- Muller’s ratchet
-
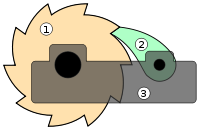 Die Sperrklinke (2) verhindert das Rückwärtsdrehen des Sperrrades (1). Genauso lässt sich die Muller-Ratsche nur in die „Richtung“ von zusätzlichen, schädlichen Genmutationen drehen.
Die Sperrklinke (2) verhindert das Rückwärtsdrehen des Sperrrades (1). Genauso lässt sich die Muller-Ratsche nur in die „Richtung“ von zusätzlichen, schädlichen Genmutationen drehen.
Muller's ratchet (nach Hermann Joseph Muller, dtsch. Muller-Ratsche oder -Sperrklinke) ist ein Konzept in der Evolutionsbiologie, das erklärt, warum Organismengruppen oder Populationen bestimmte Merkmale oder deren Entwicklungsrichtungen beibehalten und nicht verlassen oder umkehren können.
Man spricht davon, dass das Merkmal in der Population oder der Organismengruppe durch die Sperrklinke "gefangen" ist. Es kann sich nur in die Merkmalsausprägung hinein, nicht jedoch wieder heraus entwickeln, da der Rückweg versperrt ist. Bekannte Beispiele sind der extrem ausgeprägte Sexualdimorphismus der Paradiesvögel oder auch die sexuelle Fortpflanzungsweise selbst, die fast alle mehrzelligen Organismen betreiben.
Ein anschauliches Modell beschreibt einen geometrischen Raum, der eine große Menge Punkte enthält, von denen ein jeder jeweils eine mögliche Merkmalskombination repräsentiert, so dass in diesem Raum alle existierbaren organismischen Formen enthalten sind. Evolutionäre Entwicklungslinien entsprechen dann Pfaden durch diesen Raum. Dabei gibt es Pfade, die in bestimmte Richtungen nicht verlassen werden können bzw. Bereiche in diesem Raum sind von bestimmten, sperrklinkenführenden Pfaden unerreichbar. So hat sich erwiesen, dass sexuell fortpflanzende Entwicklungslinien im Verlaufe ihrer Evolution nie Raumbereiche durchqueren, in denen rein asexuell fortpflanzende Formenkreise liegen, obwohl dies biologisch möglich wäre. Auch viele sexualdimorphistische Merkmale werden meist nicht mehr aufgegeben, sobald sie einmal erworben sind.
Wie mechanische Sperrklinken sind auch evolutionäre Muller-Ratschen sehr stabil. Abstammungslinien (Populationen, Arten, höhere Taxa) mit solchen Ratschen sterben unter sich verändernden Umweltbedingungen, die eine Fortführung der eingeschlagenen Entwicklungsrichtung verhindern, mangels schlechter Anpassungsfähigkeit aus. Dies trat im Laufe der Stammesgeschichte bei Arten mit starken sexualdimorphen Merkmalen regelmäßig ein, während für die sexuelle Fortpflanzungsweise selbst bislang keine Bedingungen eingetreten sind, die sie blockieren. Sollte dies passieren, so gibt es nur bei einigen wenigen sexuell fortpflanzenden Linien die Möglichkeit, sich anzupassen.
Weil Merkmale stets durch viele Gene gesteuert werden und somit auf verschiedenen Ebenen gleichzeitig wirksam sind, gelten Muller-Ratschen als evolutionär überaus stabil und schwer überwindbar.
Beispiel Sexualdimorphismus bei Vögeln
Viele Vögel, wie beispielsweise Paradiesvögel, weisen in Morphologie und Verhaltensbiologie einen ausgeprägten Sexualdimorphismus auf, der durch die bei der Partnerwahl sehr wählerisch vorgehenden Weibchen verursacht wird. Die Weibchen wählen Männchen sehr genau nach Schönheitsmerkmalen aus und gestalten so im Verlauf der Phylogenese immer aufwändigere männliche Körper- und Verhaltensformen. Analog dazu werden die Weibchen immer wählerischer. Selten kommt es dazu, dass Körperform oder Verhalten vereinfacht wird und wenn, dann nur in jeweils einem Aspekt, wobei die Entwicklungsrichtung nach höherer Komplexität sofort erneut aufgegriffen und fort geführt wird.
