- Frühmensch
-
Als Hominisation wird der stammesgeschichtliche Prozess der Menschwerdung bezeichnet, in dessen Verlauf sich die für die Gattung Homo charakteristischen körperlichen, kognitiven und kulturellen Eigenschaften herausgebildet haben. Hierzu gehören insbesondere die Entwicklung des aufrechten Ganges, des typischen menschlichen Gebisses mit verkürztem, parabolischem Zahnbogen und kleinen Eckzähnen, der späte Eintritt der Geschlechtsreife sowie die Vergrößerung des Gehirns und die hiermit verbundenen geistigen und sozialen Fähigkeiten, die die Vertreter der Gattung Homo von den anderen Menschenaffen (Hominidae) unterscheiden.
Inhaltsverzeichnis
Anatomische Besonderheiten und Evolution
Fünf Merkmale unterscheiden dem US-amerikanischen Anatomen C. Owen Lovejoy zufolge den Menschen von den anderen Arten der Menschenaffen: ein großer Neocortex, der aufrechte Gang, verkleinerte Schneide- und Eckzähne, materielle Kultur sowie ein einzigartiges sexuelles und reproduktives Verhalten.[1]
Aufrechter Gang
 Ikonografie des Fortschritts. Diese populäre Darstellung der Evolution des aufrechten Gangs kann den falschen Eindruck vermitteln, Evolution sei ein gerichteter Verbesserungsprozess.
Ikonografie des Fortschritts. Diese populäre Darstellung der Evolution des aufrechten Gangs kann den falschen Eindruck vermitteln, Evolution sei ein gerichteter Verbesserungsprozess.Anhand der Skelettmerkmale zahlreicher Fossilfunde konnte belegt werden, dass sich der aufrechte, zweibeinige Gang in der Familie der Menschenaffen deutlich früher entwickelte als die starke Vergrößerung des Gehirns. C. Owen Lovejoy, ein Anatom an der Kent State University, bezeichnete 1988 den Übergang zum aufrechten Gang als die augenfälligste Veränderung der Anatomie, die man in der gesamten Evolutionsbiologie bisher nachgewiesen habe.[2] Die Fähigkeit zum dauerhaft aufrechten Gehen setzt eine Vielzahl von Anpassungen des Skletts voraus, insbesondere im Bereich des Beckengürtels und der Hüftgelenke, der Kniegelenke, der Füße sowie des Foramen magnum.[3]
Richard Leakey zufolge ist diese Veränderung derart einzigartig, „dass wir berechtigt sind, alle Arten von zweibeinigen Menschenaffen als menschlich (human) zu bezeichnen.“[4] Eine so weitgehende Festlegung ist allerdings in Fachkreisen umstritten, da nicht alle Fossilfunde von zumindest zeitweise aufrecht gehenden, menschenaffen-ähnlichen Individuen der unmittelbaren Vorfahrenreihe des Menschen zuzuordnen sind.[5]
Aufgrund von Fossilfunden wie „Little Foot“ und „Lucy“ gilt als gesichert, dass schon die Vertreter der Gattung Australopithecus über größere Strecken hinweg aufrecht gehen konnten. Bereits Homo erectus verfügte, wie bei Laetoli freigelegte, 1,51 bis 1,53 Millionen Jahre alte fossile Fußspuren belegen, sowohl über einen im Wesentlichen dem modernen Menschen entsprechenden Bau der Füße als auch über eine vergleichbare Form der aufrechten, zweibeinigen Fortbewegungsweise.[6] Den Analysen zufolge waren die Zehen relativ kurz, der große Zeh war – anders als bei Affen – parallel zu den anderen Zehen ausgerichtet; die Füße waren wie beim modernen Menschen ein wenig nach oben gewölbt. Beim Laufen verlagerte sich das Gewicht von der Ferse zum Ballen. Aus den Abständen der Fußabdrücke sowie aus ihrer Größe und Tiefe schlossen die Forscher, dass die Erzeuger der Spuren eine ähnliche Körpergröße und ein ähnliches Gewicht wie die modernen Menschen besaßen.
In der gleichen Region waren zuvor schon 3,6 Millionen Jahre alte Fußspuren von Australopithecus afarensis entdeckt worden,[7] die dessen bipede Fortbewegungsweise konserviert hatten.
Die Hypothese von der Entwicklung des aufrechten Gangs auf Bäumen

Orang Utan, aufrecht im Baum stehend; Zeichnung nach einem Titelbild von Science [8] Nach Auffassung eines Forscherteams um Susannah Thorpe von der Universität Birmingham und Robin Crompton von der Universität Liverpool könnte sich der aufrechte Gang bereits bei den auf Bäumen lebenden Vorfahren des Menschen entwickelt haben, um auf diese Weise die Früchte am Ende dünner Zweige besser erreichen zu können. Nach einem Bericht der Zeitschrift Science aus dem Jahre 2007 haben die Forscher ein Jahr lang Orang Utans auf der Insel Sumatra beobachtet.[9] Diese Menschenaffen verbringen ihr ganzes Leben auf Bäumen und seien daher ein Modell dafür, wie unsere Vorfahren vor mehreren Millionen Jahre gelebt haben könnten. Die Analyse von rund 3000 Bewegungen ergab, dass die Orang-Utans sich auf sehr dünnen Zweigen auf zwei Beinen fortbewegen, sich dabei mit den Händen an darüber hängenden Zweigen festhalten und mit den Armen ihr Gewicht ausbalancieren. An mitteldicken Zweigen ließen sie sich dagegen eher hängen, sehr dicke Äste würden im Vierfüßler-Gang gemeistert. Demzufolge wären unsere Vorfahren weitaus früher, als zuvor angenommen, auf zwei Beinen unterwegs gewesen. Erst später, als die afrikanischen Wälder während einer Trockenperiode nach und nach verschwanden, hätten sie mit dem „Umzug“ auf den Boden reagiert, wo sie den aufrechten Gang weiter entwickelten und perfektionierten. Die andere Linie, die zu den heutigen Schimpansen und Gorillas führte, hätte dagegen den Vierfüßler-Gang auf den Handknöcheln („Knöchelgang“) weiterentwickelt, um in den ausgedünnten Wäldern rasch von einem Baum zum nächsten gelangen zu können. Unterstützt wird diese Hypothese durch die Tatsache, dass einige Homininifunde aus Gegenden stammen, die zu ihren Lebzeiten eindeutig bewaldet waren. Dies gelte zum Beispiel für den im Jahr 2000 entdeckten Orrorin tugenensis und für Australopithecus-Funde wie „Lucy“.
