- Eutardigrada
-
Eutardigrada 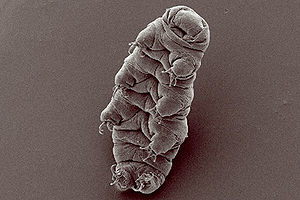
Das Eutardigrada-Bärtierchen Hypsibius dujardini
Systematik Abteilung: Gewebetiere (Eumetazoa) Unterabteilung: Bilateria ohne Rang: Urmünder (Protostomia) Überstamm: Häutungstiere (Ecdysozoa) Stamm: Bärtierchen (Tardigrada) Klasse: Eutardigrada Wissenschaftlicher Name Eutardigrada Marcus 1929 Ordnungen - Parachela
- Apochela
Als Eutardigrada bezeichnet man eine Klasse von Bärtierchen (Tardigrada), denen im Gegensatz zur anderen Bärtierchen-Klasse, den Heterotardigrada, auffällige Merkmale wie Kopfanhänge oder verhärtete Rückenpanzer fehlen. Sie verfügen dafür über spezielle Ausscheidungsorgane, die Malpighischen Drüsen.
Das 1929 durch den deutschen Zoologen Ernst Marcus erstmals beschriebene Taxon umfasst mehrheitlich süßwasser- und landlebende (limno-terrestrische) Formen; einige Arten finden sich allerdings auch im Salzwasser.
Inhaltsverzeichnis
Merkmale
Bärtierchen stellen eine insgesamt sehr homogene Gruppe da, deren Arten sich anatomisch nur in Details unterscheiden. Im folgenden werden daher nur die für die Eutardigrada besonders charakteristischen Eigenheiten beschrieben, eine ausführlichere Einführung in den Körperbau der Tiere findet sich im Hauptartikel zu den Bärtierchen. Die durchschnittliche Körpergröße der Tiere liegt wie bei allen Bärtierchen bei etwa 100 bis 150 Mikrometern, eine Art aus der Gattung Macrobiotus wird dagegen bis zu 1,5 Millimeter lang.
Cuticula
Die nicht-zellige Außenhaut (Cuticula) der Eutardigrada ist sehr oft vollkommen glatt, gelegentlich granulär, also von körnigem Aussehen, und manchmal auch mit kleinen Höckern, den Tuberkeln, bedeckt. Sie besitzt aber nie auffällige Verhärtungen wie die Skleriten genannten Panzerplatten, die sich auf dem Rücken der Heterotardigrada finden; auch dornige Cuticula-Fortsätze sind bei weitem nicht so deutlich ausgeprägt wie in jener Gruppe. Dafür ist die Cuticula sehr oft durch Pigmente bunt gefärbt.
Kopf- und Körperanhänge
Lange Kopfanhänge wie die fädigen Cirri der Heterotardigrada besitzen die Eutardigrada nicht, allerdings finden sich um die Mundöffnung herum angeordnet oft noppenförmige Erhebungen, die Papillen oder Papulae und lappige Strukturen. Die grundlegende Beinanatomie entspricht der aller Bärtierchen; allerdings befinden sich die Klauen immer direkt am Beinende und sind niemals in Haftscheiben umgewandelt. Sie bestehen immer aus einem Doppelklauenpaar, wobei jede Doppelklaue aus einer gemeinsamen Basis, dem Klauenansatz und zwei Armen, dem primären und dem sekundären Arm besteht, deren genaue Form und Anordnung wichtige Merkmale bei der Art-Identifikation sind. Um den Klauenansatz herum findet sich oft eine aus Cuticula bestehende Verdickung, die man als Lunulus bezeichnet.
