- Atlanthropus
-
Homo erectus .jpg)
Schädeldach „Sangiran II“
(Original, 1,5 mya), Sammlung Koenigswald im Naturmuseum Senckenberg. Man beachte den Überaugenwulst über dem linken Auge.Zeitraum Pleistozän 1,8 bis 0,04 Millionen Jahre Fossilfundorte Systematik Altweltaffen (Catarrhini) Menschenartige (Hominoidea) Menschenaffen (Hominidae) Hominini Homo Wissenschaftlicher Name Homo erectus (Dubois, 1892) Homo erectus ist eine ausgestorbene Hominiden-Art der Gattung Homo. Das Art-Epitheton erectus kommt aus dem lateinischen und bedeutet „aufgerichtet“. Dieser Art werden Fossilien aus dem Pleistozän zugeordnet, deren älteste dem frühesten Altpleistozän entstammen. Einige Forscher stellen noch Fossilien aus dem mittleren Jungpleistozän zu Homo erectus, so dass dieser „Frühmensch“[1] – aus dieser Sicht – fast zwei Millionen Jahre Afrika, Asien und Europa besiedelt hat. Aus Homo erectus entwickelte sich in Europa der Neandertaler und – parallel zu diesem, aber unabhängig davon – in Afrika der moderne Mensch (Homo sapiens). Die Abgrenzung des Homo erectus von anderen Arten der Gattung Homo ist in Fachkreisen – zwischen sogenannten Lumpern und Splittern – umstritten.
Homo erectus war laut Richard Leakey „die erste hominine Art, die das Feuer benutzte; die erste, die das Jagen als ein wesentliches Element zur Sicherung ihrer Nahrungsversorgung einsetzte; die erste, die wie ein moderner Mensch laufen konnte.“ [2]
Inhaltsverzeichnis
Vorkommen
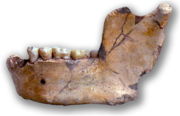
Unterkiefer von Ternifine, zunächst bezeichnet als Atlanthropus mauritanicus Homo erectus war die erste Art der Gattung Homo, die sich weit über Afrika hinaus verbreitete. Heute werden zahlreiche Fossilienfunde dem Homo erectus zugeordnet, die von ihren Entdeckern zunächst eigene Gattungs- und Artnamen erhalten hatten, so zum Beispiel der von Eugène Dubois benannte Anthropithecus („Java-Mensch“), der von Davidson Black benannte Sinanthropus pekinensis („Peking-Mensch“), der von John T. Robinson benannte Telanthropus capensis („Zielmensch“, ein Fund aus Swartkrans, Südafrika) und der von Camille Arambourg benannte Atlanthropus mauritanicus („Atlas-Mensch“, ein Fund aus Ternifine bei Muaskar, Algerien).
Zugleich herrschen unter den Paläoanthropologen unterschiedliche Auffassungen über die Abgrenzung des Homo erectus von anderen Arten der Gattung Homo. Seit etwa 1990 werden die ältesten, rund 1,8 bis 1,5 Millionen Jahre alten afrikanischen Funde von Homo erectus – vor allem von US-amerikanischen Paläoanthropologen – als eigene Chronospezies ausgewiesen und als Homo ergaster bezeichnet. Unterschiedliche Auffassungen herrschen auch in Bezug auf jüngere Funde aus dem Mittelpleistozän, die einige Forscher zu Homo erectus stellen, andere aber als Homo rhodesiensis, Homo antecessor oder Homo heidelbergensis bezeichnen. Möglicherweise gehört auch Homo floresiensis zum Formenkreis des späten Homo erectus.
Seit seiner Entdeckung Anfang der 1960er-Jahre galt Homo habilis als wahrscheinlichster Vorfahre des Homo erectus. 2007 wurde jedoch ein Fund veröffentlicht, der dem Homo habilis zugeordnet wird und auf ein Alter von 1,44 Millionen Jahren datiert wurde. Das würde bedeuten, dass Homo habilis und Homo erectus bis zu einer halben Million Jahre nebeneinander existierten. Die Wissenschaftler vermuten, dass beide Arten unterschiedliche ökologische Nischen besetzten und so keine direkten Konkurrenten waren.[3] Diese Befunde bedeuten zugleich, dass Homo habilis kein Vorfahre des Homo erectus gewesen sein kann; als wahrscheinlichster Vorfahre von Homo ergaster / Homo erectus gilt daher derzeit Homo rudolfensis. Die verwandtschaftlichen Beziehungen von Homo habilis, Homo rudolfensis und Homo ergaster / Homo erectus sind in Fachkreisen allerdings umstritten.
