- Energiebereitstellung des Muskels
-

Dieser Artikel wurde aufgrund von formalen und/oder inhaltlichen Mängeln in der Qualitätssicherung Biologie zur Verbesserung eingetragen. Dies geschieht, um die Qualität der Biologie-Artikel auf ein akzeptables Niveau zu bringen. Bitte hilf mit, diesen Artikel zu verbessern! Artikel, die nicht signifikant verbessert werden, können gegebenenfalls gelöscht werden.
Lies dazu auch die näheren Informationen in den Mindestanforderungen an Kurzartikel der Biologie.
Inhaltlich überprüfen bitte --Pittimann besuch mich 17:31, 3. Feb. 2009 (CET)
Die Muskelarbeit ist ein entscheidender Faktor im täglichen Leben. Sie ist dafür verantwortlich dass der Mensch jegliche Form von Arbeit, welche immer in Verbindung mit einer Muskelkontraktion steht, leisten kann. Die Muskelzelle ist gekennzeichnet und dafür ausgelegt, dass die ihr zugrundeliegende Struktur eine ganz bestimmte Leistung erbringen kann und muss, nämlich diese Kontraktion welche bis zu 50mal pro Sekunde (vgl. Weineck 2006, 99), abhängig von dem Muskelfasertyp, durchgeführt werden kann. Hierbei wird jedes einzelne Sarkomer im Muskel um bis zu 30 % seiner Ruhelänge verkürzt (vgl. Heck 1990, 21). Für diese mechanische Arbeit ist das Myosinmolekül auf einen chemischen Energieumsatz angewiesen, welcher an Größe von keinem anderen Zellverband erreicht wird. Der Energieumsatz der Muskelzelle hat sowohl für die Erhaltung und Wandlung zu dienen als auch für die energetischen Leistungen. Unzweifelhaft besteht ein enges Zusammenspiel zwischen der Struktur der Muskelzelle und ihren Umsätzen.
Bei der Energiebereitstellung für die Muskelzelle muss zwischen zwei grundlegenden energieliefernden Vorgängen unterschieden werden. Einmal gibt es die exogenen auf dem Blutweg zugeführten energiereichen Substrate und andererseits die endogenen Substrate. Die Energiebereitstellung der Muskelzelle und somit auch ihre Leistungsfähigkeit sind abhängig von der Mobilisierung, dem Transport sowie dem Abbau energiereicher Substrate unter ATP-Gewinnung. Da der intrazelluläre ATP-Vorrat aber nur sehr begrenzt und gering ist, bedient sich die Muskelfaser verschiedener Wege der ATP-Resynthese. Hierbei unterscheidet man zwischen drei energieliefernden Stoffwechselwegen. Es gibt die anaerob-alaktazide Energiebereitstellung welche anaerob (ohne Zuhilfenahme von Sauerstoff) und alaktazid, also ohne (nennenswerte) Milchsäureproduktion (Laktatproduktion), verläuft. Der zweite Resyntheseweg ist der anaerob-laktazide, welcher wie der Name schon sagt zwar auch anaerob verläuft, aber mit einer Laktatproduktion verbunden ist. Des Weiteren gibt es noch den Weg der aeroben Energiebereitstellung. Hierbei arbeitet die energieliefernde Reaktion aerob, also unter Beihilfe von Sauerstoff.