Leibeshöhle und Verdauungstrakt
Die fachsprachlich Hämocoelom genannte Leibeshöhle fällt bei den Eutardigrada meist etwas geräumiger aus; sie wird wie bei allen Bärtierchen vom Verdauungstrakt durchzogen. Dieser beginnt mit der Mundhöhle, die sich bei manchen Arten in einen starren vorderen und einen elastischen hinteren Teil gliedert. Die Grenze zwischen beiden Regionen liegt etwa auf Höhe der Stiletthalter, welche die beiden seitlich der Mundhöhle gelegenen, oft kräftigen und manchmal gekrümmten Stilette befestigen. Zum Vorderende der Mundhöhle hin findet man oft zwei Bänder mit kleinen „Zähnchen“, den Mucrones, hinter denen sowohl rück- als auch bauchseitig quer zur Körperlängsachse (transversal) verlaufende Erhebungen auftreten können, die ebenfalls Erkennungsmerkmale bei der Artbestimmung sind. Gelegentlich wird die Mundhöhle durch eine mittig auf der Bauchseite verlaufende Leiste verstärkt.
Der muskulöse Schlund ist je nach Taxon entweder kugel-, ei- oder birnenförmig geformt und durch Placoiden verstärkt: Darunter versteht man kurze Streifen aus verhärteter Cuticula, die gewöhnlich in drei Paar Längsreihen vom Vorderende des Schlundes zum Hinterende verlaufen. Anders als bei den Heterotardigrada sind diese Längsstreifen nicht durchgehend, sondern in regelmäßigen Abständen unterbrochen. Die ersten drei Placoiden einer Reihe sind meist größer als die folgenden und werden als Macroplacoiden bezeichnet, dahinter befinden sich meist noch ein bis zwei kleinere Microplacoiden. Der Darm verläuft immer glatt und besitzt keine Diverticula genannten Ausstülpungen.
Malpighische Drüsen
Eine bemerkenswerte Eigenheit der Eutardigrada sind die Malpighischen Drüsen, die normalerweise der Ausscheidung dienen. Die meisten Arten haben sowohl eine rückseitige (dorsale) als auch zwei seitlich angeordnete (laterale) Drüsen, die an der Verbindungsstelle von Mittel- und Hinterdarm münden. Sie bestehen aus drei bis zwölf Zellen und teilen sich meist in ein distales (darmfernes) und ein proximales (darmnahes) Segment auf. Beide Segmente haben oft die gleiche Zellenzahl, wobei 3+3 und 6+6 häufig zu findende Kombinationen darstellen. Das Distalsegment hat die Aufgabe, den Primärurin zu produzieren; wahrscheinlich geschieht dies durch aktiven Stofftransport über die trennende Membran hinweg, da es keine Hinweise auf eine Ultrafiltration der Leibeshöhlenflüssigkeit gibt. Das Proximalsegment modifiziert dann den Primärurin, in dem selektiv Wasser oder Nährstoffe wieder aufgenommen und dafür weitere Schad- oder Abfallstoffe abgegeben werden.
Eine Besonderheit findet man bei der meereslebenden Gattung Halobiotus, deren Malpighische Drüsen stark vergrößert sind. Sie ist vermutlich sekundär ins Meer zurückgekehrt und nutzt ihre Drüsen wahrscheinlich zur Regulation des Salzhaushalts (Osmoregulation). Auch limno-terrestrische Arten sind darauf angewiesen, ihre Wasseraufnahme aktiv zu kontrollieren, da ihre Körperflüssigkeit anisotonisch zu Süßwasser ist, also nicht denselben Salzgehalt aufweist. Die weitverbreitete Art Milnesium tardigradum besitzt vermutlich eine weitere Malpighische Drüse, die bauchseitig gelegen ist und anscheinend eine Rolle bei der geschlechtlichen Fortpflanzung spielt; dies wird gelegentlich als Beispiel für Exaptation gewertet, den Gebrauch einer ursprünglich aus einem anderen Grund entstandenen Struktur für eine neue Funktion.
Fortpflanzungsorgane
Im Gegensatz zu den Heterotardigrada besitzen die Weibchen der Eutardigrada keine separate Geschlechtsöffnung. Stattdessen mündet der Eileiter in den Hinterdarm, der sich somit als Kloake bezeichnen lässt. Diese öffnet sich in einem quer zur Körperachse (transversal) verlaufenden Schlitz vor dem vierten Beinpaar zur Außenwelt.
Manche Weibchen besitzen einen unpaarigen Samenspeicher, in dem nach einer Kopulation das Sperma der Männchen aufbewahrt wird.
