- Rhinophore
-

Dieser Artikel wurde aufgrund von formalen und/oder inhaltlichen Mängeln in der Qualitätssicherung Biologie zur Verbesserung eingetragen. Dies geschieht, um die Qualität der Biologie-Artikel auf ein akzeptables Niveau zu bringen. Bitte hilf mit, diesen Artikel zu verbessern! Artikel, die nicht signifikant verbessert werden, können gegebenenfalls gelöscht werden.
Lies dazu auch die näheren Informationen in den Mindestanforderungen an Biologie-Artikel.
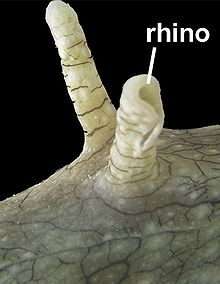
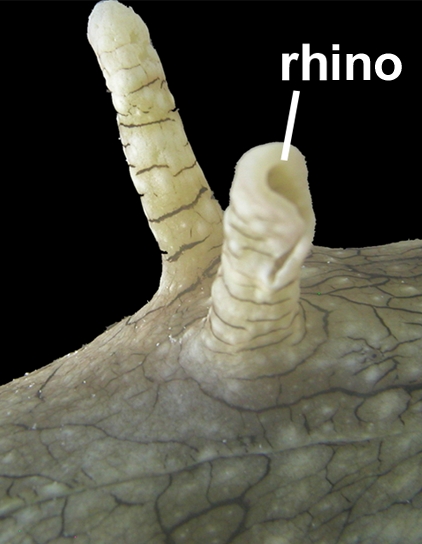
 Chromodoris coi: 2 Rhinophoren auf der Kopfoberseite
Chromodoris coi: 2 Rhinophoren auf der Kopfoberseite

Rhinophoren (altgr. ῥίς (rhis)=Nase, φορος (phoros)=tragend) bilden ein olfaktorisches Sinnesorgan, dass eine artabhängige Auswahl chemischer Substanzen wie z.B. Duftstoffe, Geschmacksstoffe, Pheromone identifiziert. Sie treten u.a. bei Nacktschnecken und bei Meeresschnecken (Opisthobranchia) der Ordnung der Nacktkiemer (Nudisbranchia) auf, insbesondere bei den doriden Nacktkiemern. Das Sinnesorgan besteht aus Chemorezeptoren, biologischen Sensoren, mit denen Substanzen „erkannt“ werden, aus Eingangsneuronen zum Nervensystem, die durch die Rezeptoren induzieren Signale für wahrgenommene Substanzen weiterleiten, und einem Paar antennenartiger Träger, welches sich in exponierter Position auf dem Kopfoberseite befindet und Rezeptoren wie Eingangsneuronen beherbergt.
Inhaltsverzeichnis
Fallstudie am Aplysia californica
Der prinzipielle Aufbau von Rhinophoren wird hier am Beispiel der Art Aplysia californica der vieluntersuchten Gattung der Seehasen beschrieben.[1]
Funktion
Bei den Alypsia dienen die Rhinophoren der Fernwahrnehmung von Substanzen wie Duft- und Geschmacksstoffen sowie der Rheorezeption (Wahrnehmung des Wasserflusses).[1]
Eine große Anzahl von Chemorezeptoren auf der Oberfläche der Rhinophoren ermöglicht ein feines Wahrnehmungsvermögen[2] mit dem Seehasen Nahrungsquellen wie Schwämmen finden und in ihrer Nähe bleiben.
Ferner konnte nachgewiesen werden, dass sie mittels der Rhinophoren Pheromone (Lock und Botenstoffe) entdecken.[1]
Aufbau
Bei den Alypsia sind die Rhinophoren ein Paar Stabantennen, die auf der Oberfläche der Kopfoberseite aufsetzen. Im Vergleich zur Größe des Tieres, sind die Rhinophoren meist recht klein: bei einem ausgewachsenen Aplysia californica betrug ihre Länge nur etwa 1 cm.[1]
Das physische Wahrnehmungszentrum der Rhinophoren ist in einer Furche an deren Spitze enthalten. Die Wahrnehmung erfolgt über spezialisierte Haarzellen (Ziliarzellen) oder Protuberanzen (kleine Ausstülpungen) an der Oberfläche des Außengewebes. Diese Haarzellen sind Chemorezeptoren für Duft- und Geschmackstoffe.