Inhaltsverzeichnis
Energiestoffwechsel
Energiereiche Phosphate
Damit ein Muskel Kontraktionsarbeit verrichten kann, braucht er Energie, welche aus chemischen Reaktionen stammt. Demnach ist der Stoffwechsel des Muskels durch die Anpassung an den Kontraktionsvorgang gekennzeichnet (vgl. Keul/Doll/Keppler 1969, S. 11). Schon 1841 beobachteten Berzelius, 1877 Du Bois-Reymond, u.a., dass ein enger Zusammenhang zwischen Muskelkontraktion und Stoffwechsel herrscht und zeigten, dass mit der Kontraktionsarbeit der Muskelzelle eine Laktatbildung verbunden ist. Diese Beobachtungen brachten 1914 Pranas und Wagner dazu, die dem Abbau des Glykogendepots des Muskels zugrundeliegende Milchsäurebildung als unmittelbare Energiequelle der Kontraktionsarbeit zu sehen. Gestützt wurde diese Aussage durch die Tatsache, dass die Bildung des Laktats aus Glykogen mit einer Energiefreisetzung verbunden ist. Jedoch wurden durch die Versuche von Hoett und Marks 1926 die Abhängigkeit des Kontraktionsvorganges von der Laktatbildung in Frage gestellt und von Clark und Eggleton 1932 bewiesen, dass eine Milchsäurebildung erst bei einer längeren Muskelarbeit auftritt. Nachdem die Substanz Kreatinphosphat im Muskel entdeckt wurde, wurde ein Zusammenhang zwischen dem Kreatinphosphatstoffwechsel mit den Kontraktionsvorgängen in der Muskelzelle geschlossen, da während des Kontraktionsvorganges ein Abfall und in der Erholungsphase ein Wiederanstieg des Kreatinphosphats zu beobachten war. Während diese und weiter Erkenntnisse schon Zweifel an der Abhängigkeit des Kontraktionsvorganges und der Milchsäurebildung aufkommen und eine wichtige Stellung des Kreatinphosphats erkennen ließen, wurde durch Lunddgaard 1931 die Milchsäuretheorie der Muskelkontraktion eindeutig widerlegt. Durch einen geeigneten Versuch unter Zuhilfenahme eines von ihm vergifteten Muskels mit einem besonderem Substrat, welches eine Laktatbildung verhindert, dieser aber dennoch kontraktionsfähig und die geleistete Arbeit proportional zum Abbau des Kreatinphosphats war, stellte er die Milchsäuretheorie unwiderruflich ein. In der Laktatbildung wurde so also ein anaerober Resyntheseweg für das Kreatinphosphat gesehen. Des Weiteren wurde festgestellt, dass unter aeroben Bedingungen eine Milchsäurebildung aus bleibt und die Resynthese des Kreatinphosphats durch oxydative Reaktionen stattfindet. Schließlich entdeckte Lohmann 1931 dann das Adenosintriphosphat (ATP). Durch diese Entdeckung wurde jetzt auch das Kreatinphosphat als unmittelbare Energiequelle der Muskelarbeit in Frage gestellt, da in der Folgezeit dem ATP eine sehr wichtige Bedeutung als Koenzym, Regulationsfaktor im Zellstoffwechsel, Energieüberträger und unmittelbar Energiequelle zugeschrieben wurde. Als herausgefunden wurde, welch hohen Energiegehalt das ATP tatsächlich besitzt, wurde es als unmittelbare Energiequelle der Muskelkontraktion erkannt und durch Lohmann, die ihm zugrundeliegenden quantitativen Reaktionen formuliert (vgl. Hollmann/Hettinger 2000, S. 62):
- Kreatinphosphat + ADP ↔ Kreatin + ATP (Kreatinkinase),
- ATP + H2O → ADP + P + Energie (Myosin-ATPase)
ATP
Die Grundlage für jede Muskelkontraktion ist der Abbau von Adenosintriphosphat zu Adenosindiphosphat (ADP) und Phosphat (P) (vgl. Weineck 1994, S. 38). Das ATP ist eine hochenergetische Verbindung, bestehend aus Adenin mit Ribose und drei Phosphaten. Es stellt die einzige Energiequelle dar, welche die Zelle direkt benutzen kann. Das so wichtige ATP ermöglicht nicht nur mechanische Arbeit, sondern auch sehr wichtige energieerfordernde Transformationen, die Aktivierung der freien Fettsäuren und die Erhaltung der labilen Proteinstrukturen (vgl. Hollmann/Hettinger 2000, 62). Die dem ATP entsprechende exergonische Reaktion ist die Myosin-ATPase:
- ATP + H2O → ADP + P + Energie.
In einer separaten chemischen Reaktion kann ATP bis zu AMP (Adenosinmonophosphat) abgebaut werden:
- ATP + H2O → AMP +PP + Energie.
