- Röteln-Virus
-
Rötelnvirus 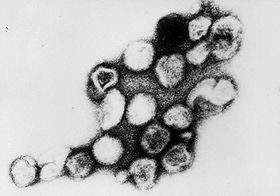 Rötelnvirus (TEM-Aufnahme)
Rötelnvirus (TEM-Aufnahme)Systematik Reich: Viren Ordnung: nicht klassifiziert Familie: Togaviridae Gattung: Rubivirus Art: Rötelnvirus Taxonomische Merkmale Genom: (+)ssRNA linear Baltimore: Gruppe 4 Symmetrie: ikosaedrisch Hülle: vorhanden Wissenschaftlicher Name Rubella virus (engl.) Taxon-Kurzbezeichnung RUBV Das Rötelnvirus (Rubella-Virus) ist der Erreger der als Kinderkrankheit bekannten Röteln, und ist bei Infektion in den ersten Schwangerschafts-Wochen der Verursacher der gefürchteten Rötelnembryopathie. Der Mensch ist der einzige bekannte Wirt des weltweit verbreiteten und mittels Tröpfcheninfektion übertragbaren Virus.
Das Rötelnvirus ist das einzige Mitglied der Gattung Rubivirus und gehört zur Familie der Togaviridae, deren Mitglieder typischerweise eine einzelsträngige RNA mit positiver Polarität als Genom besitzen, das von einem ikosaedrischen Kapsid umgeben ist. Das RNA-Genom im Inneren des Kapsids hat eine Länge von ungefähr 9'757 Nukleotiden und kodiert für zwei nichtstrukturelle Proteine sowie drei strukturelle Proteine.[1] Das Kapsidprotein sowie die beiden Hüllproteine E1 und E2 machen die drei strukturellen Proteine aus.
Die molekulare Grundlage für die Verursachung der Rötelnembryopathie ist noch nicht vollständig geklärt, aber in vitro Studien mit Zelllinien zeigten, dass das Rubella Virus einen apoptotischen Einfluss auf einzelne Zelltypen hat. Es gibt Hinweise auf einen von p53 abhängigen Mechanismus. [2]
Inhaltsverzeichnis
Morphologie
Die kugelförmigen Viruspartikel der Togaviridae haben einen Durchmesser von 50 bis 70 nm und sind von einer Lipidmembran (Virushülle) umgeben. In der Hülle sind als deutliche Spikes (Ausstülpungen) von 6 nm Höhe die Heterodimere der beiden viralen Hüllproteine E1 und E2 eingelagert.[3] Im Innern der Lipidhülle findet sich ein etwa 40 nm großes Kapsid.
Replikation
Togaviren heften sich über spezifische Rezeptoren an der Zelloberfläche an und werden durch ein sich ausbildendes Endosomvesikel aufgenommen. Bei neutralem pH-Wert außerhalb der Zelle bedeckt das E2-Hüllprotein das E1-Protein. Im Innern des Endosoms werden nun bei saurem pH die äußeren Domänen des E1-Proteins freigelegt und die Fusion von Endosommembran und Virushülle wird induziert. Somit gelangt das Kapsid in das Zytosol, zerfällt und gibt das Genom frei.
Die (+)ssRNA (positiv-einzelsträngige-RNA) dient zunächst nur der Translation der Nicht-Strukturproteine, die als großes Polyprotein synthetisiert und danach in einzelne Proteine zerschnitten werden. Die Sequenzen für die Strukturproteine werden erst über eine komplementäre (-)ssRNA als Matrize durch die virale RNA-Polymerase (Replikase) vervielfältigt und als eigene kurze mRNA translatiert. Diese kurze (subgenomische) mRNA wird beim Rubellavirus zusätzlich in das Virion verpackt. [4]Bei der Translation der Strukturproteine entsteht ebenfalls ein langes Polypeptid (110 Dalton), das in drei Stücke geschnitten werden muss. Dabei wird das Polyprotein endoproteolytisch in die Proteine E1, E2 und Kapsidprotein zerteilt. Bei E1 und E2 handelt es sich um Typ I Transmembranproteine deren Translokation in das Endoplasmatische Retikulum (ER) mit Hilfe einer N-terminalen Signalsequenz geschieht. Vom ER gelangt der als Heterodimer vorliegenden E1·E2-Komplex in den Golgi-Apparat, wo die Knospung (Budding) neuer Virionen stattfindet (im Gegensatz zu den Alphaviren, deren Knospung an der Plasmamembran stattfindet). Die Kapsidproteine hingegen bleiben im Cytoplasma und lagern sich an die genomische RNA an, mit der sie schließlich das Kapsid bilden.[5]
Kapsidprotein
Das Kapsidprotein (auch Protein C) hat verschiedene Funktionen. Die Hauptfunktionen sind die Bildung von Homooligomeren um das Kapsid zu formen, und die Bindung der genomischen RNA. Weiterhin ist es verantwortlich für die Aggregation der RNA im Kapsid, es interagiert mit den Membranproteinen E1 und E2 und bindet das menschliche Wirtsprotein p32, wobei diese Interaktion wichtig ist für die Vermehrung des Virus im Wirt.[6] Im Gegensatz zu Alphaviren macht das Kapsid keiner Autopreotolyse, sondern wird von der Signal-Peptidase vom Rest des Polyproteins abgetrennt. Die Herstellung des Kapsides erfolgt an Oberfläche der intrazellulären Membranen zeitgleich mit der Knospung des Virus.[7]
Literatur
- David M. Knipe, Peter M. Howley et al. (eds.): Fields´ Virology 4. Auflage, Philadelphia 2001
- C.M. Fauquet, M.A. Mayo et al.: Eighth Report of the International Committee on Taxonomy of Viruses, London San Diego 2005
Einzelnachweise
- ↑ Dominguez G, Wang CY, Frey TK: Sequence of the genome RNA of rubella virus: evidence for genetic rearrangement during togavirus evolution. In: Virology. 177, Nr. 1, July 1990, S. 225–38. PMID 2353453
- ↑ Megyeri K, Berencsi K, Halazonetis TD, et al: Involvement of a p53-dependent pathway in rubella virus-induced apoptosis. In: Virology. 259, Nr. 1, June 1999, S. 74–84. doi:10.1006/viro.1999.9757. PMID 10364491
- ↑ Bardeletti G, Kessler N, Aymard-Henry M: Morphology, biochemical analysis and neuraminidase activity of rubella virus. In: Arch. Virol.. 49, Nr. 2-3, 1975, S. 175–86. PMID 1212096
- ↑ Togaviridae- Classification and Taxonomy.
- ↑ Garbutt M, Law LM, Chan H, Hobman TC: Role of rubella virus glycoprotein domains in assembly of virus-like particles. In: J. Virol.. 73, Nr. 5, May 1999, S. 3524–33. PMID 10196241
- ↑ Beatch MD, Everitt JC, Law LJ, Hobman TC: Interactions between rubella virus capsid and host protein p32 are important for virus replication. In: J. Virol.. 79, Nr. 16, August 2005, S. 10807–20. doi:10.1128/JVI.79.16.10807-10820.2005. PMID 16051872
- ↑ Beatch MD, Hobman TC: Rubella virus capsid associates with host cell protein p32 and localizes to mitochondria. In: J. Virol.. 74, Nr. 12, June 2000, S. 5569–76. PMID 10823864
Weblinks
Togaviridae Information über Togaviridae (Stanford, eng.)
Wikimedia Foundation.