- CAM-Pflanzen
-
CAM-Pflanzen besitzen mit dem Crassulacean Acid Metabolism (Crassulaceen-Säurestoffwechsel) eine besonders an Trockenstandorte angepasste Form der Kohlendioxid (CO2)-Fixierung, die ähnlich derjenigen der C4-Pflanzen ist. Bei ihnen ist die CO2-Aufnahme und -Fixierung zeitlich getrennt von der CO2-Assimilation in der Photosynthese. Mit Hilfe des CAM können diese Pflanzen während der Nacht CO2 chemisch fixieren, um am Tag auf diese Reserve zurückzugreifen. Der Vorteil des CAM-Mechanismus ist, dass die Pflanze während der heißen Tagesstunden ihre Spaltöffnungen nicht zu öffnen braucht, um CO2 in ausreichender Menge zur Verfügung zu haben, wodurch sie bedeutend weniger Wasser durch Transpiration verliert. CAM-Pflanzen sind immer Sukkulenten; bekannte Vertreter sind Kakteengewächse, die Ananas, Kalanchoe, aber auch die heimischen Sedum- und Sempervivum-Arten. Benannt ist der CAM-Stoffwechsel nach der Familie der Crassulaceae (Dickblattgewächse), in der er häufig auftritt und in der er auch entdeckt wurde.
Inhaltsverzeichnis
Vorkommen
 Die Ananas ist eine der bekanntesten CAM-Pflanzen.
Die Ananas ist eine der bekanntesten CAM-Pflanzen.Der CAM-Mechanismus wurde in 25 Pflanzenfamilien nachgewiesen. Insgesamt kommt er in über 20.000 Arten vor. Neben der namensgebenden Familie der Dickblattgewächse (Crassulaceae) sind weitere Familien mit CAM-Arten:
- Kakteengewächse (Cactaceae)
- Bromeliengewächse (Bromeliaceae)
- Orchideen (Orchidaceae)
- Weinrebengewächse (Vitaceae)
- Liliengewächse (Liliaceae)
- Wolfsmilchgewächse (Euphorbiaceae)
- Aizoaceae
- Apocynaceae
- Korbblütler (Asteraceae)
- Didieraceae
- Agavaceae
- Portulacaceae
- Auch einige epiphytische Farne, wie Pyrrosia piloselloides und P. longifolia, sind CAM-Pflanzen.
Pflanzen mit CAM sind meist sukkulent. Wichtig ist dabei nicht die Organ-, sondern die Zellstruktur. Viele Arten kommen in periodisch trockenen Wuchsgebieten vor, besonders in Savannen und Wüsten der Tropen und Subtropen. Auch viele Epiphyten sind CAM-Pflanzen, so etwa 50 bis 60 % aller epiphytischen Orchideen und Bromelien. CAM-Stoffwechsel kommt aber auch bei einigen Wasserpflanzen vor, so Crassula aquatica und Littorella uniflora.
Durch sein Vorkommen in vielen, nicht miteinander verwandten Pflanzensippen muss der CAM-Stoffwechsel mehrmals unabhängig entstanden sein.
