- Kontinuierliche Erregungsleitung
-
Die Ausbreitung einer Erregung in und auf andere Zellen im Organismus wird als Erregungsleitung bezeichnet. Der häufig verwendete Begriff „Reizleitung“ ist falsch, da ein Reiz nicht weitergeleitet werden kann, lediglich die durch ihn verursachte Erregung. Ein Reiz erzeugt eine Erregung in Rezeptoren und diese Erregung wird über Nervenzellen zum Erfolgsorgan geleitet. Innerhalb der Zelle geschieht die Erregungsleitung über verkettete Fließgleichgewichte. Zwischen den Nervenzellen erfolgt die Weiterleitung über Neurite (Axone) entweder durch Ionenflüsse (elektrotonisch, siehe Elektrotonus) oder mittels Aktionspotenzialen.
Inhaltsverzeichnis
Grundlagen
Die Probleme bei der Erregungsleitung bestehen zum einen in der relativ geringen Leitfähigkeit der Ionen und zum anderen in der recht hohen elektrischen Kapazität der Neurite zur Umgebung. Der ohmsche Widerstand kann aufgrund der kaum zu verändernden Ionenkonzentrationen nur durch die Dicke des Neurits beeinflusst werden. Eine Verdopplung des Durchmessers führt zu einer Verbesserung der Leitfähigkeit auf das Vierfache. Allerdings wird auch durch die damit verbundene Oberflächenvergrößerung der Isolationswiderstand der Membran halbiert. Wichtig für die Leitungsgeschwindigkeit ist die Membranzeitkonstante Τ, das pro Längeneinheit berechnete Produkt aus dem Leitungswiderstand R und der Kapazität C. Letztere ist proportional zur Oberfläche der Membran des Neurits und umgekehrt proportional zu der Dicke. Damit gibt es zwei mögliche Methoden, die Leitungsgeschwindigkeit zu erhöhen: durch Verringerung des Leitungswiderstandes oder durch Verringerung der Membrankapazität.
Elektrotonische Erregungsleitung
Die elektrotonische Erregungsleitung geschieht nur über kurze Entfernungen. Da die Membran um das Axon herum ein relativ schlechter Isolator ist, nimmt das Potenzial mit zunehmendem Abstand ab. Ein Beispiel für eine elektrotonische Erregung findet sich in der menschlichen Netzhaut. Hier wird die Erregung als graduierte, reizanaloge Depolarisation elektrotonisch von den Photorezeptoren zu den Ganglienzellen (über die Fortsätze der Horizontal-, Bipolar- und Amakrinen Zellen) übertragen. Diese Form der Erregungsleitung reicht aufgrund des Verhältnisses zwischen Ionenleitung im Inneren (elektrischer Widerstand des Cytoplasmas) und Isolation nach außen (gewisse Leitfähigkeit der das Cytoplasma umgebenden Zellmembran) im besten Falle einige Hundertstel Millimeter weit. Das Potenzial muss dann wieder durch Aktionspotenziale (Spikes) auf ein Niveau angehoben werden, welches eine weitere Fortleitung beziehungsweise Verarbeitung der Information erlaubt.