Verbreitung und Lebensraum
Eutardigrada finden sich weltweit auf allen Kontinenten und sind auch im Meer weit verbreitet, obwohl sie in diesen Lebensraum wahrscheinlich nur sekundär zurückgekehrt sind. Besonders häufig findet man sie aber an Land, insbesondere in der Laubstreu, den obersten Zentimetern des Waldbodens oder in Moosen (Bryata). Unterschiedliche Arten bevorzugen unterschiedliche Lebensräume innerhalb eines Mooskissens; als Folge lassen sich in einem Stück Moos oft in verschiedenen Zonen unterschiedliche Arten auffinden.
Lebensweise
Die Lebensweise der Eutardigrada unterscheidet sich nicht allzu sehr von jener der Heterotardigrada, so dass an dieser Stelle für die Grundlagen erneut auf den Hauptartikel zu den Bärtierchen verwiesen wird. Parasitismus ist bei den Eutardigrada nicht verbreitet, was angesichts ihres hauptsächlich terrestrischen Lebensraums nicht Wunder nimmt: Eutardigrada sind wie alle Bärtierchen im aktiven Zustand auf einen dünnen Wasserfilm angewiesen, so dass bei Eintritt von Trockenheit entweder die Verbindung mit dem Wirt verloren gehen oder das entsprechende Individuum unkontrolliert austrocknen würde.
Dafür findet sich in der Gruppe sowohl Cyclomorphose, das Auftreten verschiedener Körperformen (Morphe) in unterschiedlichen Jahreszeiten, Cystenbildung und Kryptobiose, die wohl extremste Form der Anpassung an widrige Umweltbedingungen, bei der die betroffenen Individuen in einen todesnahen Zustand ohne Stoffwechselaktivität übergehen, der nach seiner Form als Tönnchen bezeichnet wird. Da sich kryptobiotische Zeiten auch innerhalb der Heterotardigrada finden lassen, ist diese spezielle Anpassung der Bärtierchen im Hauptartikel beschrieben.
Die Fortpflanzung der Eutardigrada kann sowohl auf asexuellem als auch auf geschlechtlichem Wege geschehen. Im ersteren Fall vermehren sich einzelne Weibchen parthenogenetisch, also ohne Beteiligung von Männchen. Man unterscheidet dabei zwischen zwei verschiedenen Formen, die beide innerhalb der Eutardigrada auftreten:
- Bei der meiotischen oder automiktischen Parthenogenese kommt es zwar zur Meiose genannten Reifeteilung, bei der sich in den Vorläuferzellen der Eizellen die Chromosomenzahl halbiert; dieser schließt sich aber vor der normalerweise nachfolgenden zweiten Teilung, einer Mitose, eine Chrosomenduplikation an, so dass nach dem aus Meiose, Duplikation und Mitose bestehenden Gesamtprozess die Chromosomenzahl unverändert geblieben ist. Aus den unbefruchteten Eiern mit doppeltem Chromosomensatz (Diploidie) entwickeln sich dann die Jungtiere.
- Bei der ameiotischen oder apomiktischen Parthenogenese unterscheiden sich Weibchen, die sich geschlechtlich fortpflanzen und solche, die sich ungeschlechtlich fortpflanzen, durch ihre Chromosomenzahl: Letztere sind oft tri-, gelegentlich sogar tetraploid, das heißt, sie besitzen statt zwei Chromosomensätzen drei oder vier und können sich aus diesem Grunde nicht sexuell fortpflanzen; ihre Eier entstehen daher durch einfache mitotische Teilung, bei der die Chromosomenzahl unverändert bleibt.
Bei der sexuellen Fortpflanzung sind fast immer die Geschlechter getrennt, obwohl in zwei Gattungen, Isohypsibius und Amphibolus auch zwittrige (hermaphroditische) Arten bekannt sind, die über eine unpaarige Keimdrüse verfügen, die gleichzeitig als Hoden und Eierstock fungiert. Selbstbefruchtung ist aber mit sehr hoher Wahrscheinlichkeit eine abgeleitete, keine ursprüngliche Eigenschaft der Gruppe.