Aufrechter Gang und Nahrungsaufnahme
Eine Hypothese zum Entstehen des aufrechten Ganges infolge der Nahrungsaufnahme in einer bestimmten Haltung („postural feeding hypothesis“) wurde vom Paläoanthropologen Kevin D. Hunt von der Indiana University ins Gespräch gebracht.[10] Diese Theorie macht geltend, dass Schimpansen bei der Nahrungsaufnahme regelmäßig zweibeinig seien. Auf dem Boden würden sie nach oben greifen, um an Früchte zu gelangen, die von kleineren Bäumen hingen, und auf den Bäumen würde die Fähigkeit zur zeitweisen Bipedie beim Greifen nach einem über ihnen befindlichen Ast genutzt. Diese zweibeinigen Bewegungen entwickelten sich der Theorie zufolge zu häufigeren Gewohnheiten. Hunts Hypothese kann somit als Vorläufer zur Hypothese von der Entwicklung des aufrechten Gangs auf Bäumen beschrieben werden.
Thermoregulatorisches Modell
Das thermoregulatorische Modell geht auf den Zoologen und Evolutionsbiologen Peter Wheeler von der Liverpool John Moores University zurück. Es beruht darauf, dass Bipedie eine erleichterte Abgabe der Körperwärme zur Folge habe: Je höher der Körper eines Menschenaffen sich über den Boden eroben habe, desto eher sei er höheren Windgeschwindigkeiten ausgesetzt gewesen. Während Hitzezeiten bewirke der stärkere Windstrom eine leichtere Abgabe der Körperwärme, was für den Organismus behaglicher sei. Außerdem erklärt Wheeler, dass eine aufrechte Haltung die direkt auf den Körper einwirkende Sonneneinstrahlung im Vergleich zur Vierbeinigkeit reduziere.
Savannen-Hypothese
Als Savannen-Hypothese wurde die Annahme bezeichnet, die Evolution der Bipedie sei bei den Menschenaffen vor rund 7 bis 8 Mio. Jahren dadurch in Gang gekommen, dass die damals noch in Wäldern lebenden Arten ihren Lebensraum in offene, baumlose Savannen verlegt und dort zum aufrechten Gehen gefunden hätten. Diese Hypothese gilt heute aufgrund zahlreicher Fossilfunde als widerlegt, da die frühesten aufrecht gehenden und daher zu den Hominini gestellten Arten wie Sahelanthropus, Orrorin tugenensis und Ardipithecus ramidus keine reinen Savannen-Bewohner waren, sondern in Galeriewäldern lebten.
Verhaltensmodell
Der Anatom C. Owen Lovejoy von der Kent State University leitete 1981 die Bipedie aus dem Sozialverhalten der frühen männlichen Menschenaffen ab.[11] Seine Theorie besagte, dass die Bipedie infolge einer monogame Lebensweise entstanden sei: Die Männchen jener Menschenaffen-Arten, die sich zu den frühen Hominini fortentwickelten, seien monogam geworden und hätten tagsüber ihre Familien alleingelassen, um nach Nahrung zu suchen. Diese Nahrung hätten sie zu ihrer Familie tragen müssen, und die effektivste Fortbewegungsweise sei in dieser Situation das zweibeinige Laufen gewesen. Lovejoys Hypothese, die mangels fossiler Überlieferung nicht unmittelbar durch paläoanthropologische Befunde zu belegen ist, rief umgehend massive Kritik hervor.[12] Insbesondere wurde darauf verwiesen, dass monogame Primaten in der Regel keinen Sexualdimorphismus aufweisen; männliche Exemplare von Australopithecus afarensis besaßen jedoch fast das doppelte Gewicht von Weibchen – dies sei ein Merkmal, das man bei polygamen Arten erwarten würde. Ferner seien monogame Primaten stark territorial, lebten also nicht in größeren sozialen Verbänden; fossile Hinweise zeigten jedoch, dass Australopithecus afarensis in Gruppen lebte.