Zu den in jüngster Zeit entdeckten Funden, die möglicherweise Homo erectus zuzurechnen sind, gehören die in Georgien entdeckten Fossilien von Dmanisi.
Erstbeschreibung
- Hauptartikel: Java-Mensch
Im Oktober 1891 entdeckte Eugène Dubois auf der indonesischen Insel Java ein Schädeldach mit einem markanten, durchgehenden Querwulst des Stirnbeins oberhalb der Nasenwurzel („Überaugenwulst“). Dieses Fossil – genannt Trinil II und später Java-Mensch – wurde von Dubois zunächst als nicht-homininer Menschenaffe interpretiert, da er eine Ähnlichkeit mit den rezenten Schimpansen zu erkennen glaubte; daher wurde der Fund der neuen Gattung Anthropithecus [4] zugeordnet. Im August 1892 entdeckte Dubois jedoch einen vollständigen fossilen Oberschenkelknochen (Trinil III), der in Form und Aufbau dem des modernen Menschen glich, was als Hinweis gedeutet werden konnte, dass sein Besitzer sich ähnlich aufrecht wie der moderne Mensch bewegte. Daraufhin erhob Dubois umgehend (1892) beide Fossilien sowie einen bereits im September 1891 entdeckten Backenzahn (Trinil I) zum Typusexemplar einer neuen Gattung und Art Anthropithecus erectus.[5] 1894 änderte er den Gattungsnamen in einem Fachaufsatz erneut in Pithecanthropus erectus [6] und beschrieb ihn als Übergangsform, die zum modernen Menschen geführt habe. Dubois griff dabei einen Namensvorschlag auf, den Ernst Haeckel 1863 eingeführt hatte, als er eine hypothetische Übergangsform zwischen Mensch und Affe beschrieben und diese Pithecanthropus genannt hatte (pithecus = Affe, anthropus = Mensch).[7]
Die von Eugène Dubois entdeckten und beschriebenen Fossilien waren die ersten Funde von Fossilien der Hominini außerhalb Europas und zugleich die ältesten bis dahin aus dieser Klade entdeckten. Aufgrund ihrer Fundsituation am Rande eines ehemaligen Gewässers ist die Datierung der Knochen problematisch; nach heutigem Kenntnisstand wird ihnen ein Alter von rund einer Million Jahre zugeschrieben.[8]
1950 veranlasste ein Vortrag von Ernst Mayr beim Symposium von Cold Spring Harbour die Paläoanthropologen, alle inzwischen entdeckten, mutmaßlichen Vorfahren des Menschen der Gattung Homo zuzuordnen.[9] Dies hatte zur Folge, dass auch die als Pithecanthropus erectus bezeichneten Fossilien erneut umbenannt wurden und seitdem der Holotypus von Homo erectus sind.
Weitere frühe Funde aus Asien
 Sangiran 17 = Pithecanthropus VIII, entdeckt 1969 (Kopie)
Sangiran 17 = Pithecanthropus VIII, entdeckt 1969 (Kopie)Dubois Zuweisung der von ihm in Trinil entdeckten Fossilien zu den Hominini war zunächst äußerst umstritten. Erst nachdem in den 1920er-Jahren die Peking-Menschen und in den 1930er-Jahren von Gustav Heinrich Ralph von Koenigswald auf Java das Fossil Sangiran II sowie weitere, ähnlich alte Funde gemacht worden waren, galt die Zuordnung der Java-Menschen zu den Ahnen des modernen Menschen als gesichert. Allerdings wurden die neuen Funde mit jeweils eigenem Art-Epitheton (meist nach ihrem Fundort) benannt, auch wurden zusätzliche Gattungsnamen vorgeschlagen, so dass die Fossilien aus Java u.a. auch als Homo soloensis, Homo modjokertensis, Pithecanthropus robustus, Pithecanthropus dubius und Meganthropus palaeojavanicus bezeichnet wurden.
Die in Asien entdeckten frühen Vertreter der Hominini führten dazu, dass die Paläoanthropologen zu dem Fehlschluss kamen, der moderne Mensch habe sich in Asien aus affenähnlichen Vorfahren entwickelt, obwohl bereits Charles Darwin 1871 vermutet hatte,[10] der Mensch habe sich in Afrika entwickelt, da seine nächsten Verwandten – Schimpansen und Gorillas – dort beheimatet sind. Diese Fehleinschätzung wiederum verhinderte jahrelang die Anerkennung des ersten afrikanischen Australopithecus africanus-Fundes – des Kindes von Taung – als zugehörig zur Ahnenreihe des modernen Menschen.