Diese stellt aber eine untergeordnete Rolle in der Energiegewinnung dar. Die Spannungsentwicklung des Muskels ist stark abhängig vom vorliegenden ATP-Gehalt. Erniedrigungen dieses Gehaltes führen (ab einem kritischen Schwellenwert) zunächst zu einer Einschränkung der Spannungsentwicklung und schließlich bis zur Kontraktionsunfähigkeit bei Erregungsreizen. Somit sind die Veränderungen des ATP-Gehalts von der Arbeitsleistung der Muskelzelle abhängig. Sich verkürzende Muskeln, welche jedoch keine Arbeit leisten, zeigen keinen, beziehungsweise nur einen unwesentlichen ATP-Abfall. Arbeit verrichtende Muskeln, welche auch unter Belastung stehen, zeigen einen in Abhängigkeit dieser Arbeitsleistung erniedrigten ATP-Spiegel und einer dementsprechenden Wärmebildung. Die entstehende Wärmeentwicklung bei der Muskelarbeit lässt sich somit mit einer Veränderung des ATP-Spiegels erklären (vgl. Keul/Doll/Keppler 1969, S. 19).
Durch den Abbau des durch die Myosin-ATPase entstandenen ADP kann durch eine geeignete Reaktion, der Myokinase:
- 2 mol ADP → 1 mol ATP + 1 mol AMP,
unter extremen Bedingungen ATP gewonnen werden. Dadurch wird das wichtige ATP unmittelbar für weitere Kontraktionsarbeit bereitgestellt. Folgernd durch die aus der Myosin-ATPase hohen ADP-Konzentrationen und der Sensivität der Myokinase auf hohe ADP-Spiegel, d. h. ihre Aktivität wird durch einen hohen ADP-Spiegel gesteigert, gilt die ADP-Konzentration als Regelgröße für die Bereitstellung von ATP aus ADP. Somit ist das scheinbare Endprodukt ADP keine zu vernachlässigende Größe, denn ihre energiereiche Phosphatbindung kann noch genutzt werden.
Der direkt verfügbare ATP-Speicher reicht unter starken muskulären Beanspruchungen nur für ca. 1 Sekunde um Energie bereit zu stellen. Selbst unter der Voraussetzung, dass ATP bis zu AMP gespalten wird, herrscht im ruhenden Muskel nur ein ATP-Vorrat von ca. 6 µMol/g = 6 mMol/kg (vgl. Keul/ Doll/ Keppler 1969, S. 20). Wenn man nun die Tatsache bedenkt, dass der Mensch täglich soviel ATP verbraucht, wie es seinem Körpergewicht entspricht, ist es also umso erstaunlicher, dass ATP, welches doch als so wichtig für die Muskelkontraktion gilt und die einzige unmittelbare Energiequelle darstellt, nur so beschränkt in der Muskelzelle vorhanden ist.
Kreatinphosphat (PKr)
Nachdem der im Muskel vorhandene ATP-Vorrat nur für eine bis drei Muskelkontraktionen ausreicht, muss der Körper ständig um eine Resynthese des ATPs als lebensnotwendige Substanz bemüht sein. Hier kommt das Kreatinphosphat ins Spiel, welches eine energiereiche chemische Verbindung aus Kreatin (Kr) und einem Phosphatrest ist. Die vorliegende Bindung zwischen dem Phosphat und dem Kreatin hat ein entsprechendes Energiepotential, das des ATP. Durch die schnell ablaufende Reaktion:
- ADP + Kreatinphosphat ↔ ATP + Kreatin,
wird durch die Abspaltung des Phosphatrestes und dessen Übertragung auf ADP das ATP resynthetisiert (vgl. Weineck 2004, S. 101). Zudem ist PKr in etwa drei- bis viermal so großer Menge – 20–30 µMol/g – gegenüber dem ATP in der Muskelzelle vorrätig (vgl. Keul/ Doll/ Keppler 1969, S. 22). Der Kreatinphosphatspeicher ist also von großer Bedeutung für die Leistungsfähigkeit der Skeletmuskulatur, da er bei starker Konzentrationsarbeit bis zu 20 Sekunden (vgl. Weineck 1994, S. 20) in der Lage ist, die dafür notwendige Energie bereit zustellen. Außerdem ist es die Energiequelle, welche das ATP sofort resynthetisieren kann, bis dann zu einem späteren Zeitpunkt andere Reaktionswege aktiviert sind.
