- Eisen-Schwefel-Welt
-
Die chemische Evolution ist die Entstehung der organischen Molekülen aus anorganischen Molekülen (Abiogenese) im Hadaikum zwischen der Entstehung der Erde vor etwa 4,6 Milliarden Jahren und der Entstehung des Lebens (Beginn der biologischen Evolution) vor etwa 4,2 bis 3,8 Milliarden Jahren.
Aus abiotisch-anorganischen Molekülen entwickelten sich unter Einwirkung von Energie zunächst organische Verbindungen und präbiotische Moleküle, aus denen später erste Lebewesen hervorgingen. Vergleichbare Prozesse sind möglicherweise auch überall dort im Universum möglich, wo gemäßigte Temperaturzonen existieren (z. B. auf Planemos/Exoplaneten). Die Entstehung von organischen Stoffen, also von Kohlenstoff-Wasserstoff-Verbindungen, wird als Voraussetzung für die Entstehung von Lebewesen angesehen, ebenso wie z. B. das Vorhandensein gemäßigter Temperaturen, die Abwesenheit energiereicher elektromagnetischer Strahlung und die Gegenwart von Wasser als Lösungsmittel und Medium (Kosmochemie).
Inhaltsverzeichnis
Einleitung
Die Rekonstruktion dieser Entwicklungsgeschichte ist schwierig, weil Fossilien und genaue Kenntnisse der geochemischen Verhältnisse der Erde vor etwa 3,8 Milliarden Jahren fehlen.
Deshalb existieren verschiedene Hypothesen zum Ablauf der chemischen Evolution. Sie werden hauptsächlich durch Experimente gestützt, die auf Annahmen über die damalige chemische Zusammensetzung der Atmosphäre, Hydrosphäre und Lithosphäre sowie klimatische Bedingungen beruhen. Auf die Ursprünge der Hydrosphäre geht der Artikel Herkunft des irdischen Wassers näher ein.
Die Experimente, welche die Hypothesen stützen, reichen momentan nicht aus zur Formulierung einer Theorie, die erklären kann, wie das Leben entstand. So konnte zwar bereits die chemische Entstehung von komplexen Molekülen beobachtet werden, die für biologische Abläufe notwendig sind, jedoch noch keine Bildung eines hinreichenden Systems daraus. Diese Beobachtungen werden aber bereits als großer Erfolg und ausreichend für die Aufstellung der Hypothese gewertet.
Übersicht
Hypothesen zur chemischen Evolution müssen verschiedene Aspekte erklären:
- Die abiogene Entstehung der Biomoleküle, das heißt ihre Entwicklung aus nichtlebenden beziehungsweise nichtorganischen Vorläufern (Kosmochemie).
- Die Entstehung sich selbst replizierender und variierender chemischer Informations-Systeme, das heißt die Entstehung der Zelle.
- Die Entstehung der gegenseitigen Abhängigkeit von Funktion (Enzyme) und Information (RNA, DNA).
- Die Umweltbedingungen der Erde vor 4,5 bis 3,5 Milliarden Jahren (oder möglicherweise derzeit existierender vergleichbarer Himmelskörper wie Exoplaneten, Monde und Planemos).
Beiträge dazu kommen unter anderem von folgenden Wissenschaftlern:
- Alexander Oparin: Koazervate (siehe unten)
- Harold C. Urey und Stanley L. Miller 1953: Entstehung einfacher Biomoleküle in einer simulierten Uratmosphäre (siehe unten)
- Sidney W. Fox: Mikrosphären aus Protenoiden (siehe unten)
- Thomas R. Cech (Universität von Colorado) und Sidney Altman (Yale-Universität New Haven Connecticut) 1981: autokatalytisches RNA-Splicing: „Ribozyme“ vereinigen Katalyse und Information in einem Molekül. Sie vermögen sich aus einer längeren RNA-Kette selbst herauszuschneiden und die verbleibenden Enden wieder zusammenzufügen.
- Walter Gilbert (Harvard-Universität Cambridge) entwickelt 1986 die Idee der RNA-Welt (siehe unten)
- Günter von Kiedrowski (Ruhr-Universität Bochum) veröffentlicht 1986 das erste selbstreplizierende System auf der Grundlage eines Hexanukleotids (DNA), wichtige Beiträge zum Verständnis der Wachstumsfunktionen selbstreplizierender Systeme.
- Manfred Eigen (Max-Planck-Institut für biophysikalische Chemie Göttingen): Evolution von RNA-Protein-Ensembles. Hyperzyklus.
- Julius Rebek jr. (MIT Cambridge) stellt ein künstliches Molekül her (Aminoadenosintriazidester), das sich in Chloroformlösung selbst repliziert. Allerdings sind die Kopien identisch mit der Vorlage, so dass eine Evolution für diese Moleküle nicht möglich ist.
- John B. Corliss (Goddard-Raumfahrtzentrum der NASA): Hydrothermale Schlote der Meere liefern Energie und Chemikalien, die eine von Meteoriten-Einschlägen weitgehend ungestörte chemische Evolution ermöglichen. Heute noch sind sie Lebensraum für die in vielen Merkmalen sehr urtümlichen Archaebakterien (Archaea).
- Günter Wächtershäuser (München): Die ersten sich selbst replizierenden Strukturen mit Stoffwechsel seien auf der Oberfläche von Pyrit entstanden. Das Eisensulfid des Pyrits habe hierzu die notwendige Energie geliefert. Mit den wachsenden und wieder zerfallenden Pyritkristallen hätten diese Systeme wachsen und sich vermehren können und die verschiedenen Populationen seien unterschiedlichen Umweltbedingungen (Selektionsbedingungen) ausgesetzt gewesen.
- A. G. Cairns-Smith (Universität Glasgow) und David C. Mauerzall (Rockefeller-Universität New York) sehen in Tonmineralien ein System, das zunächst selbst einer chemischen Evolution unterworfen ist, wodurch viele verschiedene, sich selbst replizierende Kristalle entstehen. Diese Kristalle ziehen auf Grund ihrer elektrischen Ladung organische Moleküle an und katalysieren die Synthese komplexer Biomoleküle, wobei der Informationsgehalt der Kristallstrukturen zunächst als Matrize dient. Diese organischen Gebilde werden immer komplexer, bis sie sich ohne Hilfe der Tonmineralien vermehren können.
- Wolfgang Weigand, Mark Dörr et al. (Friedrich-Schiller-Universität Jena) zeigen 2003, dass Eisensulfid die Synthese von Ammoniak aus molekularem Stickstoff katalysieren kann.
Noch steht ein einheitliches Modell zur chemischen Evolution aus, möglicherweise weil grundlegende Prinzipien noch nicht entdeckt wurden.