Erregungsleitung durch Aktionspotenziale
In Axonen von Nervenzellen bewirkt eine Depolarisation die vorübergehende Öffnung spannungsaktivierter Natriumkanäle. Die resultierende Depolarisationswelle läuft als Aktionspotential über die Nervenfaser. Je nachdem, ob das Axon myelinisiert ist oder nicht, unterscheidet man zwei verschiedene Weisen:
Kontinuierliche Erregungsleitung
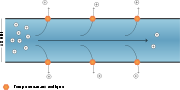 Kontinuierliche Erregungsleitung
Kontinuierliche ErregungsleitungBei marklosen Nervenfasern, das heißt bei fehlender Myelinisierung, wird der Impuls durch das Axon von Abschnitt zu Abschnitt übertragen, indem der vorhergehende Abschnitt ein Aktionspotenzial an den benachbarten, noch nicht erregten Abschnitt weiterleitet. Der vorhergehende Abschnitt ist bereits in der Repolarisationsphase, während der neu erregte Abschnitt schon seine Permeabilität ändert, um selbst ein Aktionspotenzial zu erreichen. Diese Form der Weiterleitung ist relativ langsam (meist nur 1–3 m/s, maximal 30 m/s) und findet sich bei Nerven, welche die inneren Organe versorgen, recht häufig. Bekannt sind auch die recht geringen Leitungsgeschwindigkeiten bei Nozizeptoren, die Durchmesser von unter einem Mikrometer haben. Die Leitungsgeschwindigkeit kann durch eine Verdickung des Axons vergrößert werden. Besonders bekannt sind in diesem Zusammenhang die gut untersuchten so genannten Riesenaxone bei Tintenfischen und Meeresschnecken der Gattung Aplysia mit Durchmessern bis zu einem Millimeter. Die Erhöhung des Durchmessers ist allerdings nicht sehr effektiv, da die Verringerung des Leitungswiderstandes teilweise durch eine Vergrößerung der Membrankapazität und einer Verringerung des Membranwiderstandes wieder aufgehoben wird. Eine Verdopplung des Durchmessers führt nur theoretisch zu einer Verdopplung der Leitungsgeschwindigkeit, liegt praktisch aber noch darunter.
Saltatorische Erregungsleitung
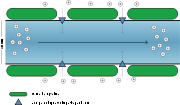 Saltatorische Erregungsleitung
Saltatorische ErregungsleitungBei Wirbeltieren (Vertebraten) sind die meisten Axone von einer Myelinscheide umhüllt (markhaltige Nervenfaser), die von Schwannschen Zellen im peripheren Nervensystem oder von Oligodendrozyten im Zentralnervensystem gebildet wird und die im Abstand von etwa 2 mm an den Knoten oder Ranvierschen Schnürringen unterbrochen ist. Durch diese Isolation vergrößert sich die Kabelkonstante des Axons (ein Maß für die Länge der elektrotonischen Ausbreitung) von wenigen Hundertstelmillimeter auf einige Millimeter. Da die Isolation auch zu einer Verringerung der elektrischen Kapazität führt, verringert sich die Membranzeitkonstante ebenfalls. Allein durch diesen Effekt sind Fortleitungsgeschwindigkeiten von bis zu 180 m/s bei kaum verändertem Querschnitt des Axons möglich. Weil sich die zur Erzeugung des Aktionspotenzials nötigen schaltbaren Kanäle lediglich in den Ranvierschen Schnürringen befinden, wird außerdem weniger Energie verbraucht. Bei der Geschwindigkeit von 120 m/s hat ein Nervenimpuls von 1 ms Dauer eine Länge von 120 mm. Das heißt, beim Durchlauf eines Impulses sind rund 60 Schnürringe gleichzeitig in Erregung. An der Vorderfront des sich ausbreitenden elektrischen Impulses gibt es einen ständigen Wechsel zwischen der elektrotonischen Leitung (zwischen den Schnürringen) und der Regeneration der Amplitude des Aktionspotenzials (in den Schnürringen). Dieser Wechsel wird oft als „Springen“ bezeichnet, obwohl die Fortleitung des Potenzials kontinuierlich ist. Es wird lediglich in den Schnürringen wieder auf den Wert des Aktionspotenzials angehoben.
Nach der Geburt fehlen die Markscheiden beim Menschen noch, bilden sich aber im Laufe der Kindheit heran. Bei demyelinisierenden Erkrankungen wie zum Beispiel Multipler Sklerose werden im Zentralnervensystem die Myelinscheiden abgebaut und führen zu vielfältigen Ausfallerscheinungen.
Erregungsleitung im Neuron
An einer Nervenzelle können bis zu 20 000 Synapsen an den Dendriten und am Zellkörper andocken. Jede einzelne Erregung wird spezifisch zum Axon geleitet. Bei einer der vielen Synapsen reicht eine einzelne Erregung, um ein Aktionspotential am Axonende auszulösen, andere Eingangserregungen hemmen, wieder andere benötigen viele Eingangsimpulse wiederholt oder parallel zur Überschreitung des Schwellenpotentials. Die meisten Eingangserregungen werden über die Zellmembran zum Axon geleitet, seltener durchs Soma. Nach jeder Erregung verändert sich das Verhalten des Neurons. Wiederholte Aktionspotentiale ermüden das Neuron. Nach jeder Aktion werden die speziellen Wege korrigiert (verfestigt, umgebaut, gelöst). Die Impulse hierzu erhält das Neuron über anliegende Astrozyten. Jede Nervenzelle ist ganz speziell optimiert.
