- Xylem
-
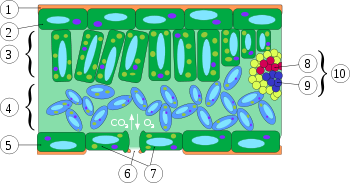 Xylem (8) im Querschnitt eines Blattes; (1) = Cuticula; (2) = obere Epidermis; (3) = Palisadengewebe; (4) = Schwammgewebe; (5) = untere Epidermis; (6) = Spalt der Spaltöffnung; (7) = Schließzellen; (8) = Xylem; (9) = Phloem; (10) = Blattader / Leitbündel
Xylem (8) im Querschnitt eines Blattes; (1) = Cuticula; (2) = obere Epidermis; (3) = Palisadengewebe; (4) = Schwammgewebe; (5) = untere Epidermis; (6) = Spalt der Spaltöffnung; (7) = Schließzellen; (8) = Xylem; (9) = Phloem; (10) = Blattader / Leitbündel
Das Xylem [ksy’le:m] (griech.: ξυλον, Holz) oder der Holzteil der höheren Pflanzen ist ein komplexes, holziges Leitgewebe, das dem Transport von Wasser und anorganischen Salzen durch die Pflanze dient, aber auch Stützfunktionen übernimmt.
Das Xylem findet sich zusammen mit dem Phloem in Leitungsbahnen, den so genannten Leitbündeln, die die Sprossachsen (bei krautigen Stängel, bei Bäumen Stamm genannt), die Blattstiele und Blätter durchziehen. Wurzeln besitzen einen zentralen Xylemkern.
Inhaltsverzeichnis
Struktur
Xylem kann gefunden werden:
- in Leitbündeln, wiederum zu finden in nicht verholzten Pflanzen und den nicht verholzten Pflanzenteilen von verholzten Pflanzen
- als sekundäres Xylem, gebildet in verholzten Pflanzen von einem Kambium zwischen primären Xylem und Phloem
- als ein Teil von Stelen, die nicht in Leitbündeln angeordnet sind, in vielen Farnen
In Übergangsphasen von Pflanzen mit sekundären Wachstum schließen sich die ersten beiden nicht gegenseitig aus, obwohl ein Leitbündel normalerweise nur primäres Xylem enthalten wird.
Die Verzweigungen des Xylems folgen Murrays Gesetz.[1]
Holz (also Xylem im sekundären Zustand) dient- als Festigungssystem
- als Wasserleitungssystem (Hydrosystem)
- und als Speichersystem für Assimilate.
Die verschiedenen Zelltypen lassen sich diesen Funktionen zuordnen. Tracheiden weisen stark verholzte Wande auf, sind abgestorben und dienen sowohl der Festigung, wie auch der Wasserleitung (mit maximal 0,4 mm/s). Tracheenglieder dienen ausschließlich der Wasserleitung und sie sind ebenfalls abgestorben. Tracheenglieder besitzen aufgelöste Zwischenwände und bilden lange Röhrensysteme, die Tracheen. Sie besitzen einen wesentlich größeren Durchmesser, was in einem deutlich geringeren Widerstand und damit schnelleren Wassertransport resultiert (bis zu 15 mm/s, in Extremfällen 40 mm/s). Holzfasern ähneln den Tracheiden, besitzen jedoch wesentlich stärker verdickte Wände und keine Tüpfel. Holzfasern sind oft tot, in dem Fall dienen sie der Festigung, aber nicht immer, in diesem Fall dienen sie auch der Speicherung. Sowohl Tracheiden, wie auch Tracheen und Holzfasern sind langgestreckt, also prosenchymatisch. Holzparenchymzellen sind lebendig, sie dienen der Speicherung von Stärke und Öl und spielen eine wichtige Rolle bei der Reparatur von Embolien. Sie sind nicht langgestreckt (sondern isodiametrisch). Das Holz der Gymnospermen besteht in erster Linie aus Tracheiden und ist monoton aufgebaut. Tracheen sind nicht vorhanden und Parenchym nur um Holzstrahlen und Harzkanäle. Das Holz der Angiospermen ist komplizierter gebaut. Hier sind Tracheen auf die Wasserleitung spezialisiert und Holzfasern auf Festigung. Holzstrahlen sind umfangreicher und aus mehreren Zellschichten aufgebaut.[2]
Das Xylem von Pflanzen, die entwicklungsgeschichtlich sehr alt sind, etwa von Farnen und Koniferen, besteht ausschließlich aus Tracheiden. Bei den meisten Bedecktsamern (Angiospermen) enthält das Xylem auch gut entwickelte Gefäße und Holzfasern. Da die Abfolge der Schritte in der Spezialisierung all dieser Gewebe gut zu beobachten ist, liefert die Erforschung des Xylems wichtige Hinweise auf die Entwicklungsgeschichte der höheren Pflanzen.