Wat-Hypothese
Diese Theorie schlägt vor, dass die Menschen die Bipedie als ein Ergebnis des zweibeinigen Watens entwickelten. Zweibeiniges Waten wird bei den teilweise zweibeinig watenden Verwandten des Menschen, den Bonobos, den Flachlandgorillas und den Nasenaffen beobachtet. Zweibeiniges Waten bietet den Vorteil den Kopf zum Atmen über Wasser zu halten. In seinem Buch „Das Geheimnis des aufrechten Ganges“ [13] versucht der Autor, Carsten Niemitz, nachzuweisen, dass für den aufrechten Gang keines der anderes Erklärungsmodell funktionieren könne. Teile dieser Theorie finden sich auch in der so genannten Wasseraffen-Theorie.[14]
Hirnentwicklung
Die Gehirnmasse der fossilen Hominini konnte anhand von Schädelfragmenten zumindest näherungsweise rekonstruiert werden. Die nachfolgende Tabelle gibt hierzu einen Überblick. Da Hirn etwa das spezifische Gewicht von Wasser hat, entspricht das Volumen ungefähr der Masse in Gramm. Hinsichtlich der Intelligenz sind zwischen Mann und Frau keine signifikanten Unterschiede festzustellen (siehe dazu: Gehirne von Männern und Frauen), da das im Mittel geringere Hirngewicht der Frauen eine Folge ihrer im Mittel geringeren Körpergröße ist. Die Angaben zu fossilen Arten sind Schätzungen auf Basis von meist unvollständigen und teils deformierten Funden; unterschiedliche Fachbücher können daher voneinander abweichenden Angaben vorlegen
Typus Volumen in cm3 Durchschnitt Frau Durchschnitt Mann Menschenaffen 400–500[15] Australopithecus 400–550[16] Homo rudolfensis ca. 750[17] Homo habilis 600–800[18] Homo erectus 850–1100[19] Neandertaler ca. 1450
(1300–1750)
[20]Homo floresiensis 380[21] Homo sapiens 1345
(900–1880)[22]1245 1375 Homo sapiens besitzt von allen Primaten den kleinsten Kauapparat und das größte Gehirnvolumen. Nach Erkenntnissen von US-Forschern um Hansell Stedman an der University of Pennsylvania in Philadelphia, die 2004 in der Zeitschrift Nature veröffentlicht wurden, erfolgte vor etwa 2,4 Millionen Jahren eine entscheidende Mutation, die dafür sorgte, dass sich die Muskulatur des Kauapparates zurückbildete, wodurch das Gehirn später „mehr Platz“ für eine Vergrößerung hatte. Die Mutation erfolgte nach Ansicht der Forscher im Gen MYH16, das für die Ausbildung von schweren Myosin-Ketten zuständig ist. Gestützt wird ihre These durch die Tatsache, dass sich erst seit jener Zeit Funde menschlicher Fossilien mit runderen Schädeln, flacheren Gesichtern, kleineren Zähnen und einem schwächeren Gebiss nachweisen lassen. Schimpansen und andere Primaten verfügen nach wie vor über das intakte, nicht mutierte Gen, was bei ihnen für die Bildung einer kräftigeren Kaumuskulatur und eines größeren Kiefers sorgt, jedoch die Vergrößerung des Gehirnschädels verhindert hatte.
Nach Ansicht der US-Anthropologin Andrea Taylor, die die Gebiss-Anatomie sowie die Nahrungsgewohnheiten der Menschenaffen näher studierte, ist jedoch die Annahme, dass sich das menschliche Gehirn allein wegen der Rückbildung der Kiefer- und Kaumuskulatur entfalten konnte, nicht schlüssig. Sie untersuchte das Verhältnis des Kauapparates zum Rest des Schädels bei Berg- und Flachland-Gorillas, Schimpansen und Bonobos und fand keine signifikanten Unterschiede im Gebiss von Blatt- und Obstfressern. „Unsere Ergebnisse zeigen zwar, dass einige Körpermerkmale vielleicht mit den Nahrungsunterschieden der afrikanischen Menschenaffen zusammenhängen. Als einziger Grund für diese Abweichungen in der Körpergestalt scheidet die Ernährung jedoch aus.“ Für die Vergrößerung des menschlichen Gehirnschädels müssten laut Taylor andere Einflüsse wie eine veränderte Umwelt in Betracht gezogen werden.
Während seit den 80er Jahren des 20. Jahrhunderts die Entwicklung des aufrechten Ganges vor allem als Reaktion auf veränderte Umweltbedingungen zurück geführt wurde, kommen französische Forscher wie die Paläontologin Anne Dambricourt-Malassé und die Kieferorthopädin Marie Josèphe Deshayes durch langjährige vergleichende anatomische Untersuchungen zu einem ganz anderen Schluss: Für sie spielt die Evolution des Keilbeins, eines geflügelten Knochens, den der menschliche Embryo als ersten Knochen im mittleren Schädelbereich ausbildet und welcher einen Teil der Schädelbasis sowie den hinteren Bereich der Augenhöhle formt, die entscheidende Rolle bei der Entwicklung des aufrechten Ganges und des Schädels im Laufe der Hominisation. Ihrer Ansicht nach hat die Lage und die Form des Keilbeins, welches sich durch ständige Mutationen immer wieder verändert, einen entscheidenden Einfluss auf die Ausbildung des Gebisses, die Verflachung des Gesichtsschädels und auf die Vergrößerung des Hirnschädels gehabt.
Behaarung
Paläoanthropologisch relevante Haarfunde sind bisher nicht entdeckt worden, daher sind Aussagen zur Behaarung ausgestorbenen Spezies nicht möglich. Auffallend beim heute lebenden Menschen ist gleichwohl die geringe Körperbehaarung. Die relative Nacktheit des Menschen wurde in Bezug gebracht zu seiner Fähigkeit, sich am Feuer zu wärmen sowie sich zu kleiden (siehe auch Artikel Nacktheit), jedoch ist dies reine Spekulation. Eine weitere Hypothese ist, dass der Verlust des Haarkleids eine Anpassung an die verringerte Sonneneinstrahlung in nördlicheren Regionen sei, wodurch die Produktion von Vitamin D in der unbehaarten Haut erleichtert wird.
Das über einige Jahre ungebremste Wachstum des Haupthaares des Menschen findet keine Parallele im Tierreich. Eine klare Funktion ist nicht ersichtlich.
Sexualverhalten
Genetisch determinierte Besonderheiten
- Versteckter Eisprung: Die Fruchtbarkeit von Tierweibchen wird in der Regel durch körperliche oder Verhaltens-Signale mitgeteilt, damit in dieser Phase eine Befruchtung stattfinden kann; bei Menschen ist dies nicht der Fall, die fruchtbare Phase ist für beide Geschlechter kaum ohne hochentwickelte Diagnostik erkennbar.