Im Kreatinphosphat wird des Weiteren eine entscheidende Rolle als Energiegefälle gesehen, wodurch hohe Substratdurchsätze ermöglicht werden. Es steht auch fest, dass der Kreatinphosphatgehalt von der Höhe und Dauer der geleisteten Arbeit abhängt. Kommt es zu extrem starken Belastungen, kann der Kreatinphosphatspeicher fast vollständig ausgeschöpft werden und nach Ende der Belastung schnell wieder aufgefüllt werden. Sollte es jedoch dazu kommen, dass die Nachlieferung der energiereichen Phosphate unterbleibt, kommt es zu einem Erlöschen der Kontraktionsfähigkeit des Muskels (vgl. Keul/Doll/Keppler 1969, S. 25).
Resynthesewege des ATP
Anaerob alaktazide Energiegewinnung
Jede sportliche Belastung verbraucht Energie. Ist es eine Arbeit höherer Intensität, bei der der Muskel den benötigten Energiebedarf nicht ausreichend oxydativ herbeiführen kann, wird der Muskel gezwungen, die notwendige Energie zur Aufrechterhaltung der Arbeit auf einem anderen Weg zu gewinnen, dem anaerob alaktaziden Energiestoffwechsel. Dieser ist gekennzeichnet durch seine Verarbeitung von ATP und Kreatinphosphat.
Die erste energieliefernde Reaktion ist die Myosin-ATPase:
- ATP + H2O → ADP + P + Energie,
bei der die entstandenen Zerfallsprodukte ADP und das anorganische Phosphat (P) die Atmung um das bis zu 100fache steigern können. Damit sorgen sie für eine hochgradige Aktivierung der für den Muskelstoffwechsel verantwortlichen Funktionssysteme. In einer zweiten chemischen Reaktion kann das ATP nochmal gespalten und bis zu AMP abgebaut werden:
- ATP + H2O → AMP + PP + Energie.
Wenn dann das Arbeitsende eintritt, womit eine Rückbildung des gewonnenen ADP und des Phosphats zu ATP verbunden ist, wird die Atmung schließlich wieder gehemmt. Diese Regulation wird als Atmungskontrolle durch den Energiebedarf bezeichnet (vgl. Senger/ Donath 1977, S. 391). Um jetzt einen Abbruch der Muskelarbeit zu verhindern und eine kontinuierliche Muskelkontraktion aufrecht zu erhalten, wird das ATP mit Hilfe des Kreatinphosphat-Speichers mit extrem hoher Geschwindigkeit wieder aufgefüllt. Die Reaktion hierfür lautet:
- ADP + Kreatinphosphat ↔ ATP + Kreatin.
Da das Kreatinphosphat in der Lage ist, die ATP-Konzentration über einen langen Zeitraum auf einem hohen Niveau zu halten, wird mit dieser Resynthese eine Gesamtarbeitszeit unter Zuhilfenahme der energiereichen Phosphate von maximal 20 Sekunden ermöglicht.
Anaerob laktazide Energiegewinnung (Milchsäuregärung)
Beim anaerob laktaziden Energiestoffwechsel wird durch den Abbau von Traubenzucker (Glukose) oder Glykogen (eine Speicherform der Glukose) über chemische Reaktionen Laktat und ATP hergestellt:
- Glykogen ↔ 2ATP + Milchsäure.
Diese Reaktion wird also als Glykolyse mit anschließender Milchsäuregärung bezeichnet, welche im Sarkoplasma statt findet (vgl. Weineck 2006, S. 101). Als Energielieferant kann Glukose als auch Glykogen herangezogen werden, wobei das intrazelluläre Glykogen energetisch vorteilhafter ist, da es nicht erst über die Blutwege herbeigeführt werden muss. Der Abbau von 1 Mol Glukose zu Laktat bringt 2 Mol ATP. Wird Glykogen verwertet bringt das 3 Mol ATP (vgl. Keul/Doll/Keppler 1969, S. 32). Das Zwischenprodukt Brenztraubensäure (Pyruvat) wird während der aeroben Glykolyse also ohne Zuhilfenahme von Sauerstoff, zu Laktat umgewandelt.
