- Uniformitätsgesetz
-
 Gregor Mendel (1865)
Gregor Mendel (1865)Die mendelschen Regeln wurden benannt nach ihrem Entdecker Gregor Mendel und beschreiben, wie die Vererbung von Merkmalen abläuft, deren Ausprägung von (nur) einem Gen bestimmt wird. Klassische, bereits von Mendel untersuchte Beispiele für solche Merkmale sind die Form und die Farbe von Erbsensamen und die Farbe von Erbsenblüten. Auch die Blutgruppen des Menschen gehören dazu. Die Regeln gelten nur für diploide Organismen (also solche, die von beiden Eltern je einen Chromosomensatz erben) mit haploiden Keimzellen. Dazu zählen die Menschen, die meisten höheren Tiere und viele Pflanzen. Es lassen sich ebenfalls Regeln für Organismen mit höherem Ploidiegrad ableiten.
Die frühere Bezeichnung mendelsche Gesetze ist ungebräuchlich geworden, da mehrere genetische Phänomene entdeckt wurden – beispielsweise Translokationen und die so genannten Springenden Gene − aufgrund derer ein Erbgang von den „Regeln“ abweichen kann.
Inhaltsverzeichnis
Entdeckung
Die mendelschen Regeln wurden in den 1860er Jahren von dem naturwissenschaftlich interessierten Augustinermönch Gregor Mendel durch Kreuzungsversuche an Erbsenpflanzen ermittelt und in einer zunächst wenig beachteten Publikation[1] formuliert. Mendel führte dafür die Begriffe „rezessiv“ und „dominierend“ ein (statt des letzteren wird heute „dominant“ verwendet). Erst 1900 wurden seine Erkenntnisse von den Botanikern Carl Correns (Tübingen), Erich Tschermak-Seysenegg (Wien), William Bateson (London) und Hugo de Vries (Amsterdam) unabhängig voneinander wiederentdeckt. Zwischenzeitlich waren die Chromosomen und ihre Verteilung an die Nachkommen beschrieben worden, so dass die mendelschen Regeln jetzt mit der Chromosomentheorie der Vererbung in Verbindung gebracht werden konnten. Heute gehören sie zum Gemeingut der klassischen Genetik.
Mendel entdeckte Gesetzmäßigkeiten, die anderen zuvor verborgen geblieben waren. Der Erfolg seiner Untersuchungen lässt sich im Nachhinein mit folgenden Faktoren begründen:[2]
- die Wahl des Versuchsobjekts Erbse (da bei ihr die betrachteten Merkmale auf unterschiedlichen Chromosomen lagen und somit keine Genkopplung auftrat),
- die Nutzung reinerbiger Stämme,
- der Schutz vor Fremdbestäubung,
- die Beschränkung auf wenige Merkmale,
- die großangelegten Versuchsreihen sowie
- die statistische Auswertung.
Die mendelschen Regeln
Die folgenden Regeln wurden ursprünglich durch Beobachtung und statistische Analyse festgestellt. Ihr Verständnis wird jedoch stark erleichtert, wenn man sich vor Augen hält, dass die betroffenen Merkmale von (nur) einem Gen (Erbanlage) festgelegt werden, welches in zwei Kopien vorliegt, von denen je eine von einem Elternteil vererbt wurde.
Regel 1: Uniformitätsregel
Diese Regel (auch Reziprozitätsregel genannt) gilt, wenn zwei Individuen („Eltern-“ oder Parentalgeneration P genannt) gekreuzt werden, die sich in einem Merkmal unterscheiden, für das sie beide jeweils homozygot (reinerbig) sind: Die Nachkommen der ersten Generation („Tochter“ oder erste Filialgeneration F1 genannt) sind dann uniform, d. h. bezogen auf das untersuchte Merkmal untereinander gleich. Dies gilt für den Phänotyp (äußeres Erscheinungsbild) wie den Genotyp (Erbausstattung), welcher bei allen heterozygot (mischerbig) ist. Dabei ist es egal, welches der beiden Allele von der Mutter und welches vom Vater vererbt wird (reziproke Kreuzung).