Als Hadrom bezeichnet man ein Xylem ohne Festigungszellen, also ohne Sklerenchymfasern. Das Hadrom ist dem Xylem sehr ähnlich, kommt allerdings nicht wie das Xylem in Kormophyten (Gefäßpflanzen), sondern in Bryophyten (Moosen) vor. Die Gesamtheit des Wasserleitgewebes der Bryophyten wird als Hadrom bezeichnet.
Zellarten
Tracheiden
→ Hauptartikel Tracheide
Tracheiden sind lang gestreckte in der Jugend lebende, später abgestorbene Zellen mit dicken verholzten Zellwänden, für die kleine, fest umrissene dünne Bereiche kennzeichnend sind, die so genannten Tüpfel. Tracheiden sind Einzelzellen, deren Querwände nicht vollständig aufgelöst, sondern "getüpfelt" sind (Hoftüpfel).
Die Funktion der Hoftüpfel mit Porus und Torus (Gymnospermen) liegt in der Wasserleitung sowie in einer Ventilfunktion, die einen Lufteintritt verhindert (Luftembolie!). Tracheiden kommen meist in Gymnospermen, aber auch bei Angiospermen vor. Gymnospermen (z. B. Nadelbäume) haben nur Tracheiden, keine Tracheen.
Tracheen
→ Hauptartikel Trachee (Pflanze)
Die Einzelzellen der Tracheen, die Gefäßglieder, sind spezialisierte Tracheiden, deren Zellwände an ihren Enden eine oder mehrere Poren aufweisen; vertikal aneinandergereihte abgestorbene Gefäßglieder bilden eine durchgehende Röhre, die als Trachee oder Gefäß bezeichnet wird. Tracheen kommen vor allem in Angiospermen vor und dienen zur Leitung von Wasser und den darin gelösten Salzen (Elektrolyten).
Sklerenchymfasern
→ Hauptartikel Sklerenchym
Sklerenchymfasern sind spezialisierte Tracheiden mit stark verdickten, verholzten Zellwänden. Der Leitung dienen sie nur in geringem Umfang, vielmehr ist ihre Aufgabe, das Xylem mechanisch zu verstärken. Fertig ausgebildet sind sie abgestorben.
Holz/Xylemparenchym
Sind wenig spezialisierte, lebende Zellen, die im Holzteil vorkommen. Sie sind normalerweise nicht längsgestreckt, also isodiametrisch (d. h. die Zelldurchmesser sind in allen Richtungen annähernd gleich).
Primäres und sekundäres Xylem
Primäres Xylem wird während des primären Wachstums in den Vegetationskegeln der Sprossachse und der Wurzeln vom Prokambium gebildet. Es umfasst Protoxylem und Metaxylem. Metaxylem entwickelt sich nach dem Protoxylem aber vor dem sekundären Xylem. Xylem entwickelt sich nach bestimmten Mustern, die in der jeweiligen Position von Proto- und Metaxylem variieren, z.B. endarch, in welchem das Protoxylem zum Zentrum des Stammes oder der Wurzel gerichtet ist und exarch, in welchem das Metaxylem zum Zentrum hin gerichtet ist.