Das bei der Milchsäuregärung entstandene Laktat hat jedoch auf den gesamten Stoffwechsel, sowohl lokal als auch allgemein Auswirkungen, da es als Ermüdungsstoff wirkt. Nach maximalen Belastungen sind im Muskel Laktatwerte von bis zu 25 mMol/kg, im Blut bis zu 20 mMol/kg (vgl. Weineck 1994, S. 39) zu finden. Diese Werte bedeuten eine extreme Übersäuerung im lokalen Gewebe sowie im arteriellen Blut, was mit einer Azidose (stark herabgesetzter pH-Wert) verbunden ist. Durch die Azidose kommt es zu einer Enzymhemmung, welche ein Erliegen der glykolytischen Stoffwechselprozesse herbeiführt. Dieser Abbruch der Maximalbelastung stellt eine wichtige Schutzfunktion für den Organismus dar. Sie verhindert eine zu starke Übersäuerung des Muskels, welche eine Zerstörung intrazellulären Eiweißstrukturen zur Folge hätte (vgl. Weineck 1994, S. 39).
Aerobe Energiegewinnung
Die aerobe Energiegewinnung dient dazu, für eine Belastungsdauer welche über eine Minute hinausgeht, die Energie bereitzustellen (vgl. Keul/ Doll/ Keppler 1969, S. 38). Die dafür notwendigen Reaktionen finden in den Mitochondrien statt. Die aktivierte Essigsäure (Acetyl-CoA), welche durch oxidative Decarboxylierung entstanden ist, durchläuft für den weiteren Abbau den Zitronensäurezyklus und die Atmungskette. Bei dieser Art von Energiebereitstellung werden aus 1 Mol Glukose 36 Mol ATP gewonnen. Wird das intrazelluläre Glykogen zum Abbau verwendet, entstehen sogar 38 Mol ATP:
- 1 Glukose + 6 O2 + 36 ADP + 36 P → 6 CO2 + 6 H2O + 36 ATP
Außerdem können, im Gegensatz zur anaeroben Energiegewinnung, hier neben Glukose auch Fette (in Form von freien Fettsäuren = FFS) verbrannt werden (1 Mol FFS ergibt ca. 130 Mol ATP). Zudem kann in Notfällen noch Eiweiß als Energielieferant dienen, wobei diese beiden Arten der Energiegewinnung (FFS und Eiweiß) nur bei sehr geringen Belastungsintensitäten möglich sind.
Bei der aeroben Energiegewinnung kann also 18 bzw. 19 mal so viel ATP gewonnen werden kann wie bei der anaeroben Glykolyse (vgl. Weineck 2004, S. 104). Diese hohe Energieausbeute hat jedoch auch einen entscheidenden Nachteil. Mit Hilfe der oxydativen Verbrennung werden zwar viele Mol ATP bereitgestellt, doch erfolgt diese Energiebereitstellung über lange Reaktionsketten, weshalb es längere Zeit dauert, bis diese Energie zur Verfügung steht.
Aufgrund der Tatsache, dass die Kontraktionsgeschwindigkeit des Muskels bei den energiereichen Phosphaten am schnellsten und bei der oxydativen Energiebereitstellung am langsamsten ist, bedingt durch verschiedene Flussraten, ist bei verschiedenen Belastungsintensitäten mit verschiedener Belastungsdauer häufig eine Mischform der energieliefernden Systeme zu beobachten. Somit verändert sich die Intensität der Muskelarbeit, also die Kontraktionsgeschwindigkeit der Muskelfaser, in Abhängigkeit von der energetisch möglichen Flussrate.
Es zeigt sich demnach die große Nützlichkeit der unterschiedlichen Flussraten. Sollen z. B. hohe Intensitäten erzielt werden (hohe Energieumsätze), dies ist vor allem bei Schnelligkeitsbelastungen der Fall, müssen höhere Flussraten erzielt werden. Folglich muss auf die anaerob-alaktazide (ATP, PKr) als auch auf die laktazide Energiegewinnung zurückgegriffen werden. Sollen geringere Arbeitsintensitäten abgedeckt werden, wie z. B. bei Langstreckenläufen, überwiegen zwangsläufig die aeroben Energiebereitstellungsprozesse.
Zusammenfassend lässt sich also sagen, dass es bei der Energiebereitstellung eine „Reihenfolge“ gibt. Die primäre Energiequelle ATP wird nacheinander durch das PKr, dann durch die anaerobe Glykolyse und zum Schluss durch die aerobe Energiegewinnung bereitgestellt, wobei sich die einzelnen Speicher jeweils auf Kosten des Nachfolgenden auffüllen.