Mechanismus
Um in Wüstenregionen überleben zu können verfügen CAM-Pflanzen über Mechanismen, um die Schritte der CO2-Fixierung von jenen des Calvin-Zyklus zeitlich trennen zu können. Vermuteter Grund: eine Austrocknung wird dadurch vermieden, dass die Stomata nur nachts zur CO2-Aufnahme geöffnet werden müssen. Während also reine C4-Pflanzen eine räumliche Trennung (zwei Zelltypen, Mesophyllzellen und Leitbündelscheidenzellen) für die Fixierung und Metabolisierung von Kohlendioxid benutzen, wird beim "Crassulacean acid metabolism" (CAM) der folgende Tag-Nacht-Rhythmus eingehalten:
Nacht
In der Nacht erfolgt die CO2-Fixierung an Phosphoenolpyruvat (PEP) durch die PEP-Carboxylase ähnlich wie bei C4-Pflanzen. Das benötigte PEP wird meist durch Abbau von Stärke bereitgestellt, andere Pflanzenarten verwenden dafür lösliche Zucker wie Saccharose und Fruktane. CAM-Pflanzen müssen große Mengen an PEP bereitstellen können - so viel, dass eine Nachtration an CO2 fixiert werden kann. Zu diesem Zweck wird tagsüber der Stärke-Speicher aufgefüllt, der nachts über Pyruvat (Pyr) PEP zu liefern vermag. Diese Vorgänge werden unter C4-Pflanzen weiter beschrieben. Die PEP-Carboxylase befindet sich im Cytosol. Das von ihr gebildete Oxalacetat (OA) wird von der ebenfalls cytosolischen NAD-Malatdehydrogenase (MDH) zu Malat reduziert. Das Malat wird in einem energieverbrauchenden Prozess über einen Kanal in die Vakuole transportiert und dort gespeichert. Die Speicherung erfolgt in der protonierten Form des Malats, der Äpfelsäure, da bei den sehr sauren Bedingungen die Äpfelsäure überwiegend in undissoziierter Form vorliegt. Dies hat den Vorteil des dadurch geringeren osmotischen Wertes. Zusätzlich sinkt der pH-Wert der Vakuole durch die erhöhte Säurekonzentration nachts ab und erreicht pH-Werte von 3,0.
Tag
Am Tag wird dieser Weg formal umgekehrt. Das gespeicherte Malat wird wahrscheinlich über einen Carrier-vermittelten Transport aus der Vakuole ins Cytosol transportiert. Der Abbau des Malats und die Freisetzung des CO2 erfolgt wie bei C4-Pflanzen je nach Art auf verschiedene Weise, worin sich die unabhängige Ausbildung des CAM in verschiedenen Taxa widerspiegelt. Die üblichen Wege sind das NADP-Malat-Enzym (bei Cactaceae, Agavaceae), das NAD-Malat-Enzym (bei Crassulaceae) und die Phosphoenolpyruvat-Carboxykinase (z.B. bei Bromeliaceae). Das entstehende CO2 wird in den Calvin-Zyklus eingeschleust, das Pyruvat dient über die Zwischenstufen Phosphoenolpyruvat und 3-Phosphoglycerat ebenfalls wieder der Kohlenhydratbildung. Da der Großteil der gebildeten Kohlenhydrate dem Auffüllen der in der Nacht zuvor verbrauchten Stärkevorräte dient, ist der Gewinn der CAM-Photosynthese gering. Zusätzlich ist das Ausmaß der Malat-Speicherung durch die Größe der Vakuole beschränkt. Daher sind auch viele CAM-Pflanzen sukkulent.
Eine Fixierung des CO2 durch die PEP-Carboxylase am Tag wird dadurch verhindert, dass das Enzym am Tag in eine inaktive (= dephosphorylierte) Form überführt wird, die durch das aus der Vakuole strömende Malat zusätzlich inhibiert wird. Das für die Regulation verantwortliche Enzym, die PEP-Carboxylase-Kinase, unterliegt einer strikten circadianen Rhythmik seiner Genexpression und, bedingt durch einem raschen Abbau, auch seiner Gesamtaktivität.
Durch den Auf- und Abbau des Malats ergeben sich starke Änderungen des pH-Wertes von pH 5 am Tag zu pH 3 in der Nacht. Man spricht daher vom diurnalen Säurerhythmus.
Übersicht: Crassulacean Acid Metabolism
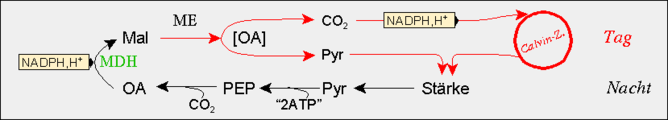
Crassulacean acid metabolism (CAM) Konsequenzen des CAM
Der ökologische Vorteil des CAM ist, dass die CO2-Aufnahme und damit die Öffnung der Stomata in der Nacht erfolgt, während sie am Tag geschlossen bleiben und somit die Verdunstung stark reduziert wird. Daraus ergibt sich ein äußerst geringer Wasserbedarf pro Trockengewicht von 50 bis 55 mL g-1 im Gegensatz zu 450 bis 950 bei C3-Pflanzen. Die Öffnung der Stomata in der Nacht bedingt, dass CAM-Pflanzen besonders häufig in Gebieten mit tiefen Nacht-Temperaturen und hoher Luftfeuchtigkeit (Nachtnebel) auftreten.