- Anders als bei fast allen Tieren wird Sexualität bei Menschen nicht nur zu bestimmten, genetisch fixierten Zeiten (Brunftzeiten) praktiziert. Menschen können grundsätzlich ganzjährig miteinander sexuell verkehren, und der Geschlechtsakt ist weniger stark als bei den Tieren mit der Fortpflanzung verbunden. Die jederzeit praktizierte Sexualität ist aber auch bei den nächsten Verwandten des Menschen angelegt: Sowohl Bonobos als auch Schimpansen sind für ihr ausgeprägtes Sexualverhalten bekannt.
- Ausbildung von Brüsten bei erwachsenen Frauen unabhängig von einer Laktation, deren Größe vom Fettgewebe bestimmt wird, nicht vom Drüsengewebe. Solche Brüste existieren nur beim Menschen. Auch die auffällige Färbung der Brustwarzenregion und die Größe der Brustwarzenhöfe ist einzigartig unter den Säugetieren. Diese Besonderheiten werden mit einer sexuellen Signalfunktion erklärt, deren genaue Bedeutung umstritten ist.
- Ausbildung eines für Primaten im erigierten Zustand außerordentlich großen Penis, der nicht (ohne Hilfsmittel) verborgen werden kann (beim Gorilla demgegenüber unscheinbar).
- Koitus ist in vielfältigen Positionen möglich, die eher häufig praktizierte zugewandte Lage kommt bei anderen Primaten - außer beim Orang Utan - nicht vor und ist auch sonst selten im Tierreich. Der Nutzen des vis-a-vis könnte in der Kommunikation liegen.
- Der direkten Ansicht verborgene primäre Geschlechtsorgane bei der erwachsenen Frau.
- Eine Hervorhebung der adulten primären Geschlechtsorgane durch deutlich abgesetzte Schambehaarung bei sonst geringer Behaarung ist im Tierreich einzigartig. Die Funktion könnte darin liegen, Menschen im unbekleideten Zustand auf größere Entfernung hinweg als adult zu erkennen.
- Beginn der Fruchtbarkeit im Vergleich zu anderen (auch langlebigen) Primaten erheblich verzögert.
- Beendigung der Fruchtbarkeit von Frauen: Ab einem Alter von ca. 50 Jahren erleben Frauen durch die Wechseljahre ein Ende ihrer Fruchtbarkeit.
Kulturell etablierte Besonderheiten
- Hauptartikel: Sexualität des Menschen
- Kopplung von Scham und Sexualität: Menschen sind die einzige Spezies, die kulturell Scham für Sexualität entwickeln kann; Geschlechtsverkehr findet üblicherweise unter Ausschluss der Öffentlichkeit statt, während Tiere in der Regel vor den Augen der Artgenossen kopulieren.
- „Offizielle“ Monogamie: Dieses umstrittene Merkmal bezieht sich darauf, dass viele (nach einer anderen Position: die meisten) menschlichen Kulturen mehr oder weniger langfristige Paarbeziehungen zwischen einer Frau und einem Mann zum Zweck der Kinderaufzucht kennen. Relativ selten kommen offizielle Polygynie oder Polyandrie vor. Die offizielle Monogamie ist allerdings nachweisbar gekoppelt mit einer Neigung beider Geschlechter zu „Seitensprüngen“.
- Vorschriften und Verbote von Sexualpraktiken in vielen (oder den meisten) Kultur- und Religionsvorschriften.
Eine Reihe von Autoren vertritt die Position, dass der versteckte Eisprung, die Sexualität zum Vergnügen und die Privatheit des Sexualaktes Merkmale sind, die die Bindung des Mannes an eine Frau, d.h. die zur Kinderaufzucht notwendige langfristige wirtschaftliche Kooperation von Paaren begünstigt haben. Die Beendigung der Fruchtbarkeit der Frau ab einem Alter, in dem die Lebensgefährdung durch eine Geburt eine bestimmte Schwelle überschreitet, komme ebenfalls der Kinderaufzucht zugute und habe sich deshalb in der Evolution bewährt und durchgesetzt.
Weitere anatomische und kulturelle Besonderheiten
Zu den anatomischen Besonderheiten zählen vor allem die leistungsfähige Greifhand, das größere Gehirn, der aufrechte Gang und sehr modulationsfähige Stimmbänder als Voraussetzung für die Fähigkeit des Sprechens.
Zu den kulturellen Merkmalen zählen die extrem gesteigerte Fähigkeit zu kollektivem Lernen durch Sprache (d.h. symbolische Kommunikation), Kooperation und Technikeinsatz (Feuer, Werkzeuge, Landwirtschaft etc.). Mit zunehmender Leistungsfähigkeit der menschlichen Technik wächst die Abhängigkeit von ihr und gleichzeitig der Aufwand, der bei der Kinderaufzucht zur Weitergabe des technischen Wissens betrieben werden muss, bis die Nachkommen sich relativ selbständig ernähren und wiederum fortpflanzen können. Dieser wachsende Aufzuchtaufwand wirkte über hunderte von Generationen als Selektionsdruck auf Anatomie und Sexualphysiologie zurück.
Der „Stammbaum“ des Menschen
Die Frühgeschichte der Menschenaffen kann etwa 32 Mio. Jahre zurückverfolgt werden, beginnend mit den Funden aus der Provinz Madinat al-Fayyum in Ägypten. Dort konnten die Überreste von sechs verschiedenen Primatengattungen gefunden werden. Man nimmt heute an, dass die Menschwerdung vor etwa 8 bis 5 Millionen Jahren in Afrika begann. Stammesgeschichtlich haben sich die Vorfahren der Schimpansen zu dieser Zeit von der zum Menschen führenden Entwicklungslinie abgetrennt.
