- Haupthistokompatibilitätskomplexe
-
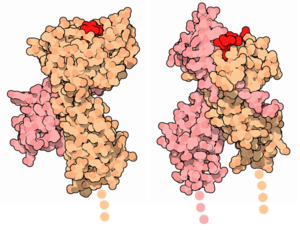 Oberflächenmodell der MHC-Klasse-I (Links) und MHC-Klasse-II-Proteinkomplexe (Rechts) mit den Untereinheiten (Orange und Rosa) und dem präsentierten Antigen (Rot).
Oberflächenmodell der MHC-Klasse-I (Links) und MHC-Klasse-II-Proteinkomplexe (Rechts) mit den Untereinheiten (Orange und Rosa) und dem präsentierten Antigen (Rot).Der Haupthistokompatibilitätskomplex (Abk. MHC von engl. Major Histocompatibility Complex) umfasst eine Gruppe von Genen bei Wirbeltieren, die Proteine codieren, welche für die Immunerkennung, die Gewebeverträglichkeit (Histokompatibilität) bei Transplantationen und die immunologische Individualität wichtig sind. MHC-Regionen finden sich in allen Wirbeltieren ab den Knorpelfischen (Haie, Rochen). Beim Menschen sind diese Gene auf dem kurzen Arm von Chromosom 6 zu finden. Die Genprodukte, die MHC-Proteinkomplexe, sind körpereigene Antigene auf der Oberfläche jeder Körperzelle, die immunologische Vorgänge regulieren. Diese Proteine kennzeichnen die Zellen als zum Körper gehörig und sind auf weißen Blutkörperchen (Leukozyten) leicht nachweisbar. Daher kommt auch die Bezeichnung HLA-System (Human Leucocyte Antigen) für das Regulationssystem der humanen Immunabwehr, dessen wichtigster Bestandteil der MHC ist.
Unter anderem werden vom Haupthistokompatibilitätskomplex die MHC-Klasse-I- und MHC-Klasse-II-Proteinkomplexe codiert, die für die Funktion des Immunsystems eine zentrale Rolle spielen.
Damit Antigene von T-Lymphozyten erkannt werden können, müssen sie zuvor aufbereitet und auf die vom MHC codierten Klasse-I- und Klasse-II-Rezeptoren, auf der Zelloberfläche präsentiert werden. Dieses Phänomen nennt man MHC-Restriktion.
Inhaltsverzeichnis
Die drei Klassen der MHC-Komplexe
Allgemein
Über den MHC-Klasse-I-Weg werden infizierte und entartete Zellen, die körperfremde Proteine herstellen, gezielt von T-Killerzellen (CD8+) identifiziert und anschließend eliminiert. Über den MHC-Klasse-II-Weg können T-Helferzellen (CD4+) die Produktion von spezifischen Antikörpern und die Aktivität von Phagozyten stimulieren, die Krankheitserreger in Körperflüssigkeiten inaktivieren und eliminieren (humorale Immunantwort). Das adaptive Immunsystem trennt also zwischen intra- und extrazellulären Pathogenen. Die MHC-Klasse-III-Komplexe sind Bestandteile des so genannten Komplementsystems, einem Teil des unspezifischen humoralen Immunsystems, welches zur Elimination von zellulären Antigenen (z.B. Bakterien) beiträgt. Die schwere Kette der MHC-Klasse-I-Komplexe sowie die α- und β-Untereinheit der MHC-Klasse-II-Komplexe treten beim Menschen in sehr vielen Allelen auf (genetischer Polymorphismus). Dies ist maßgebend für die Bedeutung des Haupthistokompatibilitätskomplexes bei der Gewebeverträglichkeit.
MHC-Klasse-I-Komplex
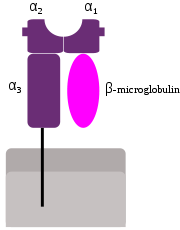 Schematische Darstellung des MHC-Klasse-I-Proteinkomplex.
