- Keimzentrum
-
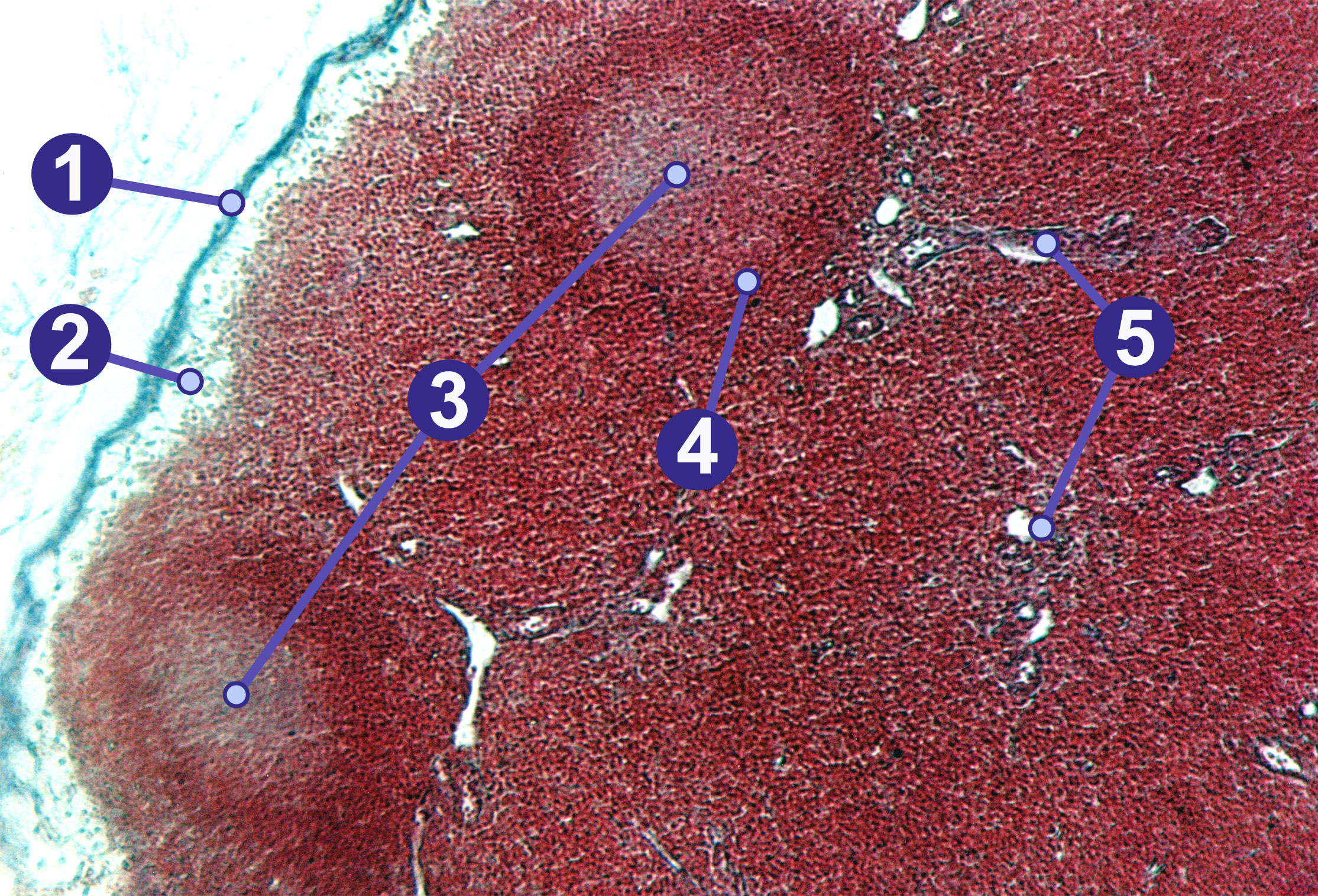
 Lymphknoten mit zwei Sekundärfollikeln: (1) Kapsel, (2) Randsinus, (3) Keimzentrum (Sekundärfollikel), (4) Parafollikulärer Raum, (5) Trabekel
Lymphknoten mit zwei Sekundärfollikeln: (1) Kapsel, (2) Randsinus, (3) Keimzentrum (Sekundärfollikel), (4) Parafollikulärer Raum, (5) Trabekel
Als Keimzentrum bezeichnet man die im Inneren eines Lymphfollikels nach Antigenkontakt entstehenden reaktiven Veränderungen, welches dadurch zu einem Sekundärfollikel wird. Das Keimzentrum ist eine lichtmikroskopisch sichtbare topografische Sonderung verschiedener Entwicklungsstadien von B-Zellen, die aufgrund immunologischer Vorgänge entsteht. Die dunkle innere Zone enthält sich vermehrende aktivierte B-Zellen (Zentroblasten). Diese wandern als Zentrozyten in die äußere helle Zone aus und werden an deren Rand selektiert. Um diese schließen sich eine Mantel- und Marginalzone an. In letzterer entwickeln sich die B-Gedächtniszellen.
Die Keimzentren werden auch als "risk zones" bezeichnet, da viele Lymphome von B-Lymphozyten abstammen.