Funde aus Afrika
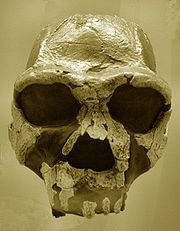
Das Fossil KNM-ER 3733
(Kopie), SenckenbergmuseumDa sich die Suche nach Vorfahren des modernen Menschen zunächst auf Asien konzentrierte und die ersten afrikanischen Funde der Gattung Australopithecus Jahrzehnte lang nicht als den Hominini zugehörig akzeptiert wurden, gab es in Afrika bis in die 1940er-Jahre kaum eine gezielte Suche nach Fossilien. Die ersten, später dem Homo erectus zugeschriebene Fossilien aus Südafrika (Saldanha, 1953) und Algerien (Ternifine, 1954) waren Zufallsfunde, zum Teil schlecht erhalten und erhielten zunächst eigene Gattungsbezeichnungen (Atlanthropus mauritanicus). Erstmals 1961 wurden auch afrikanische Funde, die seit 1949 bekannten und zuvor als Telanthropus capensis bezeichneten Fossilien aus Swartkrans, als Homo erectus interpretiert.[11] Ab den frühen 1960er-Jahren kamen dann zahlreiche weitere Funde, vor allem aus Kenia, Tansania und Äthiopien, aber auch aus Marokko (Steinbruch Thomas bei Casablanca, 1969) hinzu, darunter das 1960 geborgene fossile Schädeldach OH 9 aus Olduvai und das 1975 am Turkana-See (Koobi Fora) entdeckte Schädeldach KNM-ER 3733, die beide den Funden aus Sangiran („Java-Menschen“) und Zhoukoudian („Peking-Menschen“) ähneln.
Merkmale
Der Körperbau von Homo erectus konnte anhand der zahlreichen Funde in Asien genauer rekonstruiert werden als jener der früheren Hominini-Arten. Charakteristisch für Homo erectus ist demnach der im Vergleich zum modernen Menschen „tonnenförmigere“ und volumenreichere Rumpf[12] sowie ihr kräftiges und im Vergleich zu älteren Arten der Hominini größeres Skelett mit besonders dickwandigen Schädelknochen (Schädeldach zwischen 6 und 11 mm) und kräftigen Überaugenwülsten, „über deren Funktion man bis heute rätselt.“[13] Der Schädel ist – von hinten betrachtet – in Relation zu seiner Höhe auffällig breit.
Der Unterkiefer von Homo erectus war im Vergleich zu Homo sapiens breiter und leicht V-förmig. Ein vorspringendes Kinn fehlte. Die Körpergröße wird in der älteren Fachliteratur auf maximal 1,60 Meter geschätzt, da zunächst aussagekräftiges Skelettmaterial aus dem Bereich des Rumpfes und der Beine fehlte. Funde aus Afrika (Turkana-See) haben später dazu geführt, dass ausgewachsenen Individuen von Homo erectus inzwischen eine Körpergröße von 1,45 bis 1,80 Meter zugeschrieben wird.[14] Besonders vollständig erhalten und daher für den gesamten Körperbau besonders aussagekräftig ist das Fossil KNM WT 15000, genannt Turkana Boy.
Homo erectus verfügte bereits, wie bei Laetoli freigelegte, 1,51 bis 1,53 Millionen Jahre alte fossile Fußspuren belegen, sowohl über einen im Wesentlichen dem modernen Menschen entsprechenden Bau der Füße als auch über eine vergleichbare Form der aufrechten, zweibeinigen Fortbewegungsweise.[15]
Das Gehirnvolumen von Homo erectus vergrößerte sich im Verlauf der zwei Millionen Jahre seiner Existenz deutlich; u. a. deshalb variieren die Angaben zu seinem Gehirnvolumen selbst in der gleichen Region sehr stark: von 650 bis 1250 cm3 für ausgewachsene Individuen (zum Vergleich: Homo sapiens 1100 bis 1800 cm3).[16] In der Regel war das Volumen jedoch deutlich größer als das von Homo habilis (ca. 600 bis 700 cm3) oder Homo rudolfensis (ca. 750 cm3). Das Gehirnvolumen der Homo erectus-Funde aus Sangiran und Trinil wird mit rund 930 cm3 angegeben, das der Peking-Menschen mit rund 1060 cm3, das der Funde aus Ngandong mit rund 1150 cm3.[17]
2007 wurde in der Türkei ein Schädeldach von Homo erectus aus 500.000 Jahre altem Travertin geborgen, das deutliche Merkmale einer durch Tuberkulose ausgelösten Hirnhautentzündung (Leptomeningitis tuberculosa) aufwies.[18]
Ernährung
Unmittelbare fossile Belege für die Ernährungsweise der frühen Vertreter des Homo erectus sind nicht bekannt. Jedoch wird aus den im Vergleich mit den Australopithecinen kleineren Backenzähnen geschlossen, dass ihre Nahrung zumindest teilweise auch Früchte und andere, relativ weiche Bestandteile enthielt, häufiger aber Wurzeln, Fleisch und andere, kräftiges Kauen erfordernde Stoffe umfasste.[19] Welchen Anteil Jagdbeute an der Ernährung der frühen Homo erectus-Populationen hatte und welcher Anteil am Fleischkonsum als Aasfresser beschafft wurde, ist jedoch umstritten.[20]
Die späten Vertreter von Homo erectus – sie werden jedoch häufig zu Homo heidelbergensis gestellt – können bereits als Jäger und Sammler bezeichnet werden.