Vorüberlegungen
Biomoleküle
Die präbiotische Entstehung der komplexen organischen Moleküle kann in drei Schritte unterteilt werden:
- Entstehung einfacher organischer Moleküle (Alkohole, Säuren, Heterozyklen wie Purine und Pyrimidine) aus anorganischen Stoffen.
- Entstehung der Grundbausteine (Einfachzucker, Aminosäuren, Pyrrole, Fettsäuren, Nukleotide) komplexer organischer Moleküle aus einfachen organischen Molekülen.
- Entstehung der komplexen organischen Moleküle aus den Grundbausteinen.
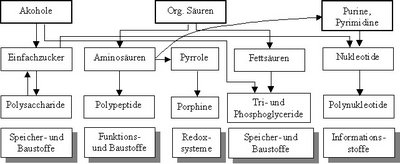 Biomoleküle – Entstehung und Funktion
Biomoleküle – Entstehung und FunktionDie Elementaranalyse dieser Moleküle führt zu der Frage, welche anorganischen Verbindungen zu ihrer Entstehung notwendig waren.
Zusammensetzung der Biomoleküle: C H O N S P Kohlenhydrate X X X Lipide X X X X X Proteine X X X X X Nucleotide X X X X X Porphine X X X X mögliche anorganische Quelle der Elemente: reduziert oxidiert Kohlenstoff (C) Methan (CH4) Kohlenstoffdioxid (CO2), Kohlenstoffmonoxid (CO) Wasserstoff (H) Wasserstoff (H2) Wasser (H2O) Sauerstoff (O) H2O Sauerstoff (O2) Stickstoff (N) Ammoniak (NH3) Nitrat (NO3-) Schwefel (S) Schwefelwasserstoff (H2S) Sulfat (SO42-) Phosphor (P) Phosphin (PH3) Phosphat (PO43-) Alle Hypothesen gehen davon aus, dass neben Wasser und Phosphat zunächst nur die reduzierten Formen der heute üblichen chemischen Verbindungen in ausreichender Menge zur Verfügung standen, da die Uratmosphäre kaum molekularen Sauerstoff enthielt.
Als Energiequelle werden UV-Strahlen, Wärme vulkanischer Prozesse, ionisierende Strahlen radioaktiver Prozesse und elektrische Entladungen angenommen. Nach einigen sehr beachtenswerten neueren Theorien käme die nötige Energie für die Entstehung von Bio-Molekülen alternativ dazu auch aus anaeroben Redoxprozessen zwischen reduzierten vulkanischen Gasen und teilweise oxidierten sulfidischen Mineralien wie Pyrit (FeS2).
Die Rolle des Wassers für die Evolution des Lebens
 H2O ist die einzige chemische Verbindung, die auf der Erde unter natürlichen Bedingungen in allen drei Aggregatzuständen vorkommt.
H2O ist die einzige chemische Verbindung, die auf der Erde unter natürlichen Bedingungen in allen drei Aggregatzuständen vorkommt.Leben, wie wir es kennen (bzw. definieren), benötigt Wasser als universelles Lösungsmittel. Es hat Eigenschaften, die nach akzeptiertem naturwissenschaftlichem Verständnis die Entstehung von Leben ermöglichen (siehe auch anthropisches Prinzip). Möglicherweise kann Leben unabhängig von Wasser entstehen und existieren, viele Wissenschaftler nehmen aber an, dass die Anwesenheit von flüssigem Wasser (in einem bestimmten Gebiet oder auf einem bestimmten Planeten, wie dem Mars) unsere Art Leben nicht nur ermöglicht, sondern sein Entstehen sogar sehr wahrscheinlich macht.
Für die Entstehung des Lebens sind folgende Eigenschaften des Wassers relevant:
- Wasser ist in einem Temperaturbereich flüssig, in dem organische Moleküle stabil sind.
- Wasser ist als polares Medium für chemische Reaktionen besonders geeignet, da es eine homogene Durchmischung ermöglicht, Protonen für Katalysen zur Verfügung stellen kann und eine hohe Wärmekapazität hat und somit überschüssige Reaktionswärme aufnimmt.
- Wasser weist global geringe Schwankungen in Temperatur und osmotischen Werten auf (lokal können große Unterschiede entstehen), was zu einem global ausgeglichenen Klima führt.
- Wasser absorbiert die für Makromoleküle schädliche UV-Strahlung. Diese durchdringt aber gefrorenes Wasser (Eis) bis zu einer gewissen Tiefe.
- Wasser, in dem Stoffe gelöst sind, wie Meerwasser, bildet beim Ausfrieren Bereiche unterschiedlicher Stoffkonzentrationen, die von Eismembranen umgeben sind (siehe Meereis-Hypothese). Diese Kompartimentierung und die entstehenden Konzentrationsgefälle werden als notwendig für die Entstehung von biologisch aktiven Molekülen angesehen.
- Die Anomalie des Wassers verhindert, dass Gewässer vom Grund aus vereisen und sorgt für einen Bereich gleichmäßiger Temperatur.
Entwicklung der Erdatmosphäre
Hauptartikel: Entwicklung der Erdatmosphäre
Die Entwicklung der Erdatmosphäre ist ein Teil der chemischen Evolution und zudem ein wichtiges Element der Klimageschichte. Sie wird heute in vier wesentliche Entwicklungsstufen unterschieden:
(1) Uratmosphäre
Am Anfang standen die Entstehung der chemischen Elemente im Universum (beschrieben in der Kosmochemie) und die Entstehung der Erde aus diesen - vor etwa 4,56 Milliarden Jahren. Dabei verfügte unser Planet Erde schon sehr früh über eine vermutlich aus Wasserstoff (H2) und Helium (He) bestehende Gashülle, die jedoch wieder verloren ging. Astronomen gehen darüber hinaus sogar davon aus, dass zunächst bei der Entstehung des Sonnensystems auf der Erde und allen anderen sonnennahen Planeten wegen der relativ hohen Temperaturen und den Effekten des Sonnenwinds nur wenig oder keine leichten Elemente (inklusive Kohlenstoff, Stickstoff und Sauerstoff) „übrig“ geblieben sind. All diese Elemente, die heute den Hauptanteil an der Biosphäre ausmachen, wären nach dieser Theorie erst nach geraumer Zeit durch Kometeneinschläge aus den äußeren Bereichen des Sonnensystems angeliefert worden, nachdem sich die Protoplaneten etwas abgekühlt hatten. Da sich während der ersten einigen hundert Millionen Jahre nach Entstehung des Sonnensystems ständig große Einschlagereignisse von Himmelskörpern wiederholten, wären lebende Systeme, die sich bereits in diesen Zeiten entwickelten, immer wieder durch globale Sterilisationen vernichtet worden, die durch große Kollisionen verursacht wurden. Die Entwicklung von Leben konnte so erst starten, nachdem sich flüssiges Wasser zumindest an den tiefsten Stellen der Meere auf Dauer halten konnte.
