurier, Flugsaurier und vielen anderen vor allem auch die Ammoniten vollständig ausstarben, hatte auf die Armfüßer nur wenig Einfluss. Andererseits entwickelten sich seit der Trias auch die Muscheln, die teilweise als direkte Konkurrenten auftraten, sodass es zu keiner weiteren Radiation kam. Heute leben noch etwa 300 Arten articulater Brachiopoden, die 75 Gattungen zugeordnet werden. Bei den inarticulaten Formen starben die meisten Formen bereits im Ordovizium und Devon aus, bis heute haben nur wenige Arten der Ligulidae und der Discinidae in fünf Gattungen und der Craniidae mit kalkiger Schale in drei Gattungen überlebt.
Stratigraphie
Aufgrund ihres zeitlich sehr gut einzuordnenden Vorkommens und ihrer Merkmalskombinationen sowie ihres zahlenmäßig häufigen Vorkommens stellen die Armfüßer wichtige Leitfossilien dar, anhand derer sich das Alter von anderen Fossilfunden aus gleichen Schichten sehr gut abschätzen lässt. Sie sind außerdem vor allem da häufig zu finden, wo andere wichtige Leitfossilien wie die Graptolithen oder die Goniatiten fehlen.
Systematik
Taxonomie der Armfüßer
Taxa mit rezenten Arten in grün, mit ausschließlich fossilen Arten in weiß
nach Williams, Carlson und Brunton, 2000Unterstamm Klasse Ordnung Ausgestorben Linguiliformea Lingulata Linguilida nein Siphonotretida Ordovizium Acrotretida Devon Paterinata Paterinida Ordovizium Craniformea Craniforma Craniida nein Craniopsida Karbon Trimerellida Silur Rhynchonelliformea Chileata Chileida Kambrium Dictyonellidina Perm Obolellata Obolellida Kambrium Kutorginata Kutorginida Kambrium Strophomenata Orthotetidina Perm Triplesiidina Silur Billingselloidea Ordovizium Clitambonitidina Ordovizium Strophomenida Karbon Productida Perm Rhynchonellata Protorthida Kambrium Orthida Karbon Pentamerida Devon Rhynchonellida nein Atrypida Devon Spiriferida Jura Thecideida nein Athyridida Kreide Terebratulida nein Heute existieren etwa 83 Gattungen mit etwa 375 Arten von Armfüßern. Die Systematik dieser Tiergruppe ist bis heute nicht vollständig geklärt. Klassisch werden die rezenten Arten ohne Schalenschloß zu den Inarticulata zusammengefasst, während alle schlosstragenden Armfüßer die Articulata bzw. Testicardines bilden. Letztere haben sich jedoch aus Vertretern der Inarticulata entwickelt, wodurch die Inarticulata eine paraphyletische Gruppe bilden und nicht mehr als Taxon gelten. Die klassische Einteilung sieht entsprechend folgendermaßen aus:
- Inarticulata oder Ecardines
- Atremata – mit chitiniger Schale, unter anderem Lingula unguis und Glottina pyramidata
- Neotremata – mit kalkiger Schale, unter anderem Crania anomalia
- Articulata oder Testicardines
- Rhynchonellida
- Terebratulida
Vor allem in der Paläontologie wird heute auf der Basis rezenter und fossiler Arten ein Stammbaum diskutiert, der die Phylogenese darstellt. Dieser wurde 1996 von Williams et al. entwickelt:
Brachiopoda ├─Linguliformea │ ├─Lingulata │ └─†Paterinata ├─Craniformea (mit Craniata) └─Rhynchonelliformea ├─†Chileata └─N.N. ├─†Obolellata ├─†Kutorginata └─Articulata ├─†Strophomenata └─RhynchonellataDie Articulata bilden auch hier eine monophyletische Gruppe, allerdings innerhalb der Rhynchonelliformea. Sie umfassen die Klassen Strophomenata und Rhynchonellata, letztere bilden die einzige überlebende Gruppe der Rhynchonelliformea. Die Inarticulata werden als paraphyletische Gruppe aufgelöst in die Linguliformea und die Craniiformea.
Eine vollständigere Darstellung stellten Williams, Carlson und Brunton 2000 vor, die in der nebenstehenden Tabelle dargestellt wird.
Literatur
- Sievert Lorenzen: Brachiopoda, Armfüßer, in: W. Westheide, R. Rieger (Hrsg.): „Spezielle Zoologie Teil 1: Einzeller und Wirbellose Tiere“; Gustav Fischer Verlag, Stuttgart, Jena.
- Bernhard Ziegler: Einführung in die Paläobiologie Teil 3 – ISBN 3-510-65179-0
- E.N.K. Clarkson: Invertebrate Palaeontology and Evolution – ISBN 0-632-05238-4
- U. Lehmann, G. Hillmer: Wirbellose Tiere der Vorzeit – ISBN 3-432-90653-6
- A. Williams, S.J. Carlson, C.H.C. Brunton, L.E. Holmer, L. Popov: A supra-ordinal classification of the Brachiopoda. In: Philosophical Transactions of the Royal Society of London, Series B 351, 1996, S. 1171–1193
- Carlson, S.J., Sandy, M.R. (Hrsg.): Brachiopods Ancient and Modern: A Tribute to G. Arthur Cooper. The Paleontological Society Special Publications 7, New Haven, 2001, ISSN 1089-3326
- Williams, A., Carlson, S.J. und Brunton, C.H.C.: Brachiopod classification in Williams, A. et al. Brachiopoda (revised), Treatise on Invertebrate Paleontology (Kaesler, R.L., ed.). Boulder, Colorado: Geological Society of America and Lawrence, Kansas: The University of Kansas, 2000, ISBN 0-8137-3108-9
Weblinks




Wikimedia Foundation.