Sekundäres Xylem wird durch Zellteilung des Kambiums, welches sich zwischen Xylem und Phloem befindet, gebildet. Das Kambium gibt nach innen Zellen des sekundären Xylems ab, nach außen Zellen des sekundären Phloems. Ein solches Kambium, welches nach zwei Seiten Gewebe bildet, nennt man dipleurisches Kambium.
Sekundäres Xylem wird in den Gnetophyta und Ginkgophyta und in geringerem Umfang auch bei Cycadophyta gefunden, jedoch sind die beiden wichtigsten Gruppen:
- Nadelbäume (Koniferen): Es gibt einige 600 Arten von Nadelbäumen. Alle Arten besitzen sekundäres Xylem, das in dieser Gruppe in seiner Struktur relativ gleichmäßig ist. Viele Koniferen werden große Bäume: der sekundäre Xylem solcher Bäume wird als Weichholz verkauft.
- Bedecktsamer (Angiospermen): Es gibt zwischen 400.000 bis einigen viertel Million Arten von Angiospermen. Sekundäres Xylem kann bei Dikotylen, nicht aber bei Monokotylen gefunden werden. Bei nicht-monokotyle Angiospermen kann sekundäres Xylem vorhanden sein, muss aber nicht. Es kann auch innerhalb einer Art durch die individuellen Umgebung der Pflanze variieren. Angesichts der Größe dieser Gruppe ist es nicht überraschend, dass es innerhalb der Angiospermen keine absoluten Regeln für die Struktur des sekundären Xylem gibt. Viele nicht-monokotyle Angiospermen werden Bäume und das sekundäre Xylem von ihnen wird als Hartholz verkauft.
Hauptfunktion – Wassertransport in obere Bereiche
Das Xylem transportiert Wasser und lösliche, mineralische Nährstoffe von den Wurzeln durch und in die Pflanze. Es wird auch verwendet, um das durch Transpiration und Photosynthese verlorene Wasser zu ersetzen. Xylemsaft besteht hauptsächlich aus Wasser und anorganische Ionen, obwohl es auch eine Reihe von organischen Molekülen enthalten kann. Die für den Transport notwendige Energie wird nicht durch die Tracheen selbst aufgebracht, die aus abgestorbenen Zellen bestehen und keine lebenden Bestandteile mehr besitzt. Statt dessen verursachen zwei Phänomene den Wassertransport im Xylem:
- Transpirationssog: Die wichtigste Ursache für ihn ist die Verdunstung von Wasser von den Oberflächen der Mesophyllzellen. Diese Transpiration verursacht Millionen von winzigen Menisken in der Zellwand des Mesophyll. Die daraus resultierende Oberflächenspannung bewirkt einen negative Druck oder bzw. einen Zug im das Xylem, der das Wasser aus der Wurzel bis ins Mesophyll zieht.
- Wurzeldruck: Wenn das Wasserpotenzial der Wurzelzellen negativer als das der Erde ist, in der Regel durch eine hohe Konzentrationen von gelösten Stoffen, wird Wasser durch Osmose aus dem Boden in die Wurzel bewegt. Dies führt zu einem positiven Druck, der den Xylemsaft aus der Wurzel in den Spross zu den Blättern hin zwingt. In einigen Fällen wird auf diese Weise der Saft aus den Blättern durch eine hydathode ausgeschieden, ein Phänomen, das als Guttation bezeichnet wird. Der Wurzeldruck ist am Morgen am höchsten, bevor die Stomata öffnen und so Transpiration möglich wird. Verschiedene Pflanzenarten können einen unterschiedlich starken Wurzeldruck auch in ähnlicher Umwelt aufweisen; Beispiele hierfür sind bis zu 145 kPa in Vitis riparia, hingegen um Null in Celastrus orbiculatus.[3]
Die primäre Kraft, die die Bewegung des Wassers nach oben in den Pflanzen erzeugt, ist die Kapillarwirkung, beruhend auf der Adhäsion zwischen dem Wasser und der Oberfläche der Leitbahnen des Xylems.[4] [5] Die Kapillarwirkung erzeugt eine Kraft, die ein Gleichgewicht herstellt, das die Schwerkraft ausgleicht. Wenn Wasser durch Transpiration an der Spitze entfernt wird, wird die Strömung aufwärts im Xylem erzeugt, um das Gleichgewicht wieder herzustellen.