Immunologische Prozesse vor der Keimzentrumbildung
Binden B-Zellen mit ihren B-Zell-Rezeptoren ein natives, lösliches Peptid-Antigen, werden die Rezeptoren samt gebundenen Antigenen in die Zelle aufgenommen. Die Antigene werden proteolytisch gespalten und als Peptid-Fragmente von 13 bis 17 Aminosäuren an MHCII-Moleküle gebunden. Diese MHCII-Peptid-Komplexe werden auf der Zellmembran präsentiert.
Diese B-Zellen erreichen dann die T-Zell-Bereiche sekundärer lymphatischer Organe. Dort befinden sich möglicherweise für dasselbe Antigen spezifische, bereits aktivierte T-Zellen. Diese erkennen mit ihren T-Zell-Rezeptoren die von den B-Zellen präsentierten MHCII-Peptid-Komplexe. Sie halten die B-Zellen fest, aktivieren sie und bilden mit ihnen in der T-Zell-Zone einen primären Focus, in dem die B-Zellen proliferieren.
Ein Teil der Nachkommen dieser B-Zellen bildet zwei bis drei Tage nach Aktivierung extrafollikuläre Foci (z. B. in der roten Pulpa der Milz oder in den Marksträngen der Lymphknoten), in denen sie proliferieren und zu Plasmazellen differenzieren. Sie sezernieren in den Foci IgM und IgG mit niedriger Antigenaffinität. Die meisten sind kurzlebig und besitzen eine 50%-Überlebensrate von drei bis fünf Tagen. Die Foci verschwinden nach etwa 14 Tagen.
Der andere Teil der B-Zellen wandert zusammen mit den T-Zellen, von denen sie aktiviert wurden, zu den B-Zell-Follikeln, wo sie noch an der Grenze zur T-Zell-Zone Keimzentren bilden.
Entstehung eines Keimzentrums

Bei der Entstehung eines Keimzentrums – Keimzentrumsreaktion – entsteht zuerst eine dunkle Zone, in der die aktivierten B-Zellen (Zentroblasten) einer massiven klonalen Expansion unterliegen. Dabei kann ein Klassenwechsel der konstanten Region der schweren Kette stattfinden und somatische Mutationen in die Gene für die variablen Regionen der leichten und schweren Ketten der Immunglobuline eingefügt werden. Dies kann zu einer Erhöhung oder Erniedrigung der Affinität der B-Zell-Rezeptoren zu ihrem Antigen führen.
Die B-Zellen wandern dann aus der dunklen Zone heraus in das Netzwerk der follikulären dendritischen Zellen und bilden die äußere, helle Zone des Keimzentrums. Der Eindruck einer „hellen Zone“ im histologischen Bild entsteht dadurch, dass aus den B-Zellen großkernige basophile B-Immunoblasten entstehen, die vom Rand in das Zentrum wandern.[1]
Die B-Zellen (Zentrozyten) teilen sich nun nicht mehr. Die follikulären dendritischen Zellen präsentieren Immunkomplexe aus Komplement, Antikörpern und Antigenen, die sie über Fc- und Komplementrezeptoren gebunden haben. Ist die Rezeptoraffinität der Zentrozyten zum Antigen zu gering, können sie mit ihren Rezeptoren nicht an das präsentierte Antigen binden. Ist ihre Affinität nicht zu gering, können sie diese Antigene binden und involvieren, abbauen und die Peptidfragmente, gebunden an MHCII, auf der Zelloberfläche exprimieren.
Die Zentrozyten wandern dann an den Rand der hellen Zone des Keimzentrums, wo sie auf die ebenfalls in gewissem Umfang klonal vermehrten CD4-T-Zellen treffen, denen sie die MHCII-Peptid-Komplexe präsentieren. Hier findet eine Selektion statt: Die B-Zellen, die eine niedrigere Affinität gewonnen haben und den T-Zellen kein Antigen präsentieren, erhalten kein Überlebenssignal – sie sterben apoptotisch. Nur B-Zellen, deren Affinität zum ursprünglichen Antigen hoch genug ist und Antigene aufgenommen haben und es den T-Zellen präsentieren, erhalten Überlebenssignale.
Sie entwickeln sich entweder zu Plasmazellen oder zu Gedächtniszellen. Die Entwicklung zu Plasmazellen findet über das Stadium der Plasmablasten statt. Plasmablasten sind noch teilungsfähig, können aber bereits Antikörper sezernieren. Plasmazellen sezernieren auch Antikörper, sind aber nicht mehr teilungsfähig. Die Entwicklung zu Gedächtnis-B-Zellen findet in der Marginalzone statt. Sie können auch in die dunkle Zone des Keimzentrums zurückkehren und erneut einer Keimzentrumsreaktion unterliegen. Ein Großteil der in Keimzentren entstandenen Plasmazellen wandert in das Knochenmark, wo sie zum Antigen hochaffine Antikörper der Klassen IgG, IgE und IgA sezernieren. Außerdem sind sie langlebig (teilweise länger als ein Jahr).
Während des ganzen Prozesses der Keimzentrumsbildung werden die durch die B-Zell-Follikel zirkulierenden B-Zellen immer mehr nach außen gedrückt und bilden eine das Keimzentrum umschließende Mantelzone.
Einzelnachweise
- ↑ Johannes W. Rohen, Elke Lütjen-Drecoll: Funktionelle Histologie. Schattauer Verlag, 4. Aufl. 2000, ISBN 9783794520442, S. 188.
Wikimedia Foundation.