- Facettenaugen
-
 Facettenaugen einer Schwebfliege
Facettenaugen einer SchwebfliegeAls Facetten-, Fassetten- oder Komplexauge oder oculi compositi bezeichnet man den häufig bei Gliederfüßern vorkommenden Augentyp, bei dem ein Auge sich aus mehreren, bei bestimmten Insekten wie z. B. den Libellen sogar aus mehreren zehntausend Ommatidien (Einzelaugen) zusammensetzt.
Inhaltsverzeichnis
Aufbau
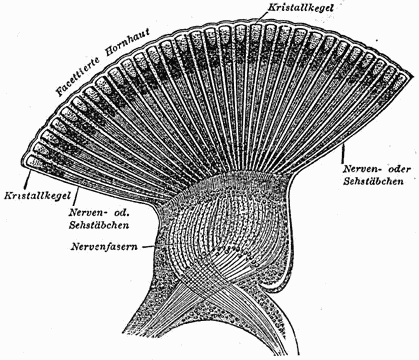
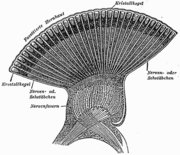 Facettenauge einer Libelle (Schnittzeichnung)
Facettenauge einer Libelle (Schnittzeichnung) Einzelauge (Ommatidium) aus einem Facettenauge (Appositionsauge)
Einzelauge (Ommatidium) aus einem Facettenauge (Appositionsauge)
A Chitinlinse, B Kristallkegel, C Hauptpigmentzelle D Trennwand zur Nachbarommatidie E Rhabdom F Sehzellen G Basalmembran H NervenfaserFacettenaugen sind meistens annähernd halbkugelförmig, was bewirkt, dass jedes ihrer Ommatidien in eine geringfügig andere Richtung blickt. Die Facettenaugen befinden sich an beiden Seiten des Insektenkopfes. Daher werden sie auch manchmal „Seitenaugen“ genannt. Sie sind starr mit der Kopfkapsel verbunden und können nicht, wie die Augen des Menschen, bewegt werden. Abhängig von der Lebensweise sind sie verschieden groß ausgebildet, bei schnell fliegenden und räuberischen Arten machen sie 70 % bis 90 % der Kopffläche aus. Das Insekt kann sich so ein Bild seiner Umgebung aus einzelnen Bildpunkten zusammensetzen.
Die Anzahl von Einzelaugen innerhalb eines Facettenauges unterscheidet sich von Art zu Art, manchmal auch innerhalb der Geschlechter einer Art. Bei der Glühwürmchengattung Lampyris z. B. haben die Weibchen pro Seite nur 300 Ommatidien, die Männchen hingegen 2.500, beim Junikäfer haben Männchen 3.700 pro Seite, Weibchen 2.700. Dieser Unterschied entstand meistens daraus, dass die Männchen bei der Paarung die Weibchen aufsuchen müssen. Generell haben schneller fliegende Insekten mehr Einzelaugen. Große Libellen haben bis zu 28.000 Ommatidien pro Auge.[1]
Aufbau eines Ommatidiums
Ein Facettenauge hat ein bienenwabenähnliches Muster, und wird daher manchmal auch „Netzauge“ genannt. Es besteht aus einer Anzahl von Einzelaugen, den Ommatidien (Einzahl: Ommatidium).