Sind, wie meist, die Geschlechter, getrennt, so lassen sie sich daran unterscheiden, dass die Männchen eine separate Geschlechtsöffnung, die Gonopore besitzen. Bei erwachsenen Männchen sind zudem oft die Klauen des ersten Beinpaares gegenüber der gleichen Struktur der Weibchen und geschlechtsunreifen Jungtiere modifiziert. Männchen sind häufig auch etwas kleiner als die Weibchen, wobei sich allerdings erwachsene männliche und jüngere weibliche Tiere nicht nach diesem Kriterium auseinanderhalten lassen.
Die Eier der Eutardigrada können entweder glatt oder durch vielfältige Strukturen ornamentiert sein, die alle bei der Artbestimmung herangezogen werden können. Glatte Eier werden meist in der bei der Häutung abgestoßenen alten Außenhaut (Cuticula) abgelegt, die nun als Exuvium bezeichnet wird. Weibchen der Gattungen Pseudobiotus und Isohypsibius schleppen dieses Exuvium oft für eine Weile mit sich herum, was als eine Art primitiver Brutpflege gedeutet werden kann. Eier mit skulptierter Oberfläche werden dagegen meist ohne weitere Schutzhülle freigesetzt. Einige süßwasserlebende Arten aus der Gattung Murrayon legen ihre Eier in die leeren Außenskelette von Wasserflöhen (Cladocera), Ostrakoden (Ostracoda) und Insekten (Insecta) ab.
Die Entwicklung vom geschlüpften Jungtier zum geschlechtsreifen Individuum verläuft immer direkt, ohne zwischengeschaltetes Larvenstadium. Die normale Lebensdauer liegt zwischen mehreren Monaten und ein bis zwei Jahren, kann innerhalb der landlebenden Formen aber durch kryptobiotische Zeiträume, in denen die Tiere nicht altern, auf mehrere Jahre, vereinzelt wohl auch Jahrzehnte, verlängert sein.
Stammesgeschichte
Moderne Formen
Die genauen stammesgeschichtlichen Verwandtschaftsverhältnisse der Eutardigrada zur anderen Klasse der Bärtierchen, den Heterotardigrada, sind noch ungeklärt. Eine häufig vertretene Ansicht geht davon aus, dass die Eutardigrada ihre Wurzeln in einer bereits landlebenden Ordnung der Heterotardigrada haben, den Echiniscoidea und zwar insbesondere der Familie Echiniscidae. Damit würde sich eine Entwicklung von den meereslebenden Arthrotardigrada, der anderen Heterotardigrada-Ordnung über die Echiniscoidea hin zu den Eutardigrada ergeben:
Bärtierchen Heterotardigrada Arthrotardigrada verschiedene Familien
Echiniscoidea verschiedene Familien
Echiniscidae verschiedene Gattungen
Eutardigrada
Vorläufige molekulargenetische Untersuchungen haben diese Hypothese jedoch nicht bestätigen können und sprechen stattdessen für eine unabhängige Entwicklung der Eutardigrada:Bärtierchen Heterotardigrada Arthrotardigrada
Echiniscoidea
Eutardigrada
Es gilt als sehr wahrscheinlich, dass die marinen Arten der Eutardigrada aus der Gattung Halobiotus sekundär ins Meer zurückgekehrt sind. Dafür sprechen insbesondere die stark vergrößerten Malpighischen Drüsen dieser Gattung, die wohl erst sekundär eine herausgehobene Funktion bei der Osmoregulation übernommen haben.Fossile Formen
Eine fossile Art, Beorn leggi, die in kreidezeitlichem Bernstein aus Kanada gefunden wurde, lässt sich bereits in die Eutardigrada einordnen, dazu kommen etwa gleichaltrige Funde aus dem US-amerikanischen Bundesstaat New Jersey.
Davon abgesehen, dass die Gruppe anscheinend seit mehr als 65 Millionen Jahren morphologisch äußerst stabil geblieben ist, wirft keiner der Funde ein Licht auf die Naturgeschichte der Eutardigrada.