Der Transpirationssog ergibt sich aus der Verdunstung von Wasser von den Oberflächen der Zellen in den Blättern. Diese Verdampfung bewirkt, dass das Wasser wieder in die Poren der Zellwand strömt. Durch Kapillarwirkung bildet das Wasser konkave Menisken in den Poren. Die hohe Oberflächenspannung des Wassers zieht die Konkavität nach außen, die resultierende Kraft ist groß genug, um Wasser die z.T. erforderlichen hundert Meter vom Boden bis in die höchsten Zweige zu heben.
Der Transpirationssog erfordert, dass die Gefäße, die das Wasser leiten, sehr klein sind im Durchmesser, da sonst Kavitation die Wassersäule zerstören würde. Da Wasser aus den Blättern verdunstet, wird mehr in die Höhe gezogen, um es zu ersetzen. Wenn der Unterdruck im Xylem ein extremes Niveau aufgrund eines geringen Wassereinstroms in die Wurzel erreicht (wenn z.B. der Boden trocken ist), dann kommen die im Wasser in Lösung vorliegenden Gase zum Vorschein und bilden Blasen - eine Embolie bildet sich. Diese verbreitet sich schnell, hin zu benachbarten Zellen, es sei denn, Hoftüpfel sind vorhanden (diese besitzen einen Torus, der die die Öffnung zwischen benachbarten Zellen in solch einem Falle verschließt).
Kohäsionstheorie
Die Kohäsionstheorie ist eine Theorie der intermolekularen Anziehung, die das aufwärts steigenden Wasser im Xylem (gegen die Schwerkraft) beschreibt, belegt von John Joly und Henry Horatio Dixon. Trotz zahlreicher Einwände ist sie die am ehesten akzeptierte Theorie für den Transport von Wasser durch das Gefäßsystems von Pflanzen, gestützt auf die klassische Forschung von Dixon-Joly (1894)[6] Askenasy (1895)[7] und Dixon (1914,1924).[8][9]
Wasser ist ein polares Molekül. Wenn Wassermoleküle miteinander in Wechselwirkung treten, bilden sich Wasserstoffbrückenbindungen. Das negativ polarisierte Sauerstoffatom eines Wassermoleküls bildet eine Wasserstoffbrücke mit einem positiv polarisierten Wasserstoff-Atom eines anderen Wassermoleküls. Diese attraktive Wechselwirkung ist (zusammen mit anderen intermolekularen Kräfte) einer der wichtigsten Faktoren für das Auftreten der Oberflächenspannung in flüssigem Wasser. Es ermöglicht Pflanzen Wasser aus der Wurzel durch das Xylem in das Blatt zu bewegen.
Wasser wird ständig durch die Transpiration im Blatt verloren. Wenn ein Wassermolekül verloren wird, wird durch Kohäsion und Adhäsion neues Wasser herangezogen. Transpirationssog, unter Verwendung der Kapillarwirkung und die inhärente Oberflächenspannung des Wassers, ist der primäre Mechanismus der Bewegung des Wassers in Pflanzen. Es ist jedoch nicht der einzige beteiligte Mechanismus. Jeglicher Verbrauch von Wasser in den Blättern produziert Kräfte, die Wasser nachsaugen.