Sauerstoffschuld
Sauerstoffschuld ist ein Begriff, der im Zusammenhang mit der anaeroben Energiegewinnung steht. Je nach Qualität und Quantität der Arbeit vergehen 2–6 Minuten, bis bei einer submaximalen Belastungsintensität die Sauerstoffaufnahme der Arbeitsintensität entspricht (vgl. Weineck 1994, S. 39). Bleibt die Arbeitsintensität konstant, so stellt sich ein Zustand namens steady state ein. Die Zeit, bis der Zustand des steady state der O2-Aufnahme eingetreten ist, wird als Arbeits-Anlaufzeit bezeichnet. Während der Arbeits-Anlaufzeit wird in Relation zur Arbeitsintensität bzw. zur Höhe des steady state zu wenig Sauerstoff aufgenommen. Diese Menge an Sauerstoff wird als O2-Defizit bezeichnet. Tritt dann das Arbeitsende ein, so kehrt die Sauerstoffaufnahme ähnlich einer e-Kurvenform nach mehreren Minuten zum Ruheausgangswert zurück. Die Menge an Sauerstoff, die nun noch über dem Ruheausgangswert liegt, wird als Sauerstoffschuld bezeichnet.
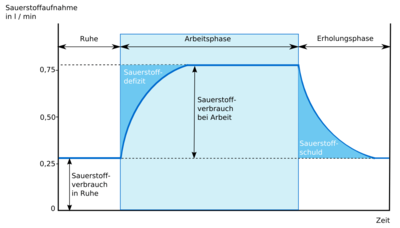
Jedoch muss erwähnt werden, dass der Begriff „Sauerstoffmehraufnahme nach Arbeitsende“ den Begriff „Sauerstoffschuld“ ersetzt, da er sich aus mehreren Faktoren zusammensetzt:
- erhöhte Tätigkeit des Herzmuskels und der Atmungseigenmuskulatur mit einem entsprechen vergrößerten Sauerstoffbedarf,
- Wiederauffüllung von O2-Speichern (Hämoglobin, Myoglobin),
- erhöhter Sauerstoffbedarf des Körpergewebes infolge einer erhöhten Körpertemperatur und eines erhöhten Katecholaminspiegels (eine erhöhte Adrenalinausschüttung verursacht eine Steigerung oxydativer Prozesse).
Sauerstoffschuld ist nur noch dann zu verwenden, wenn es sich um die Rephosphorylierung von Kreatin zu Kreatinphosphat (vgl. Cunningham/ Faulkner 1969, S. 68) und damit zur Wiederauffüllung der energiereichen Phosphate handelt.
Weitere Energieträger
Damit die Muskelzelle nicht nur auf die bisherig angeführten Energiebereitstellungspro-zesse angewiesen ist, kann die Kontraktionsfähigkeit des Muskels noch durch weitere energiereiche Phosphate ermöglicht werden, vorausgesetzt, es ist eine ausreichende Sauerstoffversorgung gegeben. Daraus wird also verständlich, dass der Energiebedarf der Muskelzelle, der fast ausschließlich durch oxydative Stoffwechselprozesse gedeckt wird, auch von anderen energiereichen Substraten gedeckt werden kann (vgl. Keul/Doll/Keppler 1969, S. 151).
Die wichtigsten Energielieferanten für die Muskelzelle sind Kohlehydrate, Fette und Eiweiße, welche ständig durch die Nahrung ergänzt werden müssen. Hierbei decken Kohlehydrate etwa 2/3 des Energiebedarfs und Fette 1/3 (vgl. Weineck 1994, S. 41). Eiweiße können an dieser Stelle vernachlässigt werden, da sie für den Energiestoffwechsel keine wichtige Rolle spielen, sondern nur für den Baustoffwechsel tätig sind (vgl. Wei-neck 2004, S. 109). In Ruhe wird der Energiebedarf also hauptsächlich durch Kohlehydrate und Fette abgedeckt (vgl. Weineck 2004, 108).