Kultur

Rekonstruktion eines Homo erectus im Westfälischen Museum für Archäologie Homo erectus lebte in jener Kulturstufe, die als Altsteinzeit bezeichnet wird. Schon für seine frühesten Vertreter ist der Gebrauch von Steingerät nachgewiesen, spätere Individuen nutzten auch das Feuer: möglicherweise bereits vor 790.000 Jahren.[21] Bemerkenswert ist, dass von den frühen Homo erectus-Fundstätten auf Java nur sehr primitive Steinwerkzeuge bekannt sind; keines dieser Werkzeuge wurde in dieser Region auf beiden Seiten bearbeitet.[22]
Die ersten Steinwerkzeuge (Chopper und Chopping Tools) gehören der Kulturstufe des Oldowan an. Vor etwa 1,5 Million Jahren wurde in Afrika die Stufe des Acheuléen erreicht. Vor etwa 500.000 Jahren kam diese Technikentwicklung auch nach Europa. Sie zeichnet sich durch beidseitige Klingen aus, die sehr sorgfältig gefertigt wurden. Man kann klare Grundtypen erkennen, die über viele Generationen gleich geblieben sind. Die Technik ist aufwändig zu erlernen. Daher ist anzunehmen, dass die afrikanischen und europäischen Homo erectus bereits über eine ausgeprägte Tradierung sowie die dafür notwendigen geistigen Fähigkeiten verfügten. Nach der Funktion kann man Hackmesser bzw. Schaber und größere Handäxte (sogenannte Faustkeile) unterscheiden. Die ältesten bisher in Asien entdeckten Faustkeile stammen aus China (Bose-Bassin) und sind 800.000 Jahre alt.[23]
Im Braunkohletagebau von Schöningen (Niedersachsen) wurden etwa 400.000 Jahre alte Holzspeere sowie ein beidseitig zugespitztes Wurfholz gefunden, die Homo erectus (bzw. Homo heidelbergensis) zugeordnet werden. Diese Schöninger Speere sind überwiegend aus Fichtenholz gefertigt und bis zu 2,5 m lang. Aufgrund der Schwerpunktlage dürften sie als Wurfspeere benutzt worden sein. Die Speere befanden sich auf einem Jagdlagerplatz zwischen den Überresten von mindestens 15 Pferden. Daher kann angenommen werden, dass der späte Homo erectus bereits die Großwildjagd zur Ernährung nutzte. Die sorgfältige Bearbeitung der Speere lässt auf eine gut ausgeprägte Kultur der Werkzeugherstellung schließen. Sehr ähnliche Funde wurden bei dem Fundplatz Bilzingsleben gemacht. Neben den Holzspeeren wurde hier ein Lagerplatz mit einfachen Wohnbauten und einem zentralen, gepflasterten Platz ausgehoben.
Zur Abgrenzung von anderen Arten der Gattung Homo
.png)
Ausbreitung und Fortentwicklung von Homo erectus, Sichtweise A: eine Art mit diversen regionalen Varianten Homo erectus galt unter Paläoanthropologen Jahrzehnte lang als jene Hominini-Art, die als erste alle drei Kontinente der Alten Welt besiedelte. Seit einigen Jahren wird dieser Konsens jedoch vor allem von US-amerikanischen Forschern infrage gestellt. „Die Kernfrage ist, ob die verschiedenen Homininen-Formen Asiens, Afrikas und Europas wirklich derselben oder aber unterschiedlichen Spezies zuzurechnen sind.“[24] Die aus Afrika, Asien und Europa bekannten Fossilien des Homo erectus weisen nämlich „eine große morphologische Streuung auf, die einige Forscher in mehrere Abstammungslinien aufspalten, während andere sie als eine einzige polytypische Art einstufen,“ [25]das heißt – ähnlich der Formenvielfalt bei Homo sapiens – als regionale Varianten einer Art.