Der Lichtbrechungsapparat des Einzelauges wird aus einer Chitinlinse und einem darunter liegenden Kristallkegel gebildet. Die Chitinhülle des Ommatidiums bildet die Chitinlinse (Cornea) (A) aus, durch die das Licht ins Auge fällt. Danach fällt es durch den Kristallkegel (B). Bei manchen Arten besteht der Kristallkegel aus Zellen, bei anderen ist es eine durch einige Zellen abgeschiedene extrazelluläre Masse. Dieser Kristallkegel leitet das Licht zu den Sehstäbchen (Rhabdom) (E). Das Rhabdom besteht aus Rhabdomeren, den jeweiligen Mikrovilli-Säumen der acht bis neun Sehzellen. Die Rhabdomere bilden den lichtempflindlichen Teil der Sehzellen und enthalten Sehpigmente (F). Am inneren Ende des Einzelauges beginnen die Nervenfasern (H) der nachgeschalteten Neurone, welche die Wahrnehmungen zum Gehirn weiterleiten. Fällt Licht auf das Rhabdom, depolarisieren die Sehzellen und leiten das Signal so an die für Nervenzellen typischen Axone weiter.[2]
Augentypen: Appositionsaugen und Superpositionsaugen
Es gibt drei unterschiedliche Typen von Facettenaugen, die sich in Aufbau und neuronaler Verschaltung voneinander unterscheiden. Die überwiegende Mehrzahl von Insekten verfügt über Appositionsaugen. Als Appositionsauge wird ein Facettenauge mit durchgängigen Pigmentzellen bezeichnet. Das heißt, die Ommatidien werden durch Pigmentzellen ganz, ohne Zwischenräume, getrennt. Das führt dazu, dass die Insekten ein scharfes Bild sehen können. Aber das Licht erreicht nur diejenige Ommatidie, die zur Linse gehört, durch die es hereingekommen ist. Das restliche Licht wird von den Pigmentzellen verschluckt. Daher ist das Auge nicht für nachtaktive Insekten (wie nachtaktive Lepidoptera) geeignet, sondern für Taginsekten. Nachtaktive Insekten haben als optische Superpositionsaugen bezeichnete Facettenaugen. Schnellfliegende Insekten (z. B. Diptera) verfügen über neuronale Superpositionsaugen.
Bei Appositionsaugen und optischen Superpositionsaugen sind die acht Rhabdomere eines Ommatidiums zu einem einzigen Rhabdom im Zentrum verschmolzen. Bei optischen Superpositionsaugen, die vor allem bei dämmerungs- und nachtaktiven Insekten (z. B. nachtaktive Lepidoptera) vorkommen, sind die Pigmentzellen um die einzelnen Ommatidien abhängig vom Lichteinfall verkürzbar. Ist es dunkel, ziehen sich die Pigmentzellen Richtung Linse zurück und das einfallende Licht kann auch auf das Rhabdom eines benachbarten Ommatidiums fallen. Die so erhöhte Lichtstärke verringert jedoch die Schärfentiefe sehr stark.
Bei neuronalen Superpositionsaugen (z. B. bei schnellfliegenden Insekten wie Diptera) sind, wie beim Appositionsauge, die Pigmentzellen durchgängig. Im Gegensatz zum Appositionsauge und zum optischen Superpositionsauge gibt es in jedem Ommatidium kein fusioniertes Rhabdom, sondern es sind nur zwei Rhabdomere der acht Pigmentzellen (F) miteinander fusioniert. Die sieben einzelnen Rhabdomere sind so angeordnet, dass es ein zentrales Rhabdomer und darum jeweils sechs periphere Rhabdomere gibt. Jedes periphere Rhabdomer ist neuronal mit dem zentralen Rhabdomer des gegenüberliegenden Ommatidiums verschaltet, weshalb dieser Augentyp als neuronales Superpositionsauge bezeichnet wird. [3]
Leistungen
Die räumliche Auflösung des Facettenauges ist durch die Anzahl der Bildpunkte begrenzt und ist somit weit geringer als etwa die Auflösung des menschlichen Linsenauges. Zudem ist die Empfindlichkeit des Facettenauges gering und nimmt mit steigender Auflösung, also mehr und damit kleineren Facetten, stark ab. Dies ist prinzipbedingt, da das von einem Objektpunkt ausgehende Licht im Idealfall nur von einer einzigen der winzigen Linsenflächen (z. B. 0,001 mm²) auf eine Sinneszelle geführt wird, während im Linsenauge die von der gesamten Linsenöffnung (bis zu 40 mm² beim Menschen) empfangenen Strahlen eines Objektpunktes effektiv auf wenige Sinneszellen konzentriert werden.