Transpiration erzeugt Spannung (also negativen Druck) in den Mesophyllzellen. Aufgrund dieser Spannung wird das Wasser buchstäblich aus den Wurzeln bis in die Blätter gezogen, unterstützt durch Kohäsion (der Zug zwischen den einzelnen Wassermolekülen durch Wasserstoffbrückenbindungen) und Adhäsion (die Klebrigkeit zwischen Wassermolekülen und den hydrophilen Zellwänden von Pflanzen). Dieser Mechanismus der Wasserdurchfluss funktioniert durch den Wasserpotentialgradienten (Wasser fließt von Orten mit hohem Wasserpotential zu Orten mit niedrigem Wassserpotential) und die Regeln der einfachen Diffusion.[10]
Druckmessung
Bis vor kurzem konnte der negative Druck (Sog) des Transpirationssogs nur indirekt gemessen werden, durch Anlegen eines externen Druckes durch eine Scholander Bombe, um den internen Druck auszugleichen.[11] Als Techniken ausgereift genug waren, um direkte Messungen vorzunehmen, gab es Diskussionen darüber, ob die klassische Theorie korrekt war, da es zum Teil nicht möglich war, negative Drücke nachzuweisen. Neuere Messungen scheinen die klassische Theorie größtenteils zu bestätigen. Der Xylem-Transport wird durch eine Mischung aus Transpirationssog und Wurzeldruck erzeugt, was eine Interpretation von Messungen erschwert.
Literatur
- Rudolf Schubert und Günther Wagner: „Botanisches Wörterbuch. Pflanzennamen und botanische Fachwörter“. Stuttgart 2000, S. 595.
Einzelnachweise
- ↑ Katherine A. McCulloh, John S. Sperry and Frederick R. Adler: Water transport in plants obeys Murray's law. In: Nature. 421, Nr. 6926, 2003, S. 939–942. doi:10.1038/nature01444. PMID 12607000.
- ↑ A. Bresinsky, Ch. Körner, J. W. Kadereit, G. Neuhaus, U. Sonnewald: Strasburger – Lehrbuch der Botanik. 36. Auflage, Spektrum Akademischer Verlag, Heidelberg 2008. ISBN 978-3-8274-1455-7, S. 187ff.
- ↑ Tim J. Tibbetts, Frank W. Ewers: Root pressure and specific conductivity in temperate lianas: exotic Celastrus orbiculatus (Celastraceae) vs. Native Vitis riparia (Vitaceae). In: Botanical Society of America (Hrsg.): American Journal of Botany. 87, Nr. 9, 2000, S. 1272–78. doi:10.2307/2656720. PMID 10991898.
- ↑ Pierre Cruiziat and Hanno Richter in web site for Plant Physiology at Sinauer Associates.
- ↑ Editors: Anthony Yeo, Tim Flowers: Plant solute transport. Oxford UK: Blackwell Publishing 2007., ISBN 978-1-4051-3995-3 p 221 Google Books
- ↑ H Dixon, Joly: On the ascent of sap. In: Ann. Bot.. 8, 1894, S. 468–470.
- ↑ E Askenasy: Ueber das Saftsteigen. In: Bot. Cent.. 62, 1895, S. 237–238.
- ↑ H Dixon: Transpiration and the ascent of sap in plants, S. 216, New York: Macmillian 1914
- ↑ H Dixon: The transpiration stream, S. 80, London: University of London Press, Ltd 1924
- ↑ Neil Campbell: Biology, S. 759, San Francisco, CA: Pearson Education, Inc. 2002, ISBN 0-8053-6624-5
- ↑ http://bugs.bio.usyd.edu.au/learning/resources/plant_form_function/external_sites/PAP/Lab08_WaterPot/08Lab_14.html
Siehe auch
Weblinks

Wikimedia Foundation.