(2) Erste Atmosphäre
 Zeichen vulkanischer Aktivität: Schwefelablagerungen am Rand des Halema'uma'u-Kraters des Kilauea auf Hawaii
Zeichen vulkanischer Aktivität: Schwefelablagerungen am Rand des Halema'uma'u-Kraters des Kilauea auf Hawaii Vulkanausbrüche sind die spektakulärsten Erscheinungsformen des Vulkanismus
Vulkanausbrüche sind die spektakulärsten Erscheinungsformen des VulkanismusDurch die langsame Abkühlung der Erde, den dabei auftretenden Vulkanismus (Ausgasung aus dem Erdinneren) und die globale Verteilung der Materie eingeschlagener Kometen kam es zur Etablierung einer Atmosphäre. Darin sind als Verbindungen vor allem Wasserdampf (H2O; bis zu 80 %), Kohlendioxid (CO2; bis zu 20 %), Schwefelwasserstoff (bis 7 %), Ammoniak und Methan als Hauptbestandteile zu erwarten. Dabei handelt es sich um eben jene Produkte des Vulkanismus beziehungsweise der Kometen, die wir auch heute noch beobachten können. Der hohe Anteil des Wasserdampfs erklärt sich dadurch, dass die Erdoberfläche zu diesem Zeitpunkt noch zu warm war, um große Meere zu bilden. Das Wasser verdampfte zum Großteil sofort wieder, obwohl es über Millionen von Jahren pausenlos geregnet haben muss. Der eigentliche Ursprung des Wassers (Vulkane? Kometen?) ist jedoch noch teilweise umstritten. Vor allem aus Wasser, Methan und Ammoniak können sich unter den Bedingungen der frühen Erde zunächst kleine organische Moleküle (Säuren, Alkohole, Aminosäuren), später auch organische Polymere (Polysaccharide, Fette, Polypeptide) bilden, die in der oxidierenden Atmosphäre nicht stabil sind.
(3) Zweite Atmosphäre
Nachdem die Temperatur der Atmosphäre unter den Siedepunkt des Wassers fiel, kam es zu einem extrem langen Dauerregen, nach dessen Ende sich die Ozeane gebildet hatten. Dementsprechend wurden die anderen Atmosphärengase relativ zum Wasserdampf angereichert. Die hohe UV-Einstrahlung bedingte eine photochemische Zerlegung der Wasser-, Methan- und Ammoniakmoleküle, wodurch sich Kohlendioxid und Stickstoff ansammelten. Die leichten Gase wie Wasserstoff oder Helium verflüchtigten sich großteils in den Weltraum, Kohlendioxid löste sich in großen Mengen in den Ozeanen, wodurch ihr Wasser angesäuert wurde und ihr pH-Wert sich auf etwa 4 absenkte. Der inerte und wenig lösliche Stickstoff N2 blieb unverändert, sammelte sich mit der Zeit an und bildete vor etwa 3,4 Milliarden Jahren den Hauptbestandteil der Atmosphäre.
Die Ausfällung des Kohlendioxids mit Metallionen als Carbonate und die spätere Entwicklung von Lebewesen, die Kohlendioxid assimilierten, führte zu einer Verringerung der CO2-Konzentration und einem Wieder-Ansteigen der pH-Werte der Gewässer.
(4) Dritte Atmosphäre
Der Sauerstoff O2 spielt die Hauptrolle bei der weiteren Entwicklung hin zu unserer heutigen Atmosphäre. Er wurde durch das Auftreten von Lebewesen mit oxygener Photosynthese gebildet, und zwar seit möglicherweise etwa 3,5 Milliarden Jahren; vermutlich waren es Cyanobakterien oder Cyanobakterien-ähnliche Prokaryoten. Deren CO2-Assimilation führte zwar zu einem weiteren Absinken der Kohlenstoffdioxidkonzentration, die Sauerstoffkonzentration der Atmosphäre blieb jedoch gering. Der Grund dafür ist, dass O2 in den Ozeanen sofort zur Oxidation von zweiwertigen Eisenionen und anderen leicht oxidierbaren Stoffen verbraucht wurde und sich in der Atmosphäre erst langsam ansammelte, nachdem diese Verbindungen vor etwa zwei Milliarden Jahren fast komplett oxidiert waren.
Der sehr reaktive Sauerstoff reagiert leicht mit einigen empfindlichen organischen Biomolekülen. Das vermehrte Auftreten von Sauerstoff vor zwei Milliarden Jahren stellte deshalb für die frühen Organismen einen gewaltigen selektierenden Umweltfaktor dar. Nur wenige der damaligen vermutlich obligat anaeroben Organismen konnten sich in sauerstofffreie Lebensräume zurückziehen, ein anderer Teil entwickelte offenbar Enzyme, die toxische Peroxide oder andere Sauerstoff-Metabolite unschädlich machen (zum Beispiel Superoxid-Dismutasen, Peroxidasen und Katalasen). Schließlich entwickelten sich in einigen Mikroorganismen unter Beteiligung solcher Enzyme als Vorläufer-Formen sogar höchst komplexe Membran-Enzyme, die den anwesenden Sauerstoff metabolisch ausnutzen, um Energie für das Wachstum der eigenen Zellen zu erzeugen: die Endoxidasen der aeroben Atmungsketten. Je nach Organismus gibt es mehrere verschiedene Formen von Endoxidasen, wie Chinol-Oxidasen oder Cytochrom-c-Oxidasen, die jeweils auch noch unterschiedlich mit Kupfer-Ionen und Häminen enthaltenden aktiven Zentren ausgestattet sind und sich deshalb höchstwahrscheinlich aus mehreren parallelen Entwicklungslinien ableiten. In vielen Fällen sind sogar mehrere Endoxidase-Typen nebeneinander im selben Organismus zu finden und werden je nach den herrschenden Umweltbedingungen synthetisiert. Diese Enzyme stellen das letzte Glied von mehreren nacheinander geschalteten Enzymkomplexen dar, die die Energie aus Redoxprozessen zunächst durch Translokation von Protonen oder Na-Ionen in Form von Membranpotential „zwischenspeichern“. Das Membranpotential wird schließlich über einen weiteren Enzymkomplex, die ATP-Synthase, wieder zu chemischer Energie in Form von ATP umgewandelt. Die ATP-Synthase und die sonstigen Komponenten der Atmungskette müssen evolutionsgeschichtlich wesentlich älter als die Endoxidasen sein, da sie (bzw. sehr ähnliche paraloge Systeme) auch in vielen obligat anaeroben Stoffwechselprozessen (anaerobe Atmung, Methanogenese und viele Gärungswege) und ebenso bei der anoxygenen und oxygenen Photosynthese bereits wichtige Rollen übernehmen.