Der Eingang in das Nervensystem erfolgt über Neuronen, die im Gewebe (Epithel) im Furchenraum eingelagert sind. Die Dendriten dieser Neuronen sind mit den Haarzellen und Protuberanzen verbunden. Bei Wahrnehmung von Substanzen durch die Rezeptorzellen an der Oberfläche werden die Neuronen aktiviert. Die Neuronen leiten das Eingangssignal dann über ihre Axone zum Rhinophorenganglion (Ansammlung von Nervenzellen und Verarbeitungszentrum im Nervensystem der Schnecke) weiter (Projektion).[1]
 Rhinophorenspitze eines Aplysia californica
Rhinophorenspitze eines Aplysia californica
Elektronenmikroskop: niedrige Auflösung
Skala: Balken=300 μm.
rg - Rhinophorenfurche
tip - Rhinophorenspitze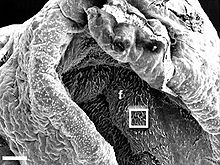 Rhinophorenfurchen mit Ziliarzellen an der Epitheloberfläche
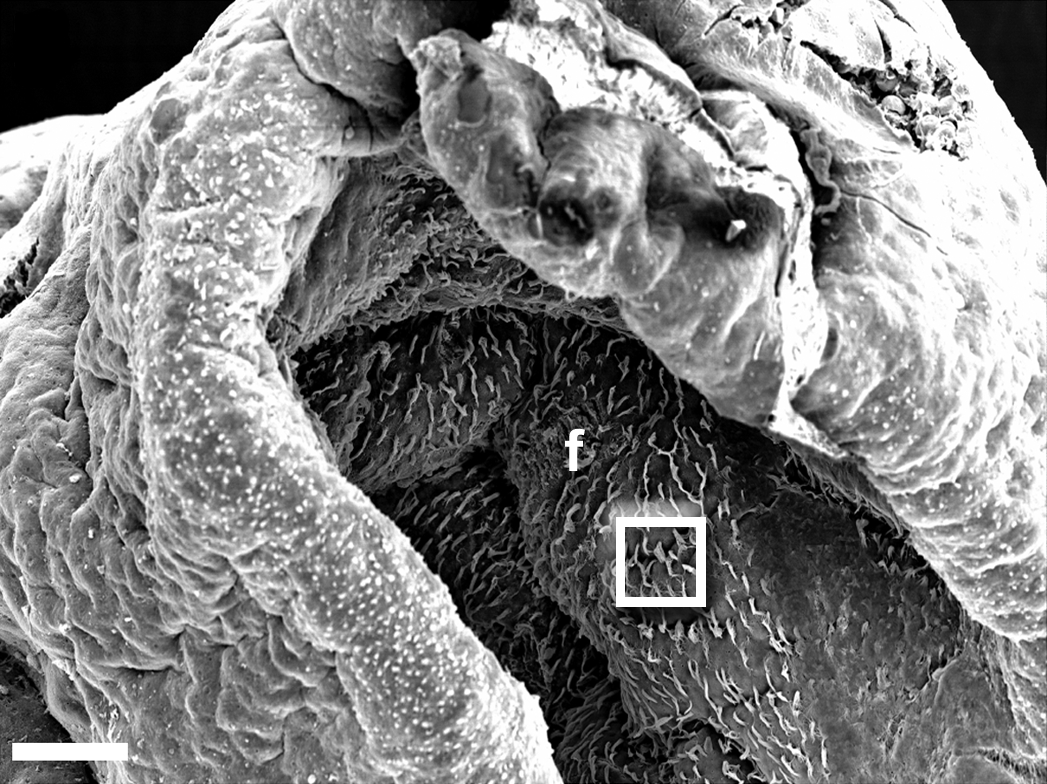
Rhinophorenfurchen mit Ziliarzellen an der Epitheloberfläche
Elektronenmikroskop: mittlere Auflösung
Skala: Balken=100 μm.
f - Faltungen
Quadrat - nächster Ausschnitt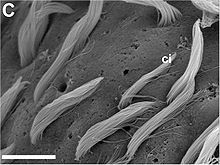 Poren mit und ohne austretende Zilienbündel.