Systematik
Die Eutardigrada gelten als monophyletisches Taxon, das heißt, man geht davon aus, dass alle dazugerechneten Arten auf eine gemeinsame Stammart zurückgehen und dass alle Nachkommen dieser Stammart auch in den Eutardigrada enthalten sind.
Man unterscheidet in der Klasse zwei Ordnungen mit insgesamt sechs Familien, die hauptsächlich durch die Feinstruktur der Beinklauen und der Schlundmuskulatur definiert sind:
- Die Parachela sind die größere Gruppe und umfassen sowohl marine als auch limno-terrestrische Formen. Die größere Primärklaue entspringt bei ihnen nicht direkt am Beinansatz sondern an der kleineren Sekundärklaue. Der Schlund hat meist ein kugel- oder eiförmiges Aussehen und besitzt stabilisierende Längsstreifen (Placoiden). Man unterscheidet nach der Klauenstruktur und den Details der Morphologie der Schlundmuskulatur fünf Familien, deren stammesgeschichtliche Verwandtschaftsbeziehungen zueinander noch ungeklärt sind:
- Bei den land- und süßwasserlebenden Macrobiotidae sind die Beinklauen in Bezug auf die Beinmittelebene symmetrisch angeordnet; die primären Klauen sitzen beide innen, die sekundären Klauen beide außen, das Muster wird entsprechend durch die Formel 2-1-1-2 beschrieben.
- Bei den terrestrischen Calophysibiidae sind die beiden kleinen Beinklauenpaare ähnlich geformt, aber in Bezug auf die Beinmittelebene asymmetrisch angeordnet; die primären und sekundären Klauenäste folgen der Folge 2-1-2-1 von außen nach innen. Im Gegensatz zu den meisten anderen Arten besitzen Calophysibiidae-Arten keine Klauenbasis.
- Bei den meereslebenden Hypsibiidae sind die beiden Klauenpaare eines jeden Beins nicht nur asymmetrisch angeordnet, sondern unterscheiden sich auch in Größe und Form. Die Klauenäste alternieren (2-1-2-1).
- Die limno-terrestrischen Eohypsibiidae zeichnen sich durch eine dreigeteilte Klauenmorphologie aus, bei der Klauenbasis, erste und sekundäre Klaue durch Trennwände (Septa) voneinander abgeteilt sind. Inneres und äußeres Klauenpaar sind immer gleich groß, die Reihenfolge von Primär- und Sekundärast wird durch die Formel 2-1-2-1 beschrieben; allerdings kann es vorkommen, dass das innere Klauenpaar um 180 Grad gedreht ist, wodurch sich die symmetrische Reihenfolge 2-1-1-2 ergibt.
- Bei den landlebenden Necopinatidae sind die stark reduzierten Klauen nur auf dem ersten Beinpaar vorhanden.
- Die Microhypsibiidae sind land- und süßwasserlebend; ihre Klauenpaare alternieren (2-1-2-1), ähneln sich aber in Größe und Form, die Klauenbasis ist sehr schmal.
- Eine Gattung, Apodibius konnte bisher keiner der vorgenannten Familien zugeordnet werden.
- Die Apochela bilden die zweite Ordnung und finden sich ausschließlich an Land. Alle Arten werden formell einer einzigen Familie Milnesiidae zugeordnet und sind dadurch gekennzeichnet, dass bei ihnen sowohl der lange, schlanke, primäre als auch der kurze, stämmige, sekundäre Ast jeder Doppelklaue direkt am Beinende entspringt; letzterer besitzt oft zwei zusätzliche Haken. Der Schlund ist bei den Apochela birnenförmig ausgeprägt und lässt keine Placoiden genannten Stützstrukturen erkennen.
Apochela und Parachela unterscheiden sich daneben noch in Details der Sensillenmorphologie.
Literatur
- I. M. Kinchin: The biology of tardigrades. Portland Press, 1994
- D. R. Nelson, N. J. Marley: The biology and ecology of lotic Tardigrada. Freshwater Biology, 44, 2000, Seite 93

Wikimedia Foundation.