Vor einer Milliarde Jahren überstieg die Sauerstoffkonzentration die Marke von einem Prozent, wodurch sich wenige hundert Millionen Jahre später eine erste Ozonschicht bilden konnte. Der heutige Sauerstoffgehalt von knapp 21 % wurde schließlich vor 350 Millionen Jahren erreicht und blieb seitdem recht stabil.
Experimente und Theorien zur chemischen Evolution
Das Miller-Urey-Experiment
Eine der bekanntesten Hypothesen zur Evolution wurde in den zwanziger Jahren des 20. Jahrhunderts vom russischen Forscher A. I. Oparin und dem britischen J. B. S. Haldane veröffentlicht. Die Theorie besagt, dass die Bedingungen der Erde schon damals bestimmte chemische Reaktionen begünstigten. Aus einfacheren Verbindungen der Atmosphäre und des Meeres sollen organische Verbindungen synthetisiert worden sein. Die nötige Energie wurde durch die sehr intensive UV-Strahlung geliefert, die durch Mangel von O2 und O3 ungehindert in die Atmosphäre eindringen konnte.
1953 wurde vom Chemiker Stanley Miller und von Harold C. Urey durch das Ursuppen-Experiment diese Hypothese überprüft. In dem Versuch beweisen sie, dass in einer - den angenommenen präbiotischen Bedingungen ähnlichen - Umgebung, mittels Zufuhr von Energie (Blitzen), aus anorganischen Verbindungen (Wasser, Ammoniak und Wasserstoff) sowie Methan komplexere organische Verbindungen wie Aminosäuren und niedere Carbon- und Fettsäuren entstehen können. In späteren, meist komplizierter aufgebauten Ursuppenversuchen konnten sowohl alle wesentlichen Bausteine der Lebewesen (Aminosäuren, Lipide, Purine (Nucleotidbasen) und Zucker), als auch die komplizierten organischen Verbindungen Porphyrine und Isoprene erzeugt werden.
Obwohl damit die grundsätzliche Möglichkeit der natürlichen Bildung organischer Moleküle gezeigt war, wird die Bedeutung dieses Resultats für den tatsächlichen Ablauf der Entstehung irdischen Lebens heute vielfach kritisch gesehen. In dem Ursuppen-Experiment wurde damals davon ausgegangen, dass die irdische Atmosphäre chemisch reduzierenden Charakter hatte, was dem damaligen Wissensstand entsprach. Heute wird dagegen oftmals von einer nur schwach reduzierenden oder gar neutralen Atmosphäre zu dieser Zeit ausgegangen, allerdings ist die Frage noch nicht abschließend geklärt und es werden auch lokale chemische Inhomogenitäten der atmosphärischen Verhältnisse diskutiert, etwa in der Umgebung von Vulkanen. Zwar konnte durch spätere Experimente gezeigt werden, dass auch unter solchen veränderten atmosphärischen Bedingungen organische Moleküle entstehen; sogar solche, die beim ursprünglichen Experiment nicht entstanden sind, wobei aber die Ausbeute stark vermindert ist. Deswegen wird oft argumentiert, dass andere Möglichkeiten der Herkunft organischer Moleküle zumindest eine zusätzliche Rolle gespielt haben müssen. Angeführt werden hierbei meist die Entstehung organischer Moleküle im Weltall und deren Transfer zur Erde durch Meteoriten oder auch die Entstehung in der Umgebung sogenannter schwarzer Raucher.
Als ein weiteres Argument gegen den Ursprung der biologischen organischen Moleküle gemäß dem Ursuppenversuch wird oftmals angeführt, dass in diesem Versuch ein Racemat, das heißt ein Gemisch aus linksdrehenden und rechtsdrehenden Aminosäuren, entstand. Alle in lebenden Organismen vorkommenden Aminosäuren sind jedoch rein linksdrehend (siehe Biologische Chiralität). Es müsste also einen natürlichen Prozess geben, der chirale Moleküle bestimmter Händigkeit bevorzugt selektiert. Von Exobiologen wird darauf hingewiesen, dass dies einfacher im Weltall zu erklären sei, da photochemische Prozesse mit zirkular polarisierter Strahlung, wie sie beispielsweise durch Pulsare erzeugt wird, in der Lage sind, nur chirale Moleküle bestimmter Händigkeit zu zerstören. Tatsächlich wurden in Meteoriten chirale organische Moleküle gefunden, bei denen die linksdrehende Händigkeit um bis zu 9 % überwog. Allerdings wurde 2001 durch Alan Saghatelian et al.[1] gezeigt, dass auch selbstreplizierende Peptidsysteme in der Lage sind, effektiv homochirale Produkte aus einem ursprünglichen Racemat zu verstärken, was nach Ansicht dieser Forscher die Auffassung der irdischen Entstehung der Händigkeit biologischer Moleküle unterstützt.
Weitere Reaktionen
Aus den beim Miller-Urey-Experiment auftretenden Zwischenprodukten Aldehyde und Cyanwasserstoff lassen sich unter den simulierten Bedingungen der Erde vor 4,5 Milliarden Jahren weitere Biomoleküle herstellen. So gelang Juan Oro 1961 die Synthese von Adenin:
Edukte Produkte 2 Formaldehyd HCN H2O Serin 5 Formaldehyd Ribose 5 HCN Adenin Aus Ribose, Adenin und Triphosphat entsteht das Adenosintriphosphat (ATP), welches in den Organismen als universeller Energieträger und als Baustein (als Monophosphat) der Ribonukleinsäuren (RNA) genutzt wird.
Mitwirkung von Mineralen und Gesteinen
- In winzigen Hohlräumen der Gesteine sind die organischen Moleküle vor UV-Strahlung geschützt.
- Kristalloberflächen können als Matrix für wachsende Makromoleküle dienen. Dabei können die Kristalloberflächen bestimmte Molekülformen bevorzugen. L- und D-Aminosäuren werden auf einem Calcit-Kristall an unterschiedlichen Stellen angelagert.
- Aharon Katchalssky (Weizmann-Institut Israel) konnte in wässriger Lösung mit Hilfe des Tonminerals Montmorillonit Proteine mit einer Kettenlänge von mehr als 50 Aminosäuren in nahezu 100-prozentiger Ausbeute erzeugen.