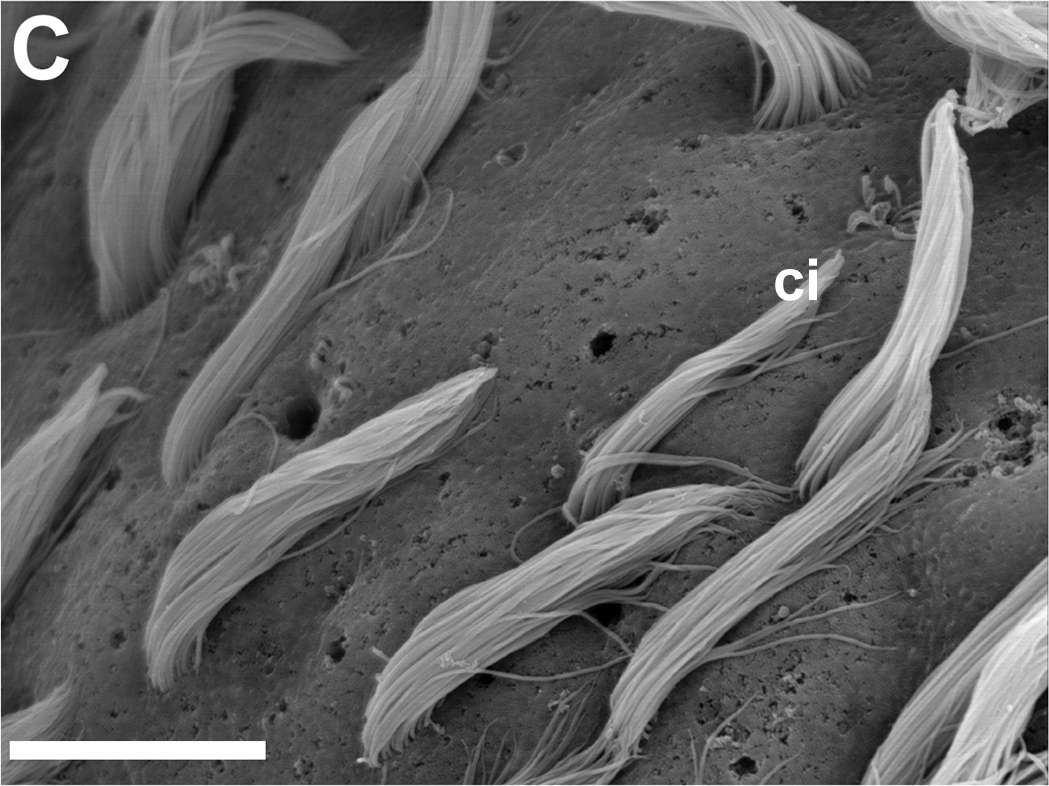
Poren mit und ohne austretende Zilienbündel.
Elektronenmikroskop: hohe Auflösung.
Skala: Balken=10 μm.
ci - Zilien verschiedener LängeIn der Schlundnähe haben die Aplysia californica Tentakel, die ebenfalls mit Rezeptorzellen besetzt sind und der Sinneswahrnehmung dienen. Man geht jedoch von keiner Funktionsüberlappung aus, sondern glaubt, die Tentakelrezeptoren seien eher Chemo- oder Mechanorezeptoren zur Kontaktwahrnehmung.[1]
Erweiterungen
Anhand des Alypsia californica sind die Hauptelemente der olfaktorischen Wahrnehmung per Rhinophoren dargestellt worden: Antenne - Chemoprezeptoren - Eingangsneuronen - Nervensystem - Ganglien. Diese Elemente und die Signalstrecke bleiben bei allen Schneckenarten mit Rhinophoren gleich. Jedoch gibt es Variabilität in den Ausprägungen, auf die nun aufmerksam gemacht wird.
Beweglichkeit von Rhinophoren
Das obige Bild der Chromodoris coi deutet an, dass Rhinophoren nicht nur starre Aufsätze sind. In der Tat können einige Schnecken ihre Rhinophoren in verschiedene Richtungen ausrichten[3].
Die Antennenform der Rhinophoren ist zwar gut für die zu erfüllenden Aufgaben, macht sie jedoch gleichzeitig zu sensiblen Angriffspunkten für ihre natürlichen Feinde: die Rhinophoren sind leicht abzureißen, an- bzw. abzufressen. Um das zu vermeiden, sind die meisten doriden Nacktkiemer in der Lage, ihre Rhinophoren in Taschen unterhalb der Hautoberfläche einzuziehen.[4]
Olfaktorische Wahrnehmung
Gegenüber dem Seh- und Hörsinn weist die olfaktorische Wahrnehmung einige Unterschiede auf.
Seh- und Hörsinn basieren auf Wellen (Licht-, Schallwellen). Wellen breiten sich relativ schnell und gleichmäßig aus. Die Signalübertragung folgt dem Dominosteinprinzip. Stößt man eine Kette von Dominosteinen an, bleiben alle Dominosteine an Ort und Stelle, ändern aber ihren Zustand von "stehend" in "gefallen", und der letzte fallende Stein teilt dem Empfänger mit, dass der Sender die Kette zuvor angestoßen hat. Physisch übertragen werden also physikalische Zustände, nicht aber Bauteile. Ferner genügen verhältnismäßig wenige Typen von Rezeptorzellen für die Wahrnehmung. Im menschlichen Auge sind z.B. Stäbchenzellen für das nächtliche Hell-Dunkel-Sehen und drei Typen von Zapfenzellen für das Herausfiltern von Frequenzbändern (Anteil an Rot-, Grün-, Blaulicht) beim Tagsehen verantwortlich.