Gestützt wird die u. a. von Berhane Asfaw, Meave Leakey und Tim White vertretene Auffassung, Homo erectus habe zahlreiche regionale Varianten aufgewiesen, durch Schädel- und Oberschenkelfunde auf der Halbinsel Bouri am Awash-Fluss in Äthiopien. [26]Hierzu gehören das Schädeldach BOU-VP-2/66, das am 27. Dezember 1997 geborgen und auf ein Alter von rund 1 Mio. Jahre datiert wurde. Aus diesem Fund wurde ein Schädelvolumen von 995 cm3 abgeleitet sowie – gestützt auch durch die Analyse mehrerer Oberschenkelknochen-Fragmente – eine enge morphologische Nähe zu gleich alten asiatischen Funden. Ein Vergleich mit jüngeren afrikanischen und asiatischen Funden ergab hingegen, dass in beiden Regionen vor 500.000 Jahren ein Formenwandel zu verzeichnen sei, der als Zeichen einer Artbildung gedeutet werden könne: In Afrika sei aus Homo erectus letztlich Homo sapiens hervorgegangen.[27]
Homo erectus und Homo ergaster
Ursache für diese sehr unterschiedlichen wissenschaftlichen Sichtweisen ist u. a., dass die Gestalt einiger zu Homo erectus gestellten afrikanischen Fossilien deutlich vom Typusexemplar dieser Art (Trinil II) abweicht.[28] Daraus leitete Bernard Wood 1992 die Berechtigung ab, die bis dahin zu Homo erectus gestellten homininen Fossilien aus der Zeit von vor ca. 1,9 bis 1,4 Mio. Jahren als Homo ergaster abzusondern. [29] Dies hat zur Folge, dass seitdem die afrikanischen Fossilien dieser Epoche einschließlich des Typusexemplars von Homo ergaster, des 1971 von Richard Leakey entdeckten Unterkiefers KNM-ER 992,[30] von einigen Forschern als Homo ergaster, von den meisten Forschern aber weiterhin – wie seit den 1950er-Jahren üblich – als Homo erectus bezeichnet werden.
Homo erectus und Homo heidelbergensis
Die Anfang der 1950er-Jahre noch bescheidene Anzahl von Hominini-Funden aus der Epoche vor 2 Millionen bis 500.000 Jahren hatte dazu geführt, dass all diese Funde der Art Homo erectus zugeordnet wurden. Dies galt auch für den Unterkiefer von Mauer, den sein Erstbeschreiber nach der nahen Universitätsstadt Homo heidelbergensis benannt hatte; diese Bezeichnung blieb jedoch bis in die 1980er-Jahre hinein – wenn überhaupt – allein auf den Unterkiefer von Mauer bezogen. Begründet wurde die Ablehnung einer weiteren Chronospezies damit, dass die stammesgeschichtliche Entwicklung in Europa gleitend von Homo erectus zum Neandertaler verlief; jede weitere Grenzziehung sei willkürlich, weswegen die Paläoanthropologen auf die Verwendung des Artnamens Homo heidelbergensis verzichteten. Diese Auffassung wird noch heute von europäischen Forschern vertreten, die daher auch den Unterkiefer von Mauer als lokale (europäische) Spätform des Homo erectus ausweisen; hierzu zählt unter anderem das Geologisch-Paläontologische Institut der Universität Heidelberg, das den Unterkiefer seit 1908 verwahrt und ihn als Homo erectus heidelbergensis ausweist.[31] Ähnlich argumentieren die französischen Entdecker des Homo erectus tautavelensis, die osteuropäischen Erforscher des Homo erectus bilzingslebensis und Alfred Czarnetzki, der Bearbeiter des Homo erectus reilingensis aus Reilingen.