Eine im Sommer 2006 in der Zeitschrift Nature veröffentlichte Genanalyse[23] des Broad Institute, eines gemeinsamen Instituts des Massachusetts Institute of Technology (MIT) und der Harvard University, wurde dahingehend interpretiert, dass eine erste Abspaltung noch wesentlich früher erfolgt sein könnte und der Entwicklungsgang der frühesten Hominini und der Vorfahren der anderen Menschenaffen ungewöhnlicher verlief, als zuvor angenommen wurde. In dieser Studie waren 20 Millionen Basenpaare von menschlicher DNA sowie von Schimpansen- und Gorilla-DNA miteiannder verglichen worden. Die Untersuchungen bezogen sich primär auf die sogenannte molekulare Uhr: Dabei wird der Verlauf der Evolution mit Hilfe von Genmutationen berechnet, die sich in besonderen „Schlüsselsequenzen“ des Erbguts abgespielt haben. Dies ermöglichte den Autoren zufolge eine weit präzisere Datierung und Bestimmung der Auseinanderentwicklung der Hominini und der anderen Menschenaffen.
Laut Studie separierte sich eine frühe Affenart bereits vor ca. 10 Millionen Jahren vom gemeinsamen Vorfahren. Die unterschiedlichen Populationen vereinigten sich jedoch einige Jahrtausende später wieder und bildeten eine Mischpopulation, die zu Kreuzungen mit den Vorfahren tendierte. Den Interpretationen der Forscher zufolge gab es über vier Millionen Jahre hinweg einen Abfolge von Kreuzungen und sich auseinander entwickelnden Gruppen, bis eine letztmalige, dauerhafte Trennung der Schimpansenvorfahren und der Hominini vor ca. 6,3 bis 5,4 Millionen Jahren erfolgte. Dieser letzte Genaustausch sei durch das durchgängig sehr junge Alter der X-Chromosomen belegt, die sich erst zu diesem späten Zeitpunkt in der für Menschen charakteristischen Form herausbildeten und den X-Chromosomen des Schimpansen sehr ähnelten. Dieses Szenario blieb allerdings nicht unwidersprochen.
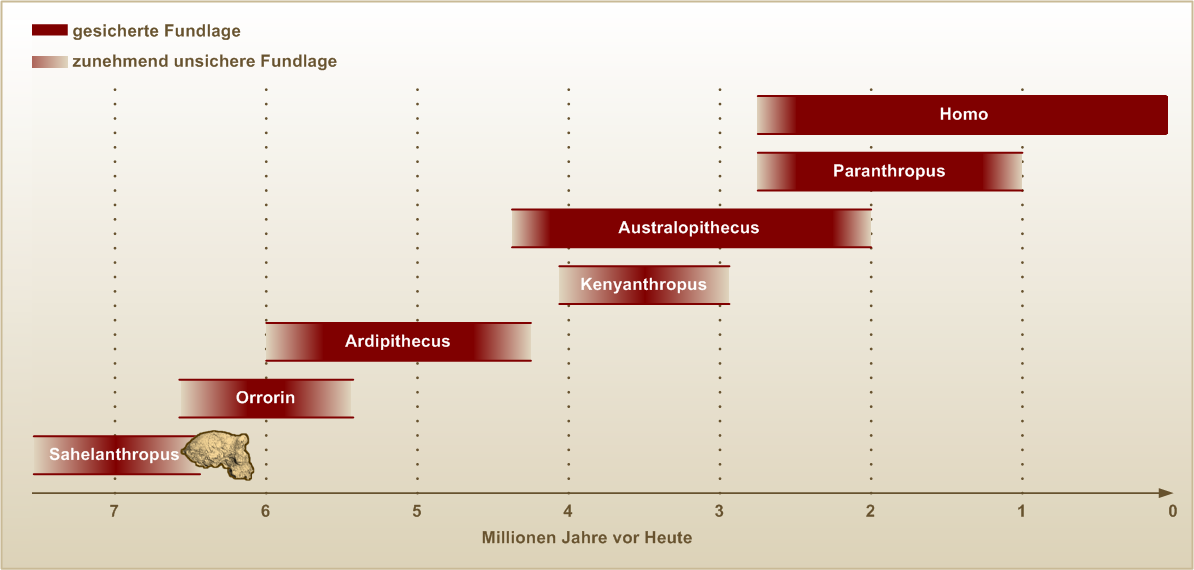
Zeittafel der Hominini-Gattungen
Die frühesten aufrecht gehenden Hominini
Nach dem Fund des Orrorin tugenensis („Millennium-Mann“) im Jahr 2000 und des Sahelanthropus tchadensis („Toumaï“) im Jahr 2001 werden diese beiden Arten als „älteste Vorläufer des Menschen“ erwogen. Sollten sich die Annahmen bestätigen, gab es schon vor 6 bis 7 Millionen Jahren aufrecht gehende Hominini. Bereits 1994 waren in Äthiopien Fossilien entdeckt worden, die etwa 4,4 Millionen Jahre alt sind und als Ardipithecus ramidus (von ramid = die Wurzel) bezeichnet werden. Auch die Vertreter dieser Art konnten vermutlich aufrecht gehen. Die verwandtschaftlichen Beziehungen zwischen diesen drei Arten sowie von Ardipithecus, Orrorin und Sahelanthropus zu den späteren Hominini sind ungeklärt.
Australopithecinen
- Hauptartikel: Australopithecinen
Zu den relativ gut bekannten, bereits aufrecht gehenden Vorläufern des Menschen gehören die Australopithecinen, deren Funde bisher auf Ost-, West- (Tschad) und Südafrika beschränkt sind. Nach den bisher vorliegenden Funden unterscheidet man verschiedene Arten, darunter Australopithecus anamensis, Australopithecus africanus und Australopithecus afarensis. Besonders bekannte Einzelfunde sind das „Kind von Taung“ (ein Australopithecus africanus), „Lucy“ (ein – vermutlich männlicher – Australopithecus afarensis) sowie das jugendliche, weibliche Skelett DIK 1-1 („Selam“).