Bei der olfaktorischen Wahrnehmung müssen fluktuierende Moleküle chemischer Substanzen aus dem Trägermedium (Wasser) herausgefiltert und zugeordnet werden. Das Herausfiltern geschieht oft durch das Andocken und Eingehen chemischer Bindungen, die dann eine interne Signalkette auslösen. Für das "Erkennen" vieler Substanzen wird daher auch ein hinreichend breites Spektrum an Rezeptorzellen benötigt (ein Mensch hat ~320 Chemorezeptoren, ein Schäferhund ~1200). Die Signalübertragung erfolgt über den physischen Transfer von Molekülen vom Ausgangs- zum Wahrnehmungsort. Moleküle sind aber (z.B. im Vergleich zu Elektronen) recht groß und anfällig für Barrieren. Zum anderen unterliegen sie auf der Signalstrecke Einflüssen des Transportmediums (z.B. Strömungen). Chemische Signale sind daher, wenn sie nicht sehr stark sind, diffus und nicht so gerichtet wie Licht- und Schallwellen.
Daraus kann man allgemeine Gütekriterien für olfaktorische Sinnesorgane aufstellen: 1) pro Typ an Chemorezeptor sollten möglichst viele Rezeptoren vorhanden sein, 2) die Rezeptoren sollten möglichst nicht punktuell konzentriert sondern sinnvoll räumlich verteilt werden, 3) je mehr Typen an Chemorezeptoren, desto besser die Wahrnehmung. Im Raum gut verteilte Rezeptorflächen mit guter Durchsetzung an verschiedenen Typen sind daher wünschenswert.
Form von Rhinophoren
In dem Sea Slug Forum[5] werden diverse Formen von Rhinophoren gezeigt. Diese differieren sowohl in der Form der sensorischen Felder, in der Position und in der Antennenform. Unter obigen Gesichtspunkten kann man die Formen der hervorgebrachten Rhinophoren bewerten.
Viele Rhinophoren haben als Grundform zwei Stabantennen mit aufgesetzten sensorischen Feldern. Stabantennen sind einfache, flexible Konstruktionen. Sie können in diverse Richtungen geneigt, gedreht und zu Schutzzwecken ein-/ausgefahren werden. Andererseits sind die Antennen selbst nicht sehr raumgreifend.
Obige Bildern der Canthodoris pilosa, Chromodoris annulata und Chromodoris coi zeigen Aufsätze mit sensorischen Feldern unterschiedlicher Form, Länge und Riefung. Bei der Chromodoris coi ist fast der gesamte Stab mit spiralig Riefen belegt, bei der Canthodoris pilosa nur der oberer Teil. Des Weiteren sind u.a. noch Felder in Bürstenform, Federform, mit Längsfalten dokumentiert sowie sich am Stabende auffächernde Felder.[5] Bürsten, Federn, Fächer sind geometrische Formen mit großen Oberflächen auf geringem Raum.
Neben Schnecken mit reinen Stabantennen gibt es Schnecken (Sternenschnecke Bornella stellifer), bei denen die Antennen keine Kopfaufsätze sind, sondern sich verzweigenden Tentakeln entstammen.
Etymologie
Der Name Rhinophoren leitet sich von seiner Funktion als Riechorgan ab. "Rhino-" kommt vom alt-griechischen Wort ῥίς (rhis) für Nase. "Phore" stammt ab vom (neulateinischen) "phore" für tragen, und vom griechischen Wort φορος (phoros), was tragend bedeutet.
Quellen
Dieser Artikel enthält Inhalte[1], die der Lizenz CC-BY-2.0, jedoch nicht der Lizenz GFDL unterliegen.
- ↑ a b c d e f g Cummins, F.S; Erpenbeck, D.; Zou, Z.; Claudianos, Ch.; Moroz, L.L.; Nagle, G.T. & Degnan, B.M. 2009. Candidate chemoreceptor subfamilies differentially expressed in the chemosensory organs of the mollusc Aplysia. BMC Biology 2009, 7:28. doi:10.1186/1741-7007-7-28.
- ↑ Rhinophore in nudibranchs. Sea Slug Forum, Zugang 8. Juli 2009.
- ↑ Sea Slug Forum: Drehen der Rhinophoren
- ↑ Sea Slug Forum: Ein-/Ausfahren von Rhinophoren, Philip Cromwell, 28. Juni 2007
- ↑ a b Sea Slug: Formen von Rhinophoren
Literatur
- Wertz, A.; Rössler, W.; Obermayer, M. & Bickmeyer, U. (6. April 2006): "Functional neuroanatomy of the rhinophore of Aplysia punctata". Frontiers in Zoology 3: 6. doi:10.1186/1742-9994-3-6



Wikimedia Foundation.