- Adhäsions-Plaque
-
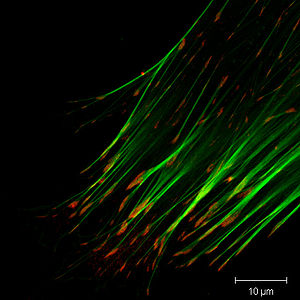 Immunfluoreszenzfärbung des Aktin-Zytoskeletts (grün) und des fokalen Adhäsionsproteins Vinculin (rot) bei einer Fibroblasten-Zelle. Die Adhäsionsstellen sind als rote Flecken an den Enden der Aktin-Bündel erkennbar.
Immunfluoreszenzfärbung des Aktin-Zytoskeletts (grün) und des fokalen Adhäsionsproteins Vinculin (rot) bei einer Fibroblasten-Zelle. Die Adhäsionsstellen sind als rote Flecken an den Enden der Aktin-Bündel erkennbar.Fokale Adhäsionen (engl. focal adhesion) sind verankernde Zellverbindungen, die das Aktin-Zytoskelett einer Zelle mechanisch an das Substrat (Extrazelluläre Matrix, kurz: ECM) koppeln. Sie sind auf klar abgegrenzte Bereiche der Zelle beschränkt, an denen die Plasmamembran bis auf 15 nm an das Substrat heranreicht[1]. Fokale Adhäsionen können sich aus über 50 verschiedenen Proteinen zusammensetzen, was auf einen beträchtlichen Funktionsumfang schließen lässt[2].
Tatsächlich dienen sie nicht nur der Verankerung der Zelle, sondern fungieren darüber hinaus als Signalüberträger, die die Zelle über den Zustand der ECM informieren und ihr Verhalten somit beeinflussen[3]. Bei sessilen Zellen sind Fokale Adhäsionen im Normalfall recht stabil, während sie bei wandernden Zellen kurzzeitig auf- und abgebaut werden. Dies spielt beispielsweise eine wichtige Rolle für die Immunabwehr, bei der Leukozyten entlang des Gefäßendothels eine abbremsende Rollbewegung vollführen und schließlich in entzündetes Gewebe einwandern.
Inhaltsverzeichnis
Bau
Der Kontakt zu Proteinen der ECM wird hauptsächlich über transmembrane Integrine vermittelt, die sich zu großen Proteinclustern zusammenlagern und mit ihrer extrazellulären Domäne an Proteine mit spezifischer Aminosäuresequenz (RGD-Sequenz) binden (z. B. Fibronektin, Laminin, Vitronektin oder Kollagen). Integrine sind Heterodimere und setzen sich jeweils aus einer α- und einer β-Untereinheit zusammen. Diese Untereinheiten liegen in verschiedenen Isoformen vor, die sich in ihrer Bindungsfähigkeit zu ECM-Proteinen unterscheiden. Die β-Untereinheit ist auf der intrazellulären Seite über Adapterproteine (z. B. Talin, α-Aktinin, Filamin und Vinculin) an Aktinfilamente gekoppelt. Neben strukturgebenden Proteinen finden sich in Fokalen Adhäsionen viele signalübertragende Proteine wie die Tyrosinkinasen c-Src oder FAK (focal adhesion kinase).[2]
Adhäsionsdynamik bei migrierenden Zellen
Der dynamische Auf- und Abbau von fokalen Adhäsionen spielt vor allem bei der Zellmigration eine zentrale Rolle. Bei migrierenden Zellen ändert sich die Proteinzusammensetzung und Morphologie von Fokalkontakten im Laufe ihres Bestehens, weshalb man sie in verschiedene Typen einteilen kann. Frühe Adhäsionen entstehen beim Vorschub des Lamellipodiums (Protrusion) nahe der Zellfront und werden Fokalkomplexe genannt. Sie sind recht klein (0.25μm²) und enthalten neben α₅β₃-Integrin andere Proteine wie Talin, Paxilin und Phosphotyrosin. Die meisten Fokalkomplexe lösen sich während des Zurückziehens des Lamellipodiums (Retraktion) wieder auf. Die übrigen vergrößern sich und reifen zu stabilen fokalen Adhäsionen, indem sie weitere Proteine wie z. B. Zyxin rekrutieren.[1] Fokale Adhäsionen bewegen sich relativ zum Substrat nur sehr wenig, sodass die Zelle im Verlauf der Migration über sie hinwegwandert. Die Adhäsionsstellen bewegen sich daher relativ zur Zelle von vorne nach hinten.
Am hinteren Ende der Zelle müssen die Fokalen Adhäsionen aufgelöst werden, um das Nachziehen des Zellkörpers zu ermöglichen. Der Mechanismus dieses sogenannten rear detachment ist bislang noch recht wenig verstanden und erfolgt wahrscheinlich auf verschiedene Arten. Die Trennung der Verbindung zwischen Aktin-Zytoskelett und Substrat kann prinzipiell rein mechanisch durch zelluläre Kontaktionskräfte oder biochemisch erfolgen. Es konnte gezeigt werden, dass die Hemmung sogenannter Calpain-Proteasen zur Stabilisierung von fokalen Adhäsionen führt und die Substratablösung verringert. Da zu den Substraten von Calpain einige Proteine fokaler Adhäsionen gehören, könnte es sein, dass diese im Zuge des rear detachments gezielt abgebaut werden.[4]
Funktion als Mechanosensor
Mechanische Kräfte, die auf Fokale Adhäsionen ausgeübt werden, können das intrazelluläre Signalprotein Src aktivieren und bewirken das Wachstum der Adhäsionen. Dies zeigt, dass Fokale Adhäsionen als Mechanosensoren fungieren, und weist darauf hin, dass myosinvermittelte Zugkräfte zur Reifung der Fokalkomplexe beitragen könnten.[5][3]
Literatur
- Alberts, B. et. al.: Molecular Biology of the Cell. Garland Science, 4. Auflage, 2002. ISBN 0815340729.
Quellen
- ↑ a b Zaidel-Bar, R., M. Cohen, L. Addadi und B. Geiger: Hierarchical assembly of cell–matrix adhesion complexes. Biochemical Society Transactions, 32:416–420, 2004
- ↑ a b Zamir, E. und B. Geiger: Molecular complexity and dynamics of cell-matrix adhesions. Journal of Cell Science, 114:3583–3590, 2001
- ↑ a b Rivelinea, D., E. Zamir, N.Q. Balabanb, U.S. Schwarzc, T. Ishizakid, S. Narumiyad, Z. Kamb, B. Geiger und A.D. Bershadsky: Focal contacts as mechanosensors: externally applied local mechanical force induces growth of focal contacts by an mDia1-dependent and ROCK-independent mechanism. The Journal of Cell Biology, 153(6):1175–1186, 2001
- ↑ Huttenlocher, A., S.P. Palecek, Q. Lu, W. Zhang, R.L. Mellgren, D.A. Lauffenburger, M.H. Ginsberg und A.F. Horwitz: Regulation of cell migration by the calcium-dependent protease calpain. Journal of Biological Chemistry, 272:32719–32722, 1997.
- ↑ Wang, Y., E.L. Botvinick, Y. Zhao, M.W. Berns, S. Usami, R.Y. Tsien und S. Chien: Visualizing the mechanical activation of Src. Nature, 434:1040– 1045, 2005
Siehe auch
- Zellkontakt Übersicht über die verschiedenen Arten von Zellkontakt
- Tight Junction
- Adherens Junction
- Desmosom
- Hemidesmosom
- Gap Junction
Weblinks
Wikimedia Foundation.