Schematische Darstellung des MHC-Klasse-I-Proteinkomplex.Allgemein
Diese Proteinkomplexe finden sich auf der Oberfläche nahezu aller (kernhaltiger) Zellen des Organismus (außer Trophoblasten und Erythrozyten (kernlos)) und dienen der Antigenpräsentation für T-Killerzellen sowie dem Schutz gesunder Zellen vor einer Zerstörung durch Killerzellen (siehe auch: Missing self Hypothese). Der HLA-Klasse-I-Proteinkomplex besteht aus einer größeren an der Zellmembran verankerten Untereinheit, der schweren Kette (heavy chain, HC), und einer kleineren löslichen Untereinheit, dem β2-Mikroglobulin (β2M), sowie einem antigenischen Peptid. Im Proteinkomplex lassen sich vier Domänen unterscheiden: Die α-Untereinheit stellt drei Domänen (α1 bis α3), das β2-Mikroglobulin die vierte Domäne. Die Domänen (α1 und α2) bilden eine Grube, in der das Peptid gebunden ist. Diese Peptide werden in großer Zahl und Vielfalt durch das Proteasom von im Cytoplasma kontinuierlich synthetisierten Proteinen gebildet und stellen Spaltprodukte von ihnen dar. Das Immunsystem überwacht den Körper fortwährend auf das Vorliegen viraler Infektionen und entarteter Zellen, indem es überprüft, ob Zellen körpereigene oder fremde Proteine präsentieren und auf das vorhandensein von MHC-I-Proteinkomplexen. Die präsentierten Peptide stellen ein Abbild der in den Zellen synthetisierten Proteine dar. Die cytotoxischen T-Lymphozyten (CD8+ T-Zellen) sind so selektioniert, dass sie in der Regel mit ihrem T-Zell-Rezeptor nicht an Zellen binden, die ein Peptid präsentieren, das einem körpereigenen Protein entstammt. Dieses Phänomen nennt man Selbsttoleranz und schützt den Körper vor Angriffen des eigenen Immunsystem. Ist jedoch eine Zelle mit Viren infiziert oder von Mutationen betroffen und exprimiert somit neuartige Proteine, werden dem Immunsystem körperfremde Peptide als Teil des MHC-Klasse-I-Komplexes präsentiert und cytotoxische T-Lymphozyten aktiviert, die die betroffenen Zellen vernichten.
MHC-Klasse-I-Peptidbeladung
MHC-Klasse-I-Komplexe binden Peptide mit einer Länge von 8 bis 10 Aminosäuren in der peptidbindenden Spalte. Diese können nicht über den Spalt hinausragen - der Spalt ist an den Enden an konservierten Positionen durch Wasserstoffbrücken begrenzt - und sind über den freien Amino-Terminus und den freien Carboxyl-Terminus mit den invarianten Bereichen der Spalte assoziiert. Da die Peptide für eine stabile Bindung hydrophoben oder basischen Charakter haben müssen, werden auf dem MHC-Genlocus zusätzlich entsprechende Untereinheiten des Proteasoms codiert, die für eine häufigere Abspaltung von Peptiden mit diesen Charakteristika sorgen. Damit ist gewährleistet, dass alle cytosolischen Proteine (also sowohl körpereigene, als auch virale) in MHC-I taugliche Peptide gespalten und dort präsentiert werden können. Das Zusammensetzen des MHC-I findet im endoplasmatischen Retikulum (ER) statt, während die Proteine im Cytosol gespalten werden. Die peptidbindende Spalte bildet sich im ER und ist nie dem Cytosol ausgesetzt, daher müssen die cytosolischen Peptide in das ER transportiert werden. Da der MHC-I ohne gebundenes Peptid instabil ist, sind Peptidtransport und Faltung des MHC-I eng miteinander gekoppelt. Neu synthetisierte α-Untereinheiten des MHC-I, die ins ER transportiert werden, binden an das Chaperon-Protein Calnexin, welches diese Untereinheit in einem partiell gefalteten Zustand hält. Vermutlich ist auch die Oxidoreduktase ERp57 Bestandteil dieses Komplexes. Bindet nun β2-Mikroglobulin (β2M) an den diesen Komplex, dissoziiert der MHC-I-α-β2M Komplex vom Calnexin und wird in einem neuen Komplex, dem Peptid-Beladungskomplex (PLC, peptide loading complex) eingebettet. Weitere Bestandteile des PLC sind das Chaperon Calreticulin (dem Calnexin ähnlich, jedoch nicht membrangebunden), das auf den PLC spezialisierte Chaperon Tapasin, ERp57 und der Peptid-Transporter TAP (transporter associated with antigen presentation). Calnexin und Calreticulin binden beide das MHC-I über dessen Zuckerkette am Asparagin-Rest 86. Die P-Domäne von Calreticulin bindet außerdem an ERp57. Tapasin bindet kovalent an ERp57 und fungiert zudem noch als Bindeglied zu TAP. Über TAP werden die cytosolischen Peptide in das ER transportiert und lagern sich an MHC-I an. Ist die Bindung stabil genug, faltet sich der MHC-I vollständig und der PLC dissoziiert. Der so entstandene MHC-I-Peptid-Komplex wird in Vesikeln des sekretorischen Systems zur Oberfläche transportiert.
MHC-Klasse-II-Komplex
 Schematische Darstellung des MHC-Klasse-II-Proteinkomplex.