- Metallionen können als Katalysatoren oder als Elektronendonatoren fungieren oder in Biomoleküle eingebaut werden.
- Tonminerale weisen oft eine elektrische Ladung auf und können so geladene organische Moleküle anziehen und festhalten.
Die Eisen-Schwefel-Welt (ESW) nach Wächtershäuser
Eine besonders intensive Form der Mitwirkung von Mineralen und Gesteinen bei der präbiotischen Synthese organischer Moleküle muss sich auf der Oberfläche von Eisensulfid-Mineralen abgespielt haben. Die Theorie von Miller-Urey weist nämlich schwerwiegende Begrenzungen auf, besonders in Hinblick auf das Fehlen einer Erklärung für die Polymerisation der gebildeten monomeren Bausteine der Biomoleküle.
Ein alternatives Szenario für die frühe Evolution des Lebens wurde daher seit Anfang der 1980er Jahre von dem Münchner Patentanwalt Günter Wächtershäuser entwickelt. Danach wäre das Leben auf der Erde an der Oberfläche von Eisen-Schwefel-Mineralen entstanden, also Sulfiden, die sich heute noch durch geologische Prozesse an Tiefsee-Vulkanen bilden, zur Frühzeit der Erde noch wesentlich häufiger aufgetreten und wohl auch auf vielen Exoplaneten und Planemos vorhanden sein müssen. Der große Vorteil dieses Konzepts gegenüber allen anderen Theorien ist, dass damit erstmals die Möglichkeit besteht, die Bildung von komplexen Bio-Molekülen an eine kontinuierlich verfügbare und verlässliche Energieversorgung zu koppeln. Die Energie kommt durch die Reduktion von teilweise oxidierten Eisen-Schwefel-Mineralen wie Pyrit (FeS2) mit elementarem Wasserstoff (H2) zustande (Reaktionsschema: FeS2 + H2
 FeS + H2S) und liefert genug Energie, um eine präbiotische Ammoniaksynthese und auch um endergone Synthesereaktionen für monomere Bausteine von Biomolekülen und für deren Polymerisierung anzutreiben. Ähnlich wie Eisen-Ionen bilden auch andere Schwermetall-Ionen mit Schwefelwasserstoff unlösliche Sulfide (siehe Schwefelwasserstoffgruppe).
FeS + H2S) und liefert genug Energie, um eine präbiotische Ammoniaksynthese und auch um endergone Synthesereaktionen für monomere Bausteine von Biomolekülen und für deren Polymerisierung anzutreiben. Ähnlich wie Eisen-Ionen bilden auch andere Schwermetall-Ionen mit Schwefelwasserstoff unlösliche Sulfide (siehe Schwefelwasserstoffgruppe).Zusätzlich bieten Pyrit und andere Eisen-Schwefel-Minerale positiv geladene Oberflächen, an denen sich die überwiegend negativ geladenen Biomoleküle (organische Säuren, Phosphatester, Thiolate) anlagern (oft durch Komplexbildungsreaktionen), konzentrieren und miteinander reagieren können. Hierfür erforderliche Stoffe wie Schwefelwasserstoff, Kohlenmonoxid und Eisen-II-Salze gelangen aus der Lösung zudem direkt an die Oberfläche dieser "Eisen-Schwefel-Welt" (ESW). Wächtershäuser zieht für seine Theorie die heute noch bestehenden Grundmechanismen des Stoffwechsels heran und leitet aus diesen ein in sich konsistentes Szenario der Synthese von komplexen organischen Molekülen und Biomolekülen (organische Säuren, Aminosäuren, Zucker, Nucleinbasen, Lipide) aus einfachen anorganischen Vorläufer-Molekülen ab, die in vulkanischen Gasen zu finden sind (NH3, H2, CO, CO2, CH4, H2S).
Im Gegensatz zur Miller-Urey-Hypothese wird keine Kraft von außen in Form von Blitzen oder UV-Strahlung benötigt; zudem laufen die ersten einfachen chemischen Umsetzungen bei erhöhter Temperatur viel schneller ab, ohne dadurch (wie etwa enzym-katalysierte biochemische Reaktionen) behindert zu werden. Da bei Tiefsee-Vulkanen Temperaturen bis 350 °C erreicht werden, kann man sich die Entstehung des Lebens bei diesen hohen Temperaturen gut vorstellen. Erst später, nach der Entstehung temperatursensitiver Katalysatoren (Vitamine, Proteine), muss sich die weitere Evolution in kühleren Nischen vollzogen haben.
Das Wächtershäuser-Szenario passt deshalb auch gut zu den beobachteten Verhältnissen in Schwarzen Rauchern der Tiefsee, weil in diesen Strukturen durch steile Temperaturgradienten von innen nach außen eine solche Einnischung leicht möglich erscheint. Auch heute noch ist nachweisbar, dass die gegenwärtig lebenden Mikroorganismen, die als die ursprünglichsten angesehen werden, zugleich die thermophilsten sind mit einem (bisherigen) Temperatur-Maximum für das Wachstum bei +113 °C. Außerdem sind auch heute noch in der Biochemie Eisen-Schwefel-Zentren als aktive Zentren in vielen wichtigen Enzymen beteiligt, was auf die ursprüngliche Beteiligung von Fe-S-Mineralen an der Evolution des Lebens hindeuten könnte, zumal diese auch heute noch Stoffwechselprodukt anaerober Bakterien darstellen. (siehe folgende Abbildung, Mitte:)

Eisen und Schwefel reagieren zu Eisensulfid
.jpg)
Anaerobe Bakterien, deren archaischer Stoffwechsel Eisen und Schwefel einbezieht, existieren noch heute - Stoffwechselprodukt: Eisensulfid.