Ferner wird eine Sonderlinie von Australopithecinen mit einem robusteren Gebiss neuerdings zur Gattung Paranthropus gezählt. Umstritten ist ferner, ob der 1999 von Meave Leakey in Kenia entdeckte Kenyanthropus platyops („Flat Faced Man“) als eigenständigen Gattung anzusehen ist oder zur Gattung Australopithecus zu stellen ist.
Bei Laetoli in der Olduvai-Schlucht in Tansania wurden fossile Fußspuren von mehreren Australopithecus-afarensis-Individuen gefunden, die ebenfalls eindeutig belegen, dass Vertreter dieser Art aufrecht gingen.
Anpassungen bei Australopithecinen
Eine Klimaveränderung, ausgelöst durch die plattentektonische Hebung Ostafrikas, bewirkte eine weitgehende Versteppung des angestammten Lebensraums. Diese Grassteppe bot in erster Linie Nahrung für Grasfresser (Paarhufer, Wiederkäuer), die es vorher schon, meist in kleineren Formen, als Waldbewohner gab. Diese traten nun bald in großen Herden auf, und weil sie zahlreicher wurden, konnten sich auch Raubtiere und Aasfresser vermehren.
So differenzierten sich zwei Typen von Hominini. Die eine Strategie war eine biologische Anpassung an das neue zellulosereiche Nahrungsangebot. Australopithecus robustus und andere Arten entwickelten als Anpassung eine gewaltige Kaumuskulatur und entsprechend mächtige Molaren. Die Muskulatur setzte dabei an einem deutlich sichtbaren Knochenkamm auf dem Scheitel des Schädels an. Es gab verschiedene, meist mächtige und große Primaten, die diese ökologische Nische zu nutzen versuchten, die allerdings allesamt wieder ausstarben.
Eine andere Strategie war die des Fleischfressers. In den Anfängen dürfte sich dies aber auf Aas und Beuteraub beschränkt haben, da diese Hominini weder die Fähigkeit besaßen, als Raubtier größere Beutetiere zu stellen, noch über Klauen oder Zähne verfügte, die geeignet gewesen wären, ein Beutetier zu töten oder aufzubrechen. Sehr wahrscheinlich kam es hier zum ersten Werkzeuggebrauch, indem zufällig gefundene scharfkantige Steine dazu benutzt wurden, Beutetiere aufzubrechen.
Aber damit war die biologische Evolution nicht abgeschlossen. Unter dem Selektionsdruck, Werkzeuge und Kommunikation zu verfeinern, und dem Angebot von reichlich hochwertigem Eiweiß waren höhere intellektuelle Fähigkeiten von Vorteil. Nachdem einige Vertreter er Hominini schon Jahrmillionen zuvor den aufrechten Gang erworben hatte, entwickelte er nun auch ein größeres Gehirn. Da sich das weibliche Becken unter Einfluss des aufrechten Ganges aber nicht an den wachsenden Kopfumfang des Neugeborenen anpassen konnte, kamen diese zu einem biologisch immer weiter vorverlegten Termin, also immer weniger weit entwickelt und immer mehr und länger auf Brutpflege angewiesen, zur Welt. Zudem war mehr Zeit nötig, um die Fähigkeiten von den Erwachsenen zu erlernen (vertikale Proliferation im Gegensatz zur horizontalen Proliferation durch Vererbung).
Die Gattung Homo
Aus den Australopithecinen, womöglich aber auch aus dem „Flat Faced Man“, entwickelten sich vor zwei bis drei Millionen Jahren die ersten Vertreter der Gattung Homo, deren Fossilien insbesondere aufgrund von Werkzeugfunden zu Homo gestellt wurden.
Hierbei handelt es sich um Homo rudolfensis (benannt nach dem Rudolf-See, heute Turkana-See, in Kenia) und Homo habilis (der „geschickte“ Mensch). Als etwas jüngere Art gilt Homo ergaster. Die Verwandtschaftsbeziehungen dieser Arten sind bislang umstritten.
Vor etwa eineinhalb bis zwei Millionen Jahren entwickelte sich Homo erectus. Diese Menschenform gilt bislang als die erste, die Afrika verließ und sich über den vorderen Orient nach Europa und Asien auszubreiten begann; der erste Nachweis der Gattung Homo außerhalb Afrikas stammt allerdings von den homininen Fossilien von Dmanisi, die auf 1,8 Millionen Jahre datiert wurden und deren Anbindung an den Stammbaum der Gattung Homo ungeklärt ist. Als gesichert gilt, dass Homo erectus als erster das Feuer zu beherrschen lernte. Umstritten ist jedoch, ob die in Kenia gefundenen 1,4 Millionen Jahre alten Spuren verbrannten Lehms bereits als Feuerstellen des Homo erectus gedeutet werden können. Gesichert ist der Feuergebrauch vor 500.000 Jahren in China und vor 400.000 Jahren in der Bretagne.
Homo floresiensis, scherzhaft auch „Hobbit“ genannt, dessen Überreste 2003 auf der indonesischen Insel Flores entdeckt wurden, wird von einigen Forschern für eine späte „Zwergform“ des Homo erectus gehalten. Diese Ansicht wird durch die Analyse der Schädelform gestützt. Neuere Untersuchungen der Handgelenke deuten aber darauf hin, dass Homo floresiensis höchstens auf der Stufe eines Australopithecinen stand. Andere anatomische Merkmale wurden als krankhafte Abweichung von Homo sapiens gedeutet. Eine Klärung der Einordnung dieser Funde in den Stammbaum der Gattung Homo steht wegen der zurzeit noch widersprüchlichen Deutungen aus.