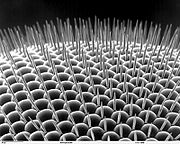 Rasterelektronenmikroskopische Aufnahme des Facettenauges einer Taufliege
Rasterelektronenmikroskopische Aufnahme des Facettenauges einer TaufliegeIm Gegensatz dazu kann die zeitliche Auflösung bei Facettenaugen weit höher sein. Sie liegt etwa bei schnell fliegenden Insekten mit neuronalem Superpositionsauge bei bis über 300 Bildern pro Sekunde, was etwa dem achtfachen des menschlichen Auges von 40 Bildern pro Sekunde entspricht. Bei einem Appositionsauge liegt die zeitliche Auflösung bei etwa 80 Bildern pro Sekunde. Im Gegensatz zum Appositionsauge wird im neuronalen Superpositionsauge das Signal der Sehzellen von nachgeschalteten Neuronen nicht nur weitergeleitet, sondern direkt ausgewertet und nur die Veränderung des Signals weitergemeldet, was die hohen Flimmerfusionsfrequenzen ermöglicht. (Es wird mathematisch die Ableitung der Depolarisation weitergeleitet, bei weniger Licht hyperpolarisiert das nachgeschaltete Neuron, bei mehr Licht depolarisiert es. Die Spannung ist dabei dem Potential der Sinneszelle entgegengesetzt.) Durch diese Datenreduktion kann das Signal schneller und effektiver bearbeitet werden, als beim Appositionsauge, bei dem nachgeschaltete Neurone das Signal der Rezeptorzelle direkt nachahmen.
 Facettenauge eines Krebses (Antarktischer Krill (Euphausia superba))
Facettenauge eines Krebses (Antarktischer Krill (Euphausia superba))Die hohe zeitliche Auflösung kann für bewegte Objekte im nachgeschalteten Gehirn zu einer weit höheren räumlichen Bildauflösung ausgewertet werden, als es nach der durch die Facettenanzahl begrenzten optischen Auflösung entspräche. (Diese Möglichkeit lässt auch dem Menschen bewegte Fernsehbilder weit schärfer erscheinen, als sie im Standbild tatsächlich sind.)
Ferner verfügen Tiere mit Facettenaugen über ein ungleich größeres Blickfeld als mit Linsenaugen. Dies rührt daher, dass prinzipiell jede einzelne der über einen weiten Blickwinkel angeordneten Facetten prinzipiell die gleiche Auflösung erreicht. Bei einem Linsenauge ist eine scharfe Abbildung auf die Bildmitte beschränkt und wird zum Rand des Blickfeldes immer unschärfer.
Bei vielen Insekten werden die Facettenaugen in ihrer Funktion noch unterstützt durch drei punktförmige Lichtsinnesorgane, sogenannte Ocellen - diese dienen jedoch vor allem dem Nachstellen der inneren Uhr.
Literatur
- Kristallmodell lässt Facettenaugen entstehen in: Naturwissenschaftliche Rundschau 10/2005, S. 546 / Science 308, 191 (2005) (englisch)
Weblinks
- Das Facettenauge im Rasterelektronenmikroskop (Uni Ulm)
- Auge der Hornisse
- Function of Compound Eye auf Scholarpedia
Quellen
- ↑ Hans Ekkehard Gruner (Hrsg.), M. Moritz, W. Dunger; 1993; Lehrbuch der speziellen Zoologie, Band I: Wirbellose Tiere, 4. Teil: Arthropoda (Ohne Insekta)
- ↑ Hans Ekkehard Gruner (Hrsg.), M. Moritz, W. Dunger; 1993; Lehrbuch der speziellen Zoologie, Band I: Wirbellose Tiere, 4. Teil: Arthropoda (Ohne Insekta)
- ↑ Rüdiger Wehner, Walter Gehring; 1990; Zoologie, 22. Auflage, S. 407ff




Wikimedia Foundation.