Handstück mit ineinander verwachsenen Eisensulfid-Kristallen FeS2
Die Bildung von Makromolekülen
Biomakromoleküle sind Proteine und Nukleinsäuren. Die Verlängerung der Molekülketten (Polymerisation) benötigt Energie und erfolgt unter Wasserabspaltung (Kondensation). Umgekehrt liefert die Spaltung der Makromoleküle (Hydrolyse) Energie. Da das chemische Gleichgewicht soweit auf der Seite der Monomere liegt, dass diese Reaktionen thermodynamisch irreversibel in Richtung der Polymer-Hydrolyse verlaufen, kann es keine Polymer-Synthese ohne ein zugeschaltenes Energie-lieferndes System geben. Auch durch theoretische Hilfskonstrukte, wie Abdampfen von Wasser, Zugabe von Salz (entzieht Wasser) oder Ausfällung der Produkte, ändert sich dieses Problem nur unwesentlich. Die Entstehung von Leben ist also sehr wahrscheinlich an die Kopplung an eine verlässliche Energiequelle gebunden, die für die Polymersynthese genutzt werden kann.
aber
Als Energiequelle wird heute in der Biochemie vor allem ATP genutzt, dessen Bildung allerdings bereits die Anwesenheit von Enzymen voraussetzt. Unter den Bedingungen der Proto-Erde kann man sich andererseits auch die Energetisierung der Polymersynthese durch hydrolytische Spaltung von Polyphosphat vorstellen, das auch heute noch von einigen Enzymen anstelle von ATP genutzt wird. Allerdings ist es auch bei Polyphosphaten schwierig vorstellbar, dass diese in den notwendigen Mengen zur Verfügung standen, da sie zwar spontan gebildet werden können, wenn phosphathaltige Lösungen eingedampft werden, aber auch relativ schnell wieder spontan hydrolysieren, wenn sie wieder in Lösung kommen. Aus diesen Überlegungen heraus müsste man als Entstehungsort des Lebens eine seichte Meeresbucht fordern, die regelmäßig austrocknet und wieder überflutet wird. Dadurch würden aber auch alle Wasser-abhängigen Prozesse immer wieder unterbrochen und die Lebensentstehung zumindest stark verzögert. Schließlich kann man sich aber auch ein ganz anderes System vorstellen, in dem sowohl die Synthese der Bausteine als auch die Energie-abhängige Bildung von Polymeren als kontinuierlicher Prozess in ununterbrochener Kopplung an eine verlässliche Energiequelle stattfindet, nämlich anaerobe Redoxreaktionen mit Metallsulfiden. Diese werden durch vulkanische Tätigkeit auch heute noch in großen Mengen am Grund der Ozeane freigesetzt, wo sie unter anderem Strukturen wie die Black Smoker ausbilden, die von diversen Mikroorganismen und höheren Tieren dicht bevölkert werden.
Das Gleichgewicht der Polymersynthese wird durch Erhöhung der Konzentration der Bausteine (Monomere) und durch Entwässerung der Produkte zugunsten der Bildung der Polymere verschoben. Voraussetzung dafür ist eine Kompartimentierung, das heißt die Abgrenzung von Reaktionsräumen voneinander, die nur noch in begrenztem Stoffaustausch zur Umwelt stehen. In der konventionellen Theorie wurde dies in flachen, kleinen Gewässern (Tümpeln) mit hoher Verdunstungsrate angesiedelt, was als Grundidee noch auf Charles Darwin zurückgeht. Allerdings werden auch heute noch in vulkanischen Gebieten der Tiefsee in den großen Strukturen von Schwarzen Rauchern ausgefällte Metallsulfide in Form von kleinen Kavernen beobachtet, die ebenfalls ein attraktives Szenario einer Umwelt darstellen, wo alle Reaktionen von der Monomersynthese über die Aufkonzentrierung und Polymerisation der Monomere mit einem „eingebauten“ Energiekonservierungssystem ablaufen können.
Weitere Lösungsansätze wurden in Betracht gezogen, haben aber alle schwerwiegende Limitationen und sind nicht gut mit den Bedingungen auf der frühen Erde in Einklang zu bringen. Meist wird dabei für einen oder mehrere Schritte Wasserausschluss gefordert, der im Chemielabor einfach zu erreichen ist, schwerer aber auf der Proto-Erde. Eines dieser Systeme ist die Polymerisation von Carbodiimiden (R-N=C=N-R) oder Dicyan (N≡C-C≡N) in wasserfreiem Medium. Hier wird die Kondensation der Bausteine mit der Reaktion von Carbodiimid gekoppelt, wobei die notwendige Energie entsteht:
(H-X-OH = Monomeres, zum Beispiel Aminosäure oder Ribose)
(wenn R = H entsteht hier Harnstoff)
Zwar bildet sich Dicyan unter UV-Einwirkung aus Cyanwasserstoff, in austrocknenden Tümpeln würde allerdings auch das leichtflüchtige Molekül verloren gehen.
Wird ein trockenes Gemisch von Aminosäuren einige Stunden auf 130 °C erhitzt, bilden sich proteinähnliche Makromoleküle. Sind Polyphosphate zugegen, genügen 60 °C. Diese Bedingungen können sich ergeben, wenn Wasser mit gelösten Aminosäuren in Kontakt mit heißer, vulkanischer Asche kommt.
Erhitzt man ein Nukleotid-Gemisch in Gegenwart von Polyphosphaten auf 55 °C, entstehen zwar Polynukleotide, allerdings findet die Verknüpfung eher über die 5’- und 2’-C-Atome der Ribose statt, da sie leichter erfolgt als die in allen Organismen vorhandene 5’-3’-Verknüpfung. Aus beiden Typen von Polynukleotiden bilden sich Doppelhelices (vergleiche Bau der DNA). Allerdings ist die 5’-3’-Doppelhelix stabiler als die 5’-2’-Helix.
Fehlt am 2’-C-Atom der Ribose die Hydroxyl-Gruppe, liegt die Desoxyribose vor. Jetzt können sich nur 5’-3’-Verknüpfungen bilden, wie sie für die DNA typisch sind.
Bildung präbiotischer Strukturen (Zellvorläufer)
Zellen erhalten ihre Funktion dadurch aufrecht, dass sie verschiedene Reaktionsräume (Kompartimente) bilden, um die Stoffwechselvorgänge voneinander zu trennen und unerwünschte Reaktion auszuschließen. Gleichzeitig können Konzentrationsgradienten aufgebaut werden.
Koazervate
Vor allem Alexander Iwanowitsch Oparin (1894–1980) beschäftigte sich mit der Möglichkeit des Stoffwechsels in Koazervaten (von lateinisch coacervatio: Aufhäufung). Er konnte zeigen, dass abgegrenzte Räume mit einem einfachen Stoffwechsel prinzipiell durch Selbstorganisation entstehen können, vorausgesetzt, dass Katalysatoren mit spezifischen Eigenschaften vorhanden sind. Da die verwendeten Substanzen dem Repertoire der heute lebenden Organismen entstammen, sind Oparins Koazervate nicht als Vorläufer von Zellen zu sehen, sondern als Analogie-Modelle für die Entstehung von Vorläufer-Zellen.
Gibt man zu kolloidalen Lösungen von Biomakromolekülen Salz hinzu, bilden sich kleine Tröpfchen mit einem Durchmesser zwischen 1 und 500 µm, die die Polymere in hoher Konzentration enthalten.