Vor ca. 800.000 Jahren entwickelte sich aus Homo erectus eine Form mit größerem Gehirn, die als Homo heidelbergensis bezeichnet wird. Es ist umstritten, ob sie als eine Unterart des Homo erectus (europäische Paläoanthropologie) oder als eigene Art (angloamerikanische Paläoanthropologie) zu klassifizieren ist. Aus Homo heidelbergensis bzw. Homo erectus heidelbergensis entwickelten sich in Europa die Neandertaler (Homo neanderthalensis), während zur selben Zeit aus den in Afrika verbliebenen Populationen des Homo erectus der Homo sapiens hervorging, der heutige Mensch (siehe auch: Archaischer Homo sapiens und Hofmeyr-Schädel).
Die vier genannten Menschenformen könnten sogar teilweise gleichzeitig gelebt haben, denn noch aus der Zeit von vor ca. 50.000 Jahren wurden in abgelegeneren Gebieten, so auf Java, Fossilien des Homo erectus gefunden.
Ursprung des Homo sapiens
- Hauptartikel: Ausbreitung des Menschen
Über die Ursprünge des Homo sapiens gingen die Meinungen lange Zeit auseinander. Von den Paläoanthropologen weitgehend akzeptiert ist heute die auf Fossilfunden basierende Out-of-Africa-Theorie, der zufolge die Ausbreitung des modernen Menschen vor etwa 60.000 bis 70.000 Jahren aus Afrika in alle anderen Regionen der Erde begann. Vertreter der „multiregionalen Hypothese“ vertraten hingegen die Ansicht, dass sich lokale Populationen des Homo erectus, die viel früher aus Afrika auswanderten, mehrfach unabhängig voneinander in verschiedenen Regionen der Welt zum modernen Menschen entwickelten, wobei es in Kontaktzonen zu einer genetischen Vermischung lokaler Gruppen der dortigen frühen Menschen gekommen sei. Genetische Analysen des Y-Chromosoms und der Mitochondrien des Menschen stützen inzwischen die Out-of-Africa-Theorie. Eine Vermischung zwischen Homo sapiens und den späten Vertretern von Homo erectus (hierzu gehört auch der europäische Neandertaler) ist diesen Befunden zufolge äußerst unwahrscheinlich, auch wenn sie nicht mit völliger Sicherheit ausgeschlossen werden kann.
Die ältesten Funde des modernen Menschen in Afrika sind etwa 160.000 Jahre alt. Außerhalb Afrikas reichen Funde des Homo sapiens bis etwa 100.000 Jahre zurück. Es handelt sich bei ihm um die einzige Menschenart, die Amerika (vor etwa 11.500 bis 15.000 Jahren, nach mancher Ansicht wesentlich früher) und Australien (vor etwa 60.000 Jahren) besiedelt hat. Homo sapiens ist zugleich der letzte Überlebende der Gattung Homo innerhalb der Familie der Menschenaffen.
Quellen
- ↑ Owen Lovejoy: The Origin of Man. Science, Band 211, 1981, S. 341–350, doi:10.1126/science.211.4480.341 – „Five characters separate man from other hominoids – a large neocortex, bipedality, reduced anterior dentition with molar dominance, material culture, and unique sexual and reproductive behavior.“
- ↑ C. Owen Lovejoy: Evolution of Human Walking. Scientific American, November 1988, S. 118–125
- ↑ Ronald J. Clarke, Phillip Tobias: Sterkfontein member 2 foot bones of the oldest South African hominid. Science, Band 269, 1995, S. 521; doi:10.1126/science.7624772
- ↑ Richard Leakey: The origin of humankind. Phoenix, a division of Orion Books Ltd., 1995, S. 13
- ↑ www.pbs.org: The Transforming Leap, from Four Legs to Two. Eine Übersicht zum Stand der Forschung
- ↑ „The Ileret prints show that by 1.5 Ma, hominins had evolved an essentially modern human foot function and style of bipedal locomotion.“ Matthew R. Bennett et. al.: Early Hominin Foot Morphology Based on 1.5-Million-Year-Old Footprints from Ileret, Kenya. Science, Band 323, 2009, S. 1197–1201; doi:10.1126/science.1168132 10.1126/science.1168132
- ↑ Mary Leakey et. al.: Pliocene footprints in the Laetoli beds at Laetoli, northern Tanzania. Nature, Band 278, 1979, S. 317–323; doi:10.1038/278317a0
- ↑ sciencemag.org: Cover von Science, 1. Juni 2007
- ↑ S. K. S. Thorpe et. al.: Origin of Human Bipedalism As an Adaptation for Locomotion on Flexible Branches. Science, Band 316, 2007, S. 1328–1331, doi:10.1126/science.1140799; vergl. dazu: www.sueddeutsche.de: „Aufrecht auf dem Ast“
- ↑ Kevin D. Hunt: The evolution of human bipedality: ecology and functional morphology. Journal of Human Evolution, Band 26, 1994, S. 183–202
Kevin D. Hunt: The postural feeding hypothesis: an ecological model for the evolution of bipedalism. South African Journal of Science, Band 92, 1996, S. 77–90. - ↑ Owen Lovejoy: The Origin of Man. Science, Band 211, 1981, S. 341–350
- ↑ nachzulesen in Science, Band 217, S. 295–304; die kritischen Stellungnahmen stammten von Glynn Isaac, Diahan Harley, James W. Wood, Linda D. Wolfe, J. Patrick Gray, John G. Robinson, Leslie S. Lieberman, Elizbeth H. Peters, Rebecca L. Cann und Allan C. Wilson
- ↑ Carsten Niemitz: Das Geheimnis des aufrechten Gangs. Unsere Evolution verlief anders. C.H.Beck, München, 2004, ISBN 3-406-51606-8
- ↑ Kuliukus, A., Wading for Food: The Driving Force of the Evolution of Bipedalism. Nutrition and Health, 16(4), 267–290, (2002). html
- ↑ Ronald M. Nowak: Walker’s mammals of the world. 6. Auflage. Johns Hopkins University Press, Baltimore 1999, S. 613
Wolfgang Maier: Primates, Primaten, Herrentiere. In: Wilfried Westheide, Reinhard Rieger (Hrsg.): Spezielle Zoologie. Teil 2. Wirbel- oder Schädeltiere. Spektrum Akademischer Verlag, Heidelberg/Berlin 2004, S. 573 - ↑ Steve Jones et. al.: The Cambridge Encyclopedia of Human Evolution. Cambridge University Press, 1992, S. 236
- ↑ G. J. Sawyer, Viktor Deak: Der lange Weg zum Menschen. Lebensbilder aus 7 Millionen Jahren Evolution. Spektrum Akademischer Verlag, Heidelberg, 2008, S. 79
- ↑ Ulrich Welsch: Wie aus den ersten Primaten Homo wurde (1): Die Fossilgeschichte des Menschen. Biologie in unserer Zeit, 1/2007, S. 47
- ↑ stanford.edu
- ↑ palomar.edu
- ↑ D. Falk et al.: Brain shape in human microcephalics and Homo floresiensis. In: PNAS 29. Feb. 2007
- ↑ palomar.edu, [1]
- ↑ Nick Patterson, Daniel J. Richter, Sante Gnerre, Eric S. Lander, David Reich (2006): Genetic evidence for complex speciation of humans and chimpanzees. In: Nature, Bd. 441 (29. Juni 2006), S. 1103-1108.