Schematische Darstellung des MHC-Klasse-II-Proteinkomplex.MHC-Klasse-II-Komplexe werden von spezialisierten Zellen des Immunsystems, so genannten professionellen Antigenpräsentierende Zellen (APC) (antigen presenting cells), präsentiert und von T-Helferzellen (CD4+ T-Zellen) erkannt. Zu den APCs gehören unter anderem die Monozyten und Makrophagen, interdigitierende follikuläre Zellen im Thymusmark, nicht phagozytierende follikuläre dendritische Zellen in Sekundärfollikeln von Lymphknoten und Milz, dendritische Zellen in Blut, Lymphe, Epidermis und anderen Geweben, phagozytoseaktive Zellen des Gefäßendothels, und B-Lymphozyten. Der HLA-Klasse-II-Proteinkomplex besteht aus zwei etwa gleich großen membranverankerten Untereinheiten, der α- und β-Untereinheit, sowie ebenfalls einem Peptid. Im Gegensatz zum Klasse-I-Komplex sind hier beide Untereinheiten in der Zellmembran verankert. Der Proteinkomplex besteht aus vier extrazellulären Domänen (α1 und α2 sowie β1 und β2). Auch dieser Komplex bildet eine Grube, die von der α1 und β1 Domäne geformt wird, und in der das Peptid gebunden ist. Die Peptide, die auf MHC-Klasse-II-Komplexen präsentiert werden, stammen von extrazellulären Proteinen ab, die z.B. durch rezeptorvermittelte Endozytose, unspezifische Makropinozytose oder Phagozytose Zugang zum sekretorischen Weg der APC gefunden haben. Wie die T-Killerzellen sind auch die T-Helferzellen so selektioniert, dass sie nur dann mit ihrem T-Zellrezeptor an einen MHC-Klasse-II-Komplex binden und damit aktiviert werden, wenn ein körperfremdes Antigen präsentiert wird.
Durch Endocytose werden die extrazellulären Proteine in Endosomen aufgenommen. Diese werden in der Zelle angesäuert, zum Beispiel durch Fusion mit einem Lysosom. In den Endosomen oder Lysosomen befinden sich Proteasen, die bei niedrigem pH-Wert aktiviert sind. Beispiele für solche Proteasen sind Cathepsin B, D, S und L. Sie bauen die Proteine zu Peptiden ab.
Wie viele andere Oberflächenproteine, bewegen sich die MHC-Klasse-II-Komplexe zunächst in das Endoplasmatische Retikulum. Damit sie hier nicht an eines der vielen Proteine binden, die sich im Endoplasmatischen Retikulum befinden, sind die MHC-Klasse-II-Komplexe mit einem Protein namens Invariante Kette, assoziiert. Ein Teil der Invarianten Kette liegt in der Bindungstasche und verhindert so die Bindung anderer Proteine. Die Invariante Kette formt mit einer anderen ein Homotrimer. Bis dieser stabile Zustand erreicht ist, sind die MHC-Klasse-II-Komplexe an Calnexin gebunden.
Nachdem der Trimer gebunden ist, verlassen die MHC-Klasse-II-Komplexe das ER. Das intrazelluläre Kompartiment mit den MHC-Klasse-II-Komplex fusioniert mit einem Endosom mit niedrigem pH-Wert. Durch die Proteasen wird die Invariante Kette mehrfach gespalten. Nach zwei Spaltungen bleibt nur noch ein kurzes Peptid, das in der Bindungstasche liegt, zurück. Dieses wird CLIP (class-II-associated invariant chain peptide) genannt und verhindert weiterhin die Bindunge eines Peptids. Um ein Peptid aus dem Endosom binden zu können muss der CLIP dissoziieren oder abgespalten werden. Dieser Vorgang wird durch ein weiteres MHC-Klasse-II-Komplex katalysiert, beim Menschen HLA-DM genannt. Es katalysiert die Abspaltung des CLIPs, bindet an leere MHC-Klasse-II-Komplexe und stabilisiert diese. HLA-DM selbst bindet keine Peptide, die Bindungstasche ist bei diesem Komplex geschlossen. Außerdem katalysiert HLA-DM den Austausch eines schwach gebundenen Peptids durch eines mit höherer Affinität zur Bindungsgrube.
Wie auch MHC-I-Komplexe sind MHC-II-Komplexe ohne gebundenes Peptid instabil und werden durch Proteasen schnell abgebaut. Auch sie präsentieren bei uninfizierten Zellen zelleigene Peptide auf der Oberfläche.
MHC-Klasse-III-Komplexe
Zu den Klasse-III-Komplexen gehören die Komplementfaktoren C2, C4 und Bf, sowie verschiedene Zytokine, wie beispielsweise der Tumornekrosefaktor (TNF). Im Gegensatz zu den anderen Klassen handelt es sich hierbei um Plasmaproteine, die an der unspezifischen Immunabwehr beteiligt sind.
Literatur
Weblinks
- Jennifer McDowall/Interpro: Protein Of The Month: Major histocompatibility complex. (engl.)


Wikimedia Foundation.