Oparin untersuchte Gemische aus Eiweißen (Histon und Albumin), Eiweißen und Kohlenhydraten (Histon und Gummi arabicum) und Eiweißen und Polynukleotiden (Histon oder Clupein und DNA oder RNA).
Enthalten Tröpfchen aus Histon und Gummi arabicum das Enzym Phosphorylase, so können diese Tröpfchen Glucose-1-phosphat aus der Umgebung aufnehmen, zu Stärke umwandeln und speichern. Das freigesetzte Phosphat diffundiert nach außen. Das Tröpfchen wird durch die Zunahme an Stärke größer, bis es in kleinere Tröpfchen zerfällt, die wieder Phosphorylase enthalten können, allerdings weniger als das Ausgangströpfchen. Damit verlangsamt sich auch der Stoffwechsel. Hier wird deutlich, dass zu einem Erhalt der Eigenschaften einer Zelle die Regeneration der Enzymausstattung nach erfolgter Teilung notwendig ist.
Wird auch das Stärke abbauende Enzym Amylase hinzugegeben, ergeben sich Koazervate mit einem zweistufigen Stoffwechsel:
- Aufnahme von Glucose-1-phosphat → Aufbau von Stärke.
- Abgabe von Phosphat → Spaltung der Stärke und Abgabe von Maltose.
Mikrosphären
Im Jahr 1970 konnte Sydney Fox nachweisen, dass sich aus den proteinartigen Produkten, die beim Erwärmen trockener Aminosäuregemische entstehen (siehe oben), auch durch Selbstaggregation wachsende Tröpfchen ergeben können, sogenannte Mikrosphären. Sie grenzen sich von der Umgebung durch eine semipermeable Membran ab und nehmen weiteres proteinartiges Material aus der Umgebung auf. Dadurch wachsen sie weiter und zerteilen sich wieder in kleinere Tröpfchen. Des Weiteren fand Fox, dass diese Systeme enzymatische Eigenschaften haben, Glucose abbauen oder sich wie Esterasen oder Peroxidasen verhalten, ohne dass von außen Enzyme hinzugefügt worden wären.
Die RNA-Welt
Die RNA-Welt-Hypothese wurde erstmals 1986 von Walter Gilbert vorgeschlagen und besagt, dass RNA-Moleküle die Vorläufer der Organismen waren. Die Hypothese lässt sich ableiten aus der Fähigkeit der RNA zur Speicherung, Übertragung und Vervielfältigung genetischer Informationen, sowie aus ihrer Fähigkeit, als Ribozyme Reaktionen zu katalysieren. In einer Evolutionsumgebung würden diejenigen RNA-Moleküle gehäuft vorkommen, die sich selbst bevorzugt vermehren.
RNA wird aufgrund diverser Eigenschaften für älter gehalten als DNA. Unter anderem wird Ribose im Gegensatz zu Desoxyribose leicht durch Aldolkondensation gebildet. Auch startet selbst die heutige DNA-Replikation mit RNA-Synthese. Ausgangspunkt der RNA-Evolution sind einfache, sich selbst replizierende RNA-Moleküle. Einige davon erhalten die Eigenschaft, die Synthese von Proteinen zu katalysieren, die selbst wieder die Synthese der RNA und ihre eigene Synthese katalysieren (Entwicklung der Translation). Einige RNA-Moleküle lagern sich zu doppelsträngigen RNA-Molekülen zusammen, die sich zu DNA-Molekülen und Trägern der Erbinformation weiterentwickeln. (Entwicklung der Transkription)
Als Grundlage dienen bestimmte RNA-Moleküle, die von beliebigen RNA-Vorlagen, und damit von sich selbst, Kopien erzeugen können. Jennifer A. Doudna und Jack W. Szostak benutzten als Vorlage zur Entwicklung dieses RNA-Typs das selbst-spleißende Intron des eukaryotischen Einzellers Tetrahymena thermophila. Damit besteht die Möglichkeit, dass in den Ribosomen die eigentlich katalytischen Moleküle die rRNA sind und somit RNA die Eiweißsynthese katalysiert. Einschränkungen bestehen allerdings darin, dass bei der selbstreplizierenden RNA als Bausteine nicht Mononukleotide sondern Oligonukleotide und Hilfsstoffe benötigt werden. 2009 identifizierten Lincoln und Joyce allerdings ein Ribozym, dass sich aus Mononukleotiden repliziert.[2]
2001 wurde entdeckt, dass die wichtigen katalytischen Zentren der Ribosomen von RNA und nicht, wie vorher angenommen, von Proteinen gestellt werden. Dies zeigt, dass eine katalytische Funktion der RNA, wie sie in der RNA-Welt-Hypothese vorgeschlagen wurde, heute von Lebewesen genutzt wird. Da Ribosomen als sehr ursprüngliche Zellbausteine gelten, gilt diese Entdeckung als wichtiger Beitrag zur Untermauerung der RNA-Welt-Hypothese. Man ist nun sicher, dass RNA-Moleküle – zumindest prinzipiell – in der Lage sind, Aminosäuren zu Proteinen zu verketten. In diesem Zusammenhang ist auch die PNA (Peptid-Nukleinsäure) als mögliches Vorläufermolekül der RNA von Interesse.
Siehe auch: Quasispezies, Manfred Eigen
Alternativ betrachtete Möglichkeiten
Biomoleküle aus dem Weltall
Die Erde ist seit Anbeginn ihrer Existenz dem Bombardement von Kometen und Meteoriten ausgesetzt, insbesondere in der als Großes Bombardement (Late Heavy Bombardment) bezeichneten ersten Phase nach der Erdentstehung. In etlichen Meteoriten wurden einfache organische Moleküle nachgewiesen, unter anderem Aminosäuren. Beachtet man die Homochiralität irdischer Biomoleküle (L-Aminosäuren und D-Zucker), so wäre eventuell eine mögliche Erklärung, dass die Aminosäuren aus dem Weltall stammen, da bei einigen dieser meteoritischen Aminosäuren ein Überschuss des L-Typ um bis zu über 9 % nachgewiesen wurde.[3] Diese Verteilung ist allerdings auch durch anorganische Feststoff-Katalysatoren auf der Erde erklärbar. Durch Versuche, in denen Weltraumbedingungen simuliert wurden, konnte inzwischen nachgewiesen werden, dass grundlegende organische Biomoleküle, insbesondere Aminosäuren, auch unter solchen Bedingungen entstehen können.[4]
Black Smoker
An diesen hydrothermalen Quellen findet sich eine möglicherweise günstige Umgebung für die chemoautotrophe Entstehung des Lebens. Diese These wird insbesondere von den Forschern William Martin und Michael Russell vertreten.[5]
Weitere Sichtweisen
Panspermie
Die Panspermie-Hypothese vertritt im Wesentlichen die Meinung, dass durch 'Animpfen' der Erde mit niederen, bakterienähnlichen Lebensformen aus dem Weltall die ersten Lebewesen auf die Erde kamen.