Bei lang- und kurzfristigen Belastungen ist der Substratumsatz des Muskels qualitativ als auch quantitativ verschieden. Kurzfristig muskuläre Höchstleistungen unter zwei Minuten werden durch den Abbau von Kohlehydraten, vielmehr durch Muskelglykogen ermöglicht (vgl. Keul/Doll/Keppler 1969, S. 153). Langfristige Arbeit über Stunden wird weniger durch das Muskelglykogen, sondern vielmehr durch das Leberglykogen ermöglicht, wodurch die Glykogenvorräte des Muskels eingespart werden können, da dem Skelettmuskel über den Blutweg die Kohlenhydratvorräte der Leber verfügbar gemacht werden (vgl. Keul/Doll/Keppler 1969, S. 153). Bei Maximalbelastungen unterschiedlicher Dauer wird die Energie also in einem intensitätsspezifischen Mischungsverhältnis bereitgestellt.
Den größten Energiespeicher im Organismus stellen die Fette dar. Jedoch hängt die Bedeutung der Fettverbrennung von unterschiedlichen Faktoren wie Arbeitstyp, Arbeitsdauer, Arbeitsintensität und vom Umfang der eingesetzten Muskelmasse ab (vgl. Weineck 1994, S. 42). In sportlicher Hinsicht, wo die pro Zeiteinheit erreichbare Höchstintensität eine entscheidende Rolle spielt, ist aber auf die Kohlehydrate als entscheidende energieliefernde Substanz hinzuweisen. Entscheidend bei der Verbrennung der verschiedenen Substrate ist nämlich nicht der absolute Wert, hier liefern die Fette 9,3 kcal/g gegenüber nur 4,1 kcal/g bei den Kohlehydraten, sondern der pro Liter Sauerstoff erreichte Brennwert (vgl. Weineck 2004, S. 109). Hierbei ergeben sich pro Gramm:
- Glukose: 5,1 kcal ≈ 6,34 ATP
- Fett: 4,5 kcal ≈ 5,7 ATP
- Eiweiß: 4,7 kcal ≈ 5,94 ATP (vgl. Weineck 2004, S. 109).
Bei gleichem Sauerstoffangebot ermöglicht diese Tatsache gegenüber der Fettverbrennung also einen Mehrgewinn von 13 % bei Glukose und 16 % bei Glykogen (vgl. Keul/Doll/Keppler 1969, S. 153). Aus diesen Werten geht demnach eine Notwendigkeit eines möglichst großen Glykogenspeichers bei Ausdauersportlern hervor. Da die Glykogenvorräte bei sehr langen Dauerbelastungen aber nicht alleine den Energiebedarf abdecken können, wird die Verbrennung von Fettsäuren bei zunehmender Zeitdauer also immer bedeutender. Bei Muskelarbeit über Stunden können die Fettsäuren 70–90 % des Energiebedarfs decken (vgl. Keul/Doll/Keppler 1969, S. 153).
Literaturverzeichnis
- Bartl/ Meinhard/ Moisl: Abitur-Training Biologie, Biologie 1. Fresising, 1987
- Cunningham, D./ Faulkner, J.: The effect of training on aerobic and anaerobic metabol-ism during short exhaustive run. Med. And Sci. in Sports 2 (1969), 65-70
- Heck, H./ Trainerakademie Köln e.V. (Hrsg.): Energiestoffwechsel und medizinische Leistungsdiagnostik. Schorndorf, 1990
- Helmich, U.: Citratzyklus. http://www.u-helmich.de/bio/stw/reihe3/citrat1.htm. (24. Januar 2009)
- Hettinger, T./ Hollmann, W.: Sportmedizin, Grundlagen für Arbeit, Training und Prä-vention. Stuttgart, 2000/ 4. Auflage
- Keul, J./ Doll, E./ Keppler, D.: Muskelstoffwechsel, München, 1969
- Senger, H./ Donath, R.: Zur Regulation der oxydativen Substratverwertung im Muskel bei erhöhtem ATP-Umsatz. Medizin und Sport 12 (1977), 391-400
- Weineck, J.: Sportbiologie. Ballingen, 1994/ 4. Auflage
- Weineck, J./ Weineck, A.: Leistungskurs Sport, Band 1. Forchheim, 2004/ 3. Auflage, 2006
Wikimedia Foundation.