.png)
Ausbreitung und Fortentwicklung von Homo erectus, Sichtweise B: die ältesten afrikanischen und die jüngeren europäischen Funde werden zu eigenen Chronospezies erhoben Eine zweite Gruppe von Forschern bezeichnet heute zwar die bis zu 1,8 Millionen Jahre alten afrikanischen Fossilien als Homo ergaster oder Homo erectus und leitet aus diesen die Besiedelung Europas und Asiens ab, benennt die europäischen Nachfahren dieser afrikanischen Auswanderer aber als Homo heidelbergensis. Ihren Publikationen zufolge verläuft die Entwicklung zunächst in Afrika von Homo ergaster zu Homo erectus und danach in Europa über Homo heidelbergensis zum Neandertaler. Diese Position wird beispielsweise aus der Datenbank des Human Evolution Research Centers (Berkeley) ersichtlich, die neben einem sehr alten Fund aus Israel ausschließlich europäische Funde der Art Homo heidelbergensis zuordnet. [32] Bernard Wood begründete die Zuordnung der europäischen Fossilien zu einer eigenen Chronospezies bereits 1984 damit, dass die Ähnlichkeit der asiatischen und der afrikanische Funde zwar „eindeutig und widerspruchsfrei“ („clear and unambiguous“) sei, die Übereinstimmung der europäischen Funde hingegen nicht überzeugend dargelegt wurde.[33]
Eine dritte Gruppierung von Forschern fordert schließlich eine noch weitergehende Änderung der Zuweisung von afrikanischen Fossilien zu einer Art der Gattung Homo. Von ihnen „wurde dieses umfassende Taxon aus chronologischen und geographischen Erwägungen aufgespalten“, Homo erectus wird von diesen Forschern seitdem „als Vertreter einer spezifisch ostasiatischen Stammlinie“ ausgewiesen.[34] Die ältesten, von anderen Wissenschaftlern weiterhin zu Homo erectus gestellten afrikanischen Fossilien werden von diesen Forschern als Homo ergaster bezeichnet, die jüngeren, unmittelbar an Homo ergaster anschließenden Fossilien aber als Homo heidelbergensis. Dieser Konvention zufolge entwickelte sich Homo ergaster bereits in Afrika zu Homo heidelbergensis fort, während aus Afrika nach Asien ausgewanderte Gruppen von Homo ergaster sich in Asien zu Homo erectus entwickelten.
Da sich die Paläoanthropologen bisher auf keine gemeinsame Konvention verständigen konnten, werden seit einigen Jahren – je nach Vorliebe der einzelnen Autoren – bestimmte Fossilien in Fachveröffentlichungen mal zur einen, mal zur anderen Art gestellt. Es wurde sogar eingewandt, dass selbst die Begrenzung des Homo erectus auf Asien noch viel zu unterschiedlich aussehende afrikanische Fossilien zu einer Art bündele. Die britische Paläoanthropologin Leslie Aiello wurde beispielsweise in der Fachzeitschrift Science zitiert, beim afrikanischen Homo heidelbergensis handele es sich um ein „Mülleimer-Taxon“; sie schlug vor, die Art Homo heidelbergensis europäischen Fossilien vorzubehalten und die afrikanischen Nachkommen von Homo ergaster zu einer bislang noch nicht benannten neuen Art zu erheben.[35]
Homo erectus, Homo rhodesiensis und Homo antecessor
.png)
Ausbreitung und Fortentwicklung von Homo erectus, Sichtweise C: nur die asiatischen Funde werden Homo erectus zugerechnet, die afrikanischen hingegen zu Homo heidelbergensis gestellt, und im Übergang von diesem zu Homo sapiens wird eine weitere Paläospezies etabliert Der Vorschlag jener Gruppierung von Forschern, die unmittelbaren afrikanischen Nachfahren von Homo ergaster als Homo heidelbergensis auszuweisen, hat zur Folge, dass der so definierte afrikanische Homo heidelbergensis als Vorfahre des Homo sapiens zu gelten hat. Wird hingegen in einer Fachveröffentlichung die alternative Lesart zugrunde gelegt, der zufolge Homo heidelbergensis eine erst in Europa aus Homo erectus entstandene Vorläuferart der Neandertaler ist, dann gibt es zwischen Homo heidelbergensis und Homo sapiens keine derart enge verwandtschaftliche Beziehung.
Vollends verwirrend wurde die Zergliederung des afrikanischen Homo erectus, als vorgeschlagen wurde, die in Afrika entdeckten unmittelbaren Vorfahren des Homo sapiens vom afrikanischen Homo heidelbergensis abzuspalten und als Homo rhodesiensis zu bezeichnen – ein Vorschlag, der sich bisher international nicht durchgesetzt hat. Diese rund 200.000 Jahre alten afrikanischen Fossilien werden heute gleichwohl – abhängig von der jeweiligen Forschergruppe – als früher Homo sapiens, archaischer Homo sapiens, Homo rhodesiensis, später Homo heidelbergensis oder später Homo erectus bezeichnet.