Erreicht ein Aktionspotential oder eine graduierte Depolarisation die Synapse, löst dies eine Folge von Reaktionen aus, die dazu führt dass kleine Bläschen, die so genannten synaptischen Vesikel, mit der präsynaptischen Membran verschmelzen (Exocytose), sich öffnen und dadurch Neurotransmitter in den synaptischen Spalt ausschütten. Diese Transmitter öffnen entweder direkt (ionotrop) oder indirekt (metabotrop) ligandengesteuerte Ionenkanäle in der postsynaptischen Membran. Die Ionenspezifität dieser Kanäle entscheidet ob die postsynaptische (Nerven-, Muskel-, Rezeptor- oder Drüsen-)Zelle depolarisiert (erregt) oder hyperpolarisiert (gehemmt) wird.
Am Beispiel der neuromuskulären Synapse (Verbindungsstelle zwischen Nervenzelle und Muskel) wird dazu aus den Vesikeln der Transmitter Acetylcholin ausgeschüttet, der den synaptischen Spalt passiert. Die Transmittermoleküle werden an Rezeptormoleküle der postsynaptischen Membran (also der Membran der Folgezelle) gebunden. Je nach Art des Transmitters wird ein erregendes oder ein hemmendes postsynaptisches Potential an die Folgezelle weitergeleitet.
Im Anschluss spaltet (in diesem Fall) die Acetylcholinesterase den Acetylcholin-Transmitter in Acetat und Cholin. Das Cholin wird durch einen Cholinkanal in der Synapse wieder aufgenommen, mit Essigsäure verbunden und wieder als Acetylcholin in einem Vesikel gebunden.
Erregungsverteilung im Nervengewebe
Die Verteilung einer Erregung auf mehrere Neurone wird Divergenz genannt (die Erregung zweigt sich auf). Erhält dagegen ein Neuron Erregung von mehreren Neuronen, dann vereinigt sich die Erregung (Konvergenz). Die hemmende Wirkung auf andere Neurone wird mit Inhibition (Neuron) beschrieben. Wegen der Ähnlichkeit zu einer Kette (die Erregung wird von Neuron zu Neuron weitergegeben), wird die Erregung von Neuron zu Neuron auch als Erregungskette bezeichnet.
Im Nervengewebe findet eine ständige Umbildung der Erregungsleitungen statt. Neue Verbindungen beginnen mit dem Wachstumskegel. Das Lösen bestehender Verbindungen wird Jäten genannt. Bahnung verfestigt bestehende Leitungen.
Ein Erregungsmuster besteht aus einer Unmenge einzelner Erregungsleitungen. Bei einer realen Reizsituation werden immer mehrere Rezeptoren erregt und diese generieren jeweils Erregungsleitungen. Am Erregungsmuster, und nicht an der einzelnen Erregungsleitung, ist die Reizsituation ersichtlich. Beispiel: Unterscheiden der Früchte Apfel und Birne.
Ein Aktivitätsmuster besteht ebenfalls aus Unmengen einzelner Erregungsleitungen. Im Unterschied zum Erregungsmuster wird die jeweilige Aktivität nicht zwingend durch Rezeptoren verursacht. Am Aktivitätsmuster, und nicht an der Aktivität einzelner Hirnareale, ist das jeweilige Denken ersichtlich. Bei den derzeitigen technischen Möglichkeiten (Hirnscan) ist es nur grob möglich, am Aktivitätsmuster die Gedanken zu erkennen.
Erregungsausbreitung im Herzen
Die Erregungsausbreitung im Herzen stellt durch die Kombination von Erregungsleitungssystem und Erregungsübergabe von Zelle zu Zelle eine Einzigartigkeit im Körper dar.
Weblinks


Wikimedia Foundation.