Siehe auch
- Liste homininer Fossilien
- Die Abstammung des Menschen und die geschlechtliche Zuchtwahl
- Wiege der Menschheit
- Toba-Katastrophen-Theorie
- Mitochondriale Eva
- Adam des Y-Chromosoms
- Holistische Sprachgenesetheorie
- Menschheitsgeschichte
- Humanethologie
Literatur
- Bucher, Zeno (1992): Die Abstammung des Menschen als naturphilosophisches Problem. Koenigshausen & Neumann, Würzburg 1992. ISBN 3-88479-721-2
- Streit, Bruno (1995) Hrsg.: Evolution des Menschen. - Verständliche Forschung. Spektrum Akademischer Verlag, Heidelberg. ISBN 3-86025-267-4
- Reichholf, Josef H. (1997): Das Rätsel der Menschwerdung. Die Entstehung des Menschen im Wechselspiel der Natur. dtv, München 1997. ISBN 3-423-33006-6
- Diamond, Jared (1998): Warum macht Sex Spaß? Die Evolution der menschlichen Sexualität. Bertelsmann, München, Goldmann, München 2000. ISBN 3570120082, ISBN 344215085X
- Diamond, Jared (1999): Der dritte Schimpanse. Fischer Taschenbuch Verlag, Frankfurt a.M.. ISBN 3-596-14092-7
- Burenhult, Göran, u. a. (2000): Die ersten Menschen. Die Ursprünge des Menschen bis 10 000 vor Christus. Jahr, Hamburg. ISBN 3-8289-0741-5
- Kuckenburg, Martin (2001): Als der Mensch zum Schöpfer wurde. An den Wurzeln der Kultur. Klett-Cotta, Stuttgart. ISBN 3-608-94034-0
- Schrenk, Friedemann, Timothy G. Bromage, Henrik Kaessmann (2002): Die Frühzeit des Menschen. Zurück zu den Wurzeln. In: Biologie in unserer Zeit. Verlag Chemie, Weinheim Bd 32, H. 6, S. 352-359. ISSN 0045-205X
- Schüring, Joachim (2006): Von der anderen Art. In: Abenteuer Archäologie. Kulturen, Menschen, Monumente. Spektrum der Wissenschaft, Heidelberg 2006, H. 1, S. 32ff. ISSN 1612-9954 (zu Neanderthaler und Homo, Out of Africa und Genanalyse)
- Shreeve, James (2006): Aus Afrika in die Welt. Was das Erbgut über unsere Herkunft und die Besiedlung der Erde erzählt. in: National Geographic Deutschland. Hamburg 2006,3 (März), S. 38-53.
- Shreeve, James (2006) Die Suche nach Adam und Eva. Ein einzigartiges Genprojekt gibt Auskunft über unsere frühe Geschichte. in: National Geographic Deutschland. Hamburg 2006,3 (März), S. 54-63.
- Junker., Thomas (2006): Die Evolution des Menschen. C.H.Beck, München 2006. ISBN 3-406-53609-3
- Henke, Winfried, Tatersall, Ian (2007) Hrsg.: Handbook of Paleoanthropology. Vol. I - III. Springer, Berlin. ISBN 978-3-540-32474-4
Weblinks
- Als sich die Stammbäume von Schimpansen und Menschen teilten (Der letzte gemeinsame Vorfahr von Mensch und Schimpanse lebte vor etwa sechs Millionen Jahren)
- Der Ursprung des Menschen - Dossier in spektrumdirekt
- focus on human origins (englisch)
- Gerald Bühler: Über den aufrechten Gang und seine Folgen. Aufsatz, Erlangen 2005.
- Der Stammbaum des Menschen - Studienarbeit über die Entwicklungsgeschichte des menschlichen Stammbaums
- Asiaten stammen aus Afrika
- Vom Werkzeugmacher zum Aasfresser: Vorstellungen von der Menschwerdung im Spiegel der Wissenschaftsgeschichte Vortrag von Dr. Inge Schröder, Anthropologisches Institut der Christian-Albrechts-Universität, Kiel; Schröder ist seit 2003 Privatdozentin und seit 2006 wissenschaftliche Geschäftsführerin des Wissenschaftszentrums Kiel
- www.zdf.de : Evolution des Menschen ZDF-Film vom 16. Juli 2006: Interaktive Grafikanimation - mit Stammbusch und Steckbrief der Menschenaffen, ihre Ausbreitung und Erklärung der morphologischen Unterschiede


Wikimedia Foundation.