Die Hypothese, dass niedrigere Lebensformen aus dem Weltall auf die Erde gelangt sind, verschiebt das Problem der Entstehung von Leben aber nur auf einen anderen Ort und ist keine Erklärung für die Entstehung des Lebens.
Religiöse Sichtweisen
Die jüdische und christliche Entstehungsgeschichte steht im ersten Buch Mose („Genesis“). Hier wird die Welt durch Gott in 6 Tagen geschaffen. Die meisten modernen Theologen deuten diesen Text symbolisch und sehen keinen Widerspruch zur Naturwissenschaft. Dies ist auch Position der lutherischen und der katholischen Kirche in Deutschland. Im Gegensatz dazu stehen Kreationisten, die die Genesis wörtlich nehmen.
Resümee
Bei aller Unsicherheit über die konkrete Entstehung von Leben auf der Erde scheint sicher zu sein, dass sich nur eine Form von Leben, nämlich die auf Nukleinsäuren (RNA und DNA) beruhende, durchgesetzt hat (falls es je mehrere verschiedene gegeben haben sollte). Das wesentliche Indiz für diese Theorie besteht in der Gleichheit der Bausteine der zwei wesentlichsten lebenstypischen Makromoleküle (Nukleinsäuren und Proteine), die fünf Nukleotide und die 20 Aminosäuren in allen bekannten Lebensformen, insbesondere aber auch der universell gültige genetische Code.
Man nimmt auch an, dass das Entstehen neuen Lebens auf unserer Erde jetzt nicht mehr möglich ist, da alle Ökosysteme bereits von Leben besetzt sind und die Entstehung neuen Lebens unter dem Konkurrenzdruck anderer Lebewesen unmöglich ist.
Siehe dazu auch: Exobiologie
Literatur
- Iris Fry: The Emergence of Life on Earth: A Historical and Scientific Overview. Rutgers University Press, 2000, ISBN 0-8135-2740-6.
- Sven P.Thoms: Ursprung des Lebens. S. Fischer Verlag GmbH, Frankfurt am Main 2005, ISBN 3-596-16128-2.
- Harald Lesch, Jörn Müller: Big Bang, zweiter Akt. Auf den Spuren des Lebens im All. S. 29-84, 124-175, 409. Wilhelm Goldmann Verlag, München 2005, ISBN 3-442-15343-3.
- Michael Wächter: Stoffe, Teilchen, Reaktionen. S. 133-138. Verlag Handwerk und Technik, Hamburg 2000, ISBN 3-582-01235-2.
- Leslie E. Orgel: Der Ursprung des Lebens. In: Spektrum der Wissenschaft. Spezial 3: Leben und Kosmos. Verlag Spektrum der Wissenschaft, Heidelberg 1995.
- Horst Rauchfuß: Chemische Evolution und der Ursprung des Lebens. Springer-Verlag, Heidelberg, Berlin 2005, ISBN 3-540-23965-0.
- Ute Kehse: Das Wunder in der Tiefsee. In: GEO kompakt. Nr. 1, S. 108-117. Gruner + Jahr, Hamburg 2004, ISBN 3-570-19570-8, ISSN 1614-6913.
- Robert Shapiro: Ein einfacher Ursprung des Lebens. In: Spektrum der Wissenschaft. November 2007, S. 64–72. ISSN 0170-2971.
- Uwe Meierhenrich: Amino Acids and the Asymmetry of Life. Springer-Verlag, Heidelberg, Berlin 2008, ISBN 978-3-540-76885-2.
Quellen
- ↑ A. Saghatelian et al.: A chiroselective peptide replicator. In: Nature. Bd. 409, S. 797-801. 2001.
- ↑ Lincoln, Tracey; Joyce, Gerald F. (January 8, 2009): "Self-Sustained Replication of an RNA Enzyme". Science (New York: American Association for the Advancement of Science). doi:10.1126/science.1167856. ISSN 1095-9203. PMID 19131595, http://www.sciencemag.org/cgi/content/abstract/1167856, abgerufen am 13. Januar 2009
- ↑ [1] J.R. Cronin, S. Pizzarello: Enantiomeric Excesses in Meteoritic Amino Acids. In: Science. Bd. 275, S. 951-955. 2002
- ↑ M.P. Bernstein, et al.: Racemic amino acids from the ultraviolet photolysis of interstellar ice analogues. In: Nature. Bd. 416, 401. 2002. und G. Munoz Caro, et al.: "Amino acids from ultraviolet irradiation of interstellar ice analogues". In: Nature. Bd. 416, 403. 2002.
- ↑ W. Martin, M.J. Russell: On the origins of cells: a hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleared cells. In: Philos. Trans. R. Soc. London Ser. Bd. 358, S. 59–85. 2003.
Siehe auch
Weblinks
- Wie dünn war die Ursuppe?, Flash-Video aus der Fernsehsendung alpha-Centauri (JavaScript benötigt)
- Außerirdischer Ursprung der Homochiralität
- Audiomitschnitt: Ein höllischer Beginn - Die Entstehung des Lebens in einer vulkanischen Eisen-Schwefel-Welt Von Günter Wächtershäuser. Öffentliche Ringvorlesung: Evolution - Zufall und Zwangsläufigkeit der Schöpfung. Universität Göttingen, Wintersemester 2007/08. MP3-Datei, ca. 38 MB.





.jpg)

![\mathrm{ [Monomere]_n + (n-1) \, H_2O \rightarrow n \, Monomere \, + W\ddot arme}](/pictures/dewiki/49/12465d90f3dbefe3a779478e2ae7affd.png)
![\mathrm{Energie + n\, Monomere \, \rightarrow [Monomere]_n + (n-1)\, H_2O}](/pictures/dewiki/101/e27c7a9a5b81a204fceb4e3e1163cd51.png)
![\mathrm{[H-X-OH] + [H-X-OH] + Energie \ \leftrightharpoons \ [X-X] + H_2O}](/pictures/dewiki/48/06e7b38e4adc5cab03ef4e803fef7c89.png)
![\mathrm{[R-N \equiv C \equiv N-R] + H_2O \ \rightarrow \ [R-NH-CO-NH-R] + Energie}](/pictures/dewiki/51/31e89f0ea2f6c8b63f82542c2de3090e.png)
Wikimedia Foundation.