Spanische Forscher haben die Vielfalt der Benennungen 1994 schließlich um eine weitere bereichert: um den bisher nur aus Spanien bekannten Homo antecessor, dessen Einstufung als eigenständige Art allerdings umstritten ist. Einige spanische Forscher vertreten in ihren Publikationen zwar die Auffassung, Homo antecessor habe sich in Afrika aus Homo erectus entwickelt, sei dann in Europa eingewandert und habe sich hier zu Homo heidelbergensis fortentwickelt. Andere Forscher ordnen den älteren Teil der spanischen Fossilien hingegen Homo erectus zu, den jüngeren Homo heidelbergensis.
Siehe auch
Weblinks
- „The long journey of an ancient human ancestor“ Übersicht zu den Funden von Homo erectus in Asien, Afrika und Europa (auf Englisch)
- anthro.palomar.edu: Übersicht zur Entdeckungsgeschichte und zu anatomischen Merkmalen (auf Englisch)
- msu.edu Eine bebilderte Übersicht zu den asiatischen Funden von Homo erectus (auf Englisch)
Einzelnachweise
- ↑ In Grzimeks Tierleben wird Homo erectus als „Frühmensch“ bezeichnet; siehe dazu die „Systematische Übersicht“ in: Grzimeks Tierleben, Band 11 (=Säugetiere 2), dtv, 1979, S. 508. Die Gattung Australopithecus wird hier erläuternd als „Gattung Vormensch“ bezeichnet.
- ↑ Richard Leakey: The origin of humankind. Phoenix, a division of Orion Books Ltd., 1995, S. XIV
- ↑ Fred Spoor et. al.: Implications of new early Homo fossils from Ileret, east of Lake Turkana, Kenya. Nature, Band 448, 2007, S. 688–691; doi:10.1038/nature05986, vergl. dazu Meldung auf Spiegel.de
- ↑ von griech. ἄνθρωπος: „Mensch“ und píthēkos: „Affe“; Anthropopithecus war damals der Gattungsname für die Schimpansen
- ↑ Dubois publizierte seine Forschungsergebnisse anfangs quartalsweise in den Berichten des Minenwesens („Verslag van het Mijnwezen“) der niederländischen Kolonialverwaltung; die Benennung erfolgte im Bericht für das 3. Quartal 1892, (erschienen 1893), S. 10–14; vergl. dazu: Aleš Hrdlička: The skeletal remains of early man. Smithsonian Institution, 1930; Volltext
- ↑ Eugène Dubois: Pithecanthropus erectus: eine menschenähnliche Übergangsform von Java. Landes-Druckerei, Batavia, 1894
- ↑ Stephanie Müller u. a.: Sangiran II. Eine Schädelkalotte und ihre wissenschaftliche Deutung. Natur und Museum, Band 138, 2008, S. 30
- ↑ G. J. Sawyer, Viktor Deak: Der lange Weg zum Menschen. Lebensbilder aus 7 Millionen Jahren Evolution. Spektrum Akademischer Verlag, Heidelberg, 2008, S. 117
- ↑ Ernst Mayr: Taxonomic categories in fossil hominids. Cold Spring Harbor Symposia on Quantitative Biology 15, 1950, S. 109–118; Exzerpt
- ↑ Charles Darwin: The Descent of Man, and Selection in Relation to Sex. London, John Murray, 1871, Band 1, S. 199: „In each great region of the world the living mammals are closely related to the extinct species of the same region. It is therefore probable that Africa was formerly inhabited by extinct apes closely allied to the gorilla and chimpanzee; and as these two species are now man's nearest allies, it is somewhat more probable that our early progenitors lived on the African continent than elsewhere.“
- ↑ Winfried Henke, Hartmut Rothe: Stammesgeschichte des Menschen. Springer Verlag, 1999, S. 199
- ↑ Robert G. Franciscus, Steven E. Churchill: The costal skeleton of Shanidar 3 and a reappraisal of Neandertal thoracic morphology. Journal of Human Evolution, Band 42, 2002, S. 303–356; doi:10.1006/jhev.2001.0528
- ↑ Friedemann Schrenk: Die Frühzeit des Menschen. Der Weg zum Homo sapiens. C. H. Beck, 5., vollständig neubearbeitete und ergänzte Auflage, München 2008, S. 93 (C.H.Beck Wissen) ISBN 978-3-406-57703-1
- ↑ David Lordkipanidze et. al.: Postcranial evidence from early Homo from Dmanisi, Georgia. Nature, Band 449, 2007, S. 305–310; doi:10.1038/nature06134
- ↑ „The Ileret prints show that by 1.5 Ma, hominins had evolved an essentially modern human foot function and style of bipedal locomotion.“ Matthew R. Bennett et. al.: Early Hominin Foot Morphology Based on 1.5-Million-Year-Old Footprints from Ileret, Kenya. Science, Band 323, 2009, S. 1197 – 1201; doi:10.1126/science.1168132 10.1126/science.1168132
- ↑ Russell L. Ciochon, E. Arthur Bettis III: Asian Homo erectus converges in time. Nature, Band 458, 2009, S. 153–154; doi:10.1038/458153a
- ↑ G. J. Sawyer, Viktor Deak, S. 85 (H. habilis), S. 79 (H. rudolfensis), S. 115 und 124 (H. erectus)
- ↑ John Kappelman et al.: First Homo erectus from Turkey and implications for migrations into temperate Eurasia. American Journal of Physical Anthropology, Band 135, 2008, S. 110 – 116; doi:10.1002/ajpa.20739
- ↑ Peter S. Ungar et. al.: Dental microwear and diets of African early Homo. Journal of Human Evolution, Band 50, 2006, S. 78–95; doi:10.1016/j.jhevol.2005.08.007; vergl. dazu: abc.net.au: Homo erectus ate crunchy food.