Dies beginnt evolutionär zunächst durch eine Gewichtung eines vorerst praktischen Merkmals, wie längere Schwanzfedern bei Männchen durch Bevorzung durch Weibchen. Sobald die Federn eine Länge erreicht haben, ab der sie wieder hinderlich sind und zu häufigerer Selektion durch Fressfeinde führen, pegelt sich ein Längen-Optimum ein, wobei die Selektion dann auf komplexere Verschnörkelungen, aufwendige Farb- und Formgebung ausweicht. Ab diesem Zeitpunkt beginnt die Ratsche zu wirken und versperrt den Rückweg. Es kommt zu einem Wettrüsten, das bei den Männchen zu komplexeren Formen, bei den Weibchen zu immer größerer Wählerischkeit führt. Die Männchen müssen, um sich erfolgreich fortpflanzen zu können, ein überaus aufwendiges Balzverhalten betreiben und ihren kostenintensiven Körper unterhalten; an der Aufzucht der Jungen oder an der Versorgung der Weibchen hingegen beteiligen sie sich nicht. Sie stecken praktisch ihre gesamten verfügbaren Ressourcen in Partnerwerbung und werden aufgrund ihrer auffälligen Erscheinung, Gleichgültigkeit angesichts von Fressfeinden oder körperlichen Behinderungen wie Schmuckfedern oder ausladenden Anhängseln weitaus häufiger selektiert als Weibchen oder Männchen, die diese handicaps nicht haben. Andererseits können sich Weibchen nur erfolgreich fortpflanzen, wenn sie "die schönsten" Männchen wählen, denn ihre eigenen Söhne müssen ebenfalls schön sein, um hohe Chancen bei anderen Weibchen zu haben.
Man könnte nun annehmen, dass dieses Wettrüsten durch eine Änderung der Umweltbedingungen durchbrochen werden kann. Aber auch das ist nicht der Fall und die Evolution kann bei diesen Vögeln nicht zu einer Vereinfachung des Dimorphismus oder zu einer Rückkehr in die partnerschaftliche Lebensweise führen.
Die Sperrklinke besteht darin, dass jede Mutation, die ein Weibchen dazu veranlasst, seinen Geschlechtspartner weniger nach Schönheit, sondern stärker nach praktischen Eigenschaften zu erwählen, von der Enkel-Generation sofort wieder aus dem Genpool entfernt wird. Aufgrund reiner numerischer Überlegenheit von älteren Genen können sich alternative Mutationen nicht durchsetzen, auch wenn sie zu einer Anpassungsverbesserung in der Umwelt führen würden. Dabei wirkt die Ratsche auf mehreren Ebenen der Entwicklungsrichtung. Beispiele:
- Ein Männchen erhält eine Mutation, die ihm ein praktischeres, aber weniger schönes Körpermerkmal beschert. Zum Beispiel ein einfacherer Körperbau mit höherer Fitness gegenüber Fressfeinden oder Verletzungen. Es lebt vielleicht länger, hat aber geringen Fortpflanzungserfolg bei den Weibchen: das mutierte Gen wird ausselektiert.
- Ein Weibchen A erhält eine Mutation, die es veranlasst, Männchen mit praktischeren einfacheren Formen zu wählen. Die Söhne von A haben dadurch ebenfalls einfachere Körperformen und werden nach (1) von jenen Weibchen ausselektiert, die die Mutation von A noch nicht haben.
Aufgrund reiner numerischer Überlegenheit der nicht mutierten Individuen können sich Mutationen nicht durchsetzen. Ändern sich die Umweltbedingungen, kann sich die Abstammungslinie nur dann halten, wenn zeitgleich Mutationen nach (1) und (2) in ausreichender Zahl und "richtiger" Abfolge ereignen. Dies tritt aufgrund der Wahrscheinlichkeitsgesetze jedoch nicht ein.
Auch die Bildung von kleinen Inselpopulationen, in denen eine Mutation rasch zur Majorität kommen kann, scheidet als Überwindungsmechanismus der Sperrklinke aus. Dies kommt daher, weil Merkmale von vielen Genen gesteuert werden. Eine einzelne, erfolgreich durchgesetzte Mutation führt nur zum Zurückdrehen der Sperrklinke um "einen Zahn". Sie rastet jedoch unter dem Druck der übrigen Gene sofort wieder ein und führt zu einer im Vergleich zur Mutterpopulation lediglich abweichenden Entwicklung (Artenbildung), die jedoch in dieselbe spezialisierende Richtung führt. Um die Ratsche auf ein Maß zurück zu drehen, auf dem die Art wieder mehr Entwicklungsfreiheit genießt, wären zahlreiche, einander abfolgende Inselbildungen mit jeweils praktischen Mutationen nötig, was in der Natur nicht vorkommen kann. Die betreffende Art kann die Ratsche so nicht überwinden. Auch einander abfolgende Rückmutationen sind faktisch völlig unmöglich.