Bei besonders starker Austrocknung bleiben die Stomata auch nachts geschlossen und es wird nur das durch die Atmung freigesetzte CO2 refixiert. Dies ermöglicht es den Pflanzen, auch lange Perioden extremer Trockenheit zu überdauern.
Der erhöhte Energiebedarf für die Kohlenstoffassimilation im Vergleich zu den C3-Pflanzen fällt angesichts der hohen Sonneneinstrahlung an den meisten Standorten der CAM-Pflanzen kaum ins Gewicht. Zudem wird die Lichtsättigung der Photosynthese meist erst bei höheren Lichtintensitäten erreicht als bei C3-Pflanzen.
Viele CAM-Pflanzen können zwischen C3 und CAM wechseln. Bei guter Wasserversorgung betreiben sie C3-Photosynthese mit tagsüber geöffneten Stomata. Bei Trockenstress wechseln sie auf CAM, um Wasserverluste zu minimieren. Diesen induzierbaren CAM findet man etwa bei Mesembryanthemum. In der Gattung Kalanchoe sind sowohl Arten mit fakultativem (induzierbarem) CAM wie auch obligate CAM-Pflanzen (konstitutiver CAM) vertreten.
Nachweis
Es gibt mehrere Charakteristika, deren Nachweis notwendig ist, um eine Pflanze dem CAM zuordnen zu können:
- Der diurnale Säurerhythmus
- Die nächtliche CO2-Fixierung kann durch Gaswechselmessungen bestätigt werden. Bei den Bromeliaceen kann man auch die Spaltöffnungsweite bei Tag und bei Nacht messen: Man streicht (am besten zweischichtig) farblosen Nagellack über die Blattunterseite - nur ca. auf einen Quadratzentimeter - und betrachtet diesen anschließend unter dem Mikroskop. Sind die Spaltöffnungen über Nacht geöffnet und am Tag geschlossen, liegt der Verdacht nahe, dass es sich um eine CAM-Pflanze handelt.
- Hohe Aktivität der PEP-Carboxylase
- Hoher Wassergehalt der Zellen, bzw. große Vakuolen, um die hohen Malat-Gehalte speichern zu können.
Isotopendiskriminierung
Wie auch bei C4-Pflanzen (siehe dort), diskriminiert auch die PEP-Carboxylase in CAM-Pflanzen nicht so stark gegen 13C wie RubisCO. Daher entspricht ihr Isotopenverhältnis (δ-13C) bei CO2-Dunkelfixierung dem von C4-Pflanzen, bei Lichtfixierung von externem CO2 jedoch dem von C3-Pflanzen. Bei zunehmendem Trockenstress nimmt der Anteil der Dunkelfixierung zu, die CAM-Pflanze wird reicher an 13C. Der δ-13C-Wert einer CAM-Pflanze ist daher ein gutes Maß für die Dürrebelastung an ihrem Wuchsort, da er eine Aussage über das Ausmaß des CAM-Stoffwechsels im Verhältnis zum normalen C3-Stoffwechsel erlaubt.
Literatur
- I.P. Ting: Crassulacean Acid Metabolism. In: Annual Review of Plant Physiology 36, 1985, S. 595-622
- K. Winter, J.A.C. Smith (Hrsg.): Crassulacean Acid Metabolism. Ecological Studies 114, 1996. Springer, Berlin
- J.B. Hafke, Y. Hafke, J.A.C. Smith, U. Lüttge, G. Thiel: Vacuolar malate uptake is mediated by an anion-selective inward rectifier. In: The Plant Journal 35, 2003, S. 116-128
- U. Lüttge: Ecophysiology of Crassulacean Acid Metabolism (CAM). In: Annals of Botany 93, 2004, S. 629-652
Weblinks


Wikimedia Foundation.