- ↑ James F. O'Connell, Kristen Hawkes, Karen Lupo, N. G. Blurton Jones: Male strategies and Plio-Pleistocene archaeology. Journal of Human Evolution, Band 43, 2002, S. 831–872; doi:10.1006/jhev.2002.0604, Volltext, PDF; vergl. dazu „Frauen sorgten für Ernährung“: Überlegungen zur Ernährung von Homo erectus
- ↑ Naama Goren-Inbar et. al.: Evidence of Hominin Control of Fire at Gesher Benot Ya`aqov, Israel. Science, Band 304, 2004, S. 725–727; doi:10.1126/science.1095443
- ↑ G. J. Sawyer, Viktor Deak: Der lange Weg zum Menschen, S. 118
- ↑ Hou Yamei: Mid-Pleistocene Acheulean-like Stone Technology of the Bose Basin, South China. Science, Band 287, 2000, S. 1622–1626; doi:10.1126/science.287.5458.1622, Kurzfassung (auf Englisch)
- ↑ Winfried Henke, Hartmut Rothe: Stammesgeschichte des Menschen, S. 205
- ↑ Richard Potts et. al.: Small Mid-Pleistocene Hominin Associated with East African Acheulean Technology. Science, Band 305, 2004, S. 75–78; doi:10.1126/science.1097661
- ↑ Berhane Asfaw et. al.: Remains of Homo erectus from Bouri, Middle Awash, Ethiopia. Nature, Band 416, 2002, S. 317–320; doi:10.1038/416317a;. Vergl. dazu. science.orf.at: Homo erectus: Weltweiter Urahne des Menschen.
- ↑ Berhane Asfaw et. al. erwähnen in diesem Zusammenhang die Funde von Zhoukoudian (des Peking-Menschen), der allerdings bisher – anders als die europäischen Funde dieser und der späteren Epoche (= Neandertaler) – keiner eigenständigen asiatischen Art, sondern Homo erectus zugeordnet wird.
- ↑ Jeffrey H. Schwartz: Getting to Know Homo erectus. Science, Band 305, 2004, S. 53–54; doi:10.1126/science.1099989
- ↑ Bernard Wood: Origin and evolution of the genus Homo. Nature 355, 1992, S. 783–790; vergl. dazu auch: Bernard Wood: Early hominid species and speciation. Journal of Human Evolution, Band 22, 1992, S. 351–365 (Volltext, PDF)
- ↑ Unterkiefer KNM-ER 992, Abbildung
- ↑ geopal.uni-hd.de
- ↑ hercdb.berkeley.edu hercdb.berkeley.edu Maske zum Abfragen der bedeutenden Fossilien von Homo heidelbergensis
- ↑ Bernard Wood: The origin of Homo erectus. Cour. Forsch. Inst. Senckenberg, Band 69, 1984, S. 99–111.
- ↑ Robert Foley: Menschen vor Homo sapiens. Wie und warum unsere Art sich durchsetzte. Jan Thorbecke Verlag, 2000, S. 153.
- ↑ Ann Gibbons: A new face for human ancestors. Science Band 276, 1997, S. 1331–1333, doi:10.1126/science.276.5317.1331
.jpg)




.png)
.png)
.png)
Wikimedia Foundation.