Beispiel der Sexuellen Fortpflanzung
Auch die Sexuelle Fortpflanzungsweise selbst ist in einer Muller-Ratsche gefangen, d.h. sexuell fortpflanzende Organismengruppen oder Abstammungslinien können sie nicht aufgeben.
Obwohl die asexuelle Fortpflanzung einen numerischen Vorteil verspricht (nicht nur Weibchen, sondern alle Individuen erzeugen Nachkommen) und z.B. die Fähigkeit zur Parthenogenese biologisch bei vielen Organismengruppen nachgewiesen ist (auch bei Wirbeltieren, Reptilien) kann eine sexuell fortpflanzende Entwicklungslinie nicht in eine asexuelle Entwicklungsrichtung einmünden.
Bei der sexuellen Fortpflanzung gibt es zwei Geschlechter, das männliche und das weibliche. Jedoch bringen nur Weibchen Junge zur Welt, weshalb Männchen in jeder sexuell fortpflanzenden Population (außer ihrer Funktion als Pollen-/Spermaspender) schlicht überflüssig sind – da sie etwa den Weibchen Nahrung und Lebensraum wegnehmen. (Nach Darwin konkurrieren die Mitglieder der selben Art am härtesten mit einander, da sie um exakt dieselben Ressourcen kämpfen.)
Einer der Auswege aus dem Dilemma, den Nutzen sexueller Fortpflanzung zu erklären, ist die Muller's ratchet bei asexuell fortpflanzenden Lebewesen:
- Selektionsdruck kann nur derzeit existierende Individuen in ihrer Reproduktionsfähigkeit (Fitness) begünstigen. Dies heißt, dass Individuen mit der derzeit geringsten Zahl an schädlichen Genmutationen begünstigt werden.
- Hier kommt die Ratsche (engl. ratchet) ins Spiel: Selektion kann kein Individuum mit der Zahl n-1 schädlichen Mutationen begünstigen, da n schädliche Mutationen bereits das Minimum der Population darstellen (siehe unten zu Rückmutation).
- Die Folge ist, dass die Selektion die Zahl der schädlichen Mutationen im Erbgut asexuell fortpflanzendender Individuen nie senken kann (da Individuen mit n Schadmutationen den größten Reproduktionserfolg haben). Jedoch werden sie durch Neumutationen langsam, aber stetig erhöht.
Bei sexuell fortpflanzenden Organismen findet aber genetische Rekombination und eine Neuverteilung der Chromosomen aufs Erbgut von Nachkommen statt. Dies schafft die Möglichkeit der Zeugung von Nachkommen mit n-x Mutationen aus Eltern, die n oder mehr Mutationen in ihrem Erbgut tragen.
Die einzige Möglichkeit, der Muller's ratchet zu entgehen, besteht auch hier in der sehr seltenen Rückmutation eines mutierten DNA-Basenpaars. (Rückdrehen der Sperrklinke um einen Zahn) Eine Neumutation geschieht während der DNA-Replikation mit einer Wahrscheinlichkeit von etwa 1:1 Milliarde – damit eine Mutation korrigiert wird, muss am selben DNA-Basenpaar eine neuerliche (Rück-)Mutation stattfinden, welche dazu noch das richtige (von vier möglichen) Basenpaaren erzeugt; damit ist der Vorgang Neumutation-Rückmutation extrem unwahrscheinlich (Wahrscheinlichkeit 1:1 Milliarde mal 1:1 Milliarde). Angesichts dessen, dass asexuell fortpflanzende Lebewesen (oft Bakterien) ein Erbgut von weit unter 1 Milliarde Basenpaare haben, spielt die Rückmutation also eine vernachlässigbare Rolle.
Siehe auch
- en:Evolution of sex in der englischen Wikipedia; erklärt ausführlich die Vor- und Nachteile der sexueller und nichtsexueller Fortpflanzung.

Wikimedia Foundation.