Für die Ausprägung des Merkmals tritt eine von zwei Möglichkeiten ein (siehe auch die oberen Hälften der Abbildungen 1 und 2):
- Beim dominant-rezessiven Erbgang haben alle Mitglieder der F1-Generation die gleiche Ausprägung wie einer der Eltern. Beispiel: Bei Erbsen ist die rote Blütenfarbe dominant gegenüber der weißen, welche daher als rezessiv bezeichnet wird. Wenn reinerbige rotblühende und reinerbige weißblühende Individuen gekreuzt werden, haben alle Mitglieder der F1-Generation eine Erbanlage (ein Gen) für weiß (von einem Elternteil) und eine Erbanlage für rot (vom anderen Elternteil) vererbt bekommen, sie sind heterozygot. Trotzdem sind sie alle rotblühend, eben weil rot gegenüber weiß dominant ist. Die verschiedenen Formen einer Erbanlage (im Beispiel die Genformen für „rot“ und „weiß“) werden auch als Allele bezeichnet.
- Beim intermediären Erbgang haben alle Mitglieder der F1-Generation eine Mischform der elterlichen Merkmale. Ein Beispiel ist die Blütenfarbe von Mirabilis jalapa: Wenn rot- und weißblütige Exemplare gekreuzt werden, so haben alle Nachkommen rosafarbene Blüten.
Ausnahmen von der 1. Regel können auftreten, wenn sich das Gen für ein untersuchtes Merkmal auf einem Geschlechtschromosom (Gonosom) befindet. Dann kann es sein, dass die F1-Generation nicht uniform ist.
Regel 2: Spaltungsregel
Diese Regel (auch Segregationsregel genannt) gilt, wenn zwei Individuen gekreuzt werden, die beide gleichartig heterozygot sind, also z. B. zwei Pflanzen, die für die Blütenfarbe beide die beiden Erbanlagen „weiß“ und „rot“ haben. Das kann etwa die F1-Generation des vorherigen Abschnitts sein. In Beschreibungen der mendelschen Regeln werden die Nachkommen einer solchen Heterozygoten-Kreuzung daher meist als Enkel oder zweite Filialgeneration, F2, bezeichnet. Diese Nachkommen sind untereinander nicht mehr uniform, sie spalten sich bezüglich der Merkmalsausprägung auf. Dabei kommen die Merkmale der P-Generation (siehe vorheriger Abschnitt) wieder zum Vorschein:
- Handelt es sich um eine dominant-rezessive Vererbung, so sind ein Viertel der F2-Individuen reinerbig mit zwei rezessiven Erbanlagen und zeigen eine entsprechende Merkmalsausprägung (z. B. weiße Erbsenblüten). Die anderen drei Viertel zeigen eine Ausprägung wie reinerbige Individuen mit zwei dominanten Erbanlagen. (Verhältnis von 3:1). Diese drei Viertel setzen sich zusammen aus reinerbigen (ein Viertel) und mischerbigen (zwei Viertel) Individuen.
- Bei intermediärer Vererbung weist je ein Viertel der Nachkommen eine der beiden reinerbigen Varianten und die Hälfte der Individuen die Mischform der 1. Generation auf (Verhältnis von 1:2:1).
1. + 2. Regel im dominant rezessiven Erbgang 1. + 2. Regel im intermediären Erbgang 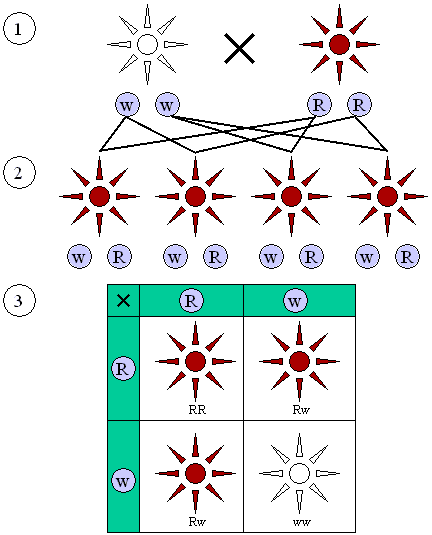 Abbildung 1: Dominant-rezessiver Erbgang wie er z. B. bei der Blütenfarbe der Erbse auftritt.
Abbildung 1: Dominant-rezessiver Erbgang wie er z. B. bei der Blütenfarbe der Erbse auftritt.
(1) Elterngeneration mit reinerbigen Anlagen (w/w oder R/R).
(2) F1 Generation: Alle Individuen sehen gleich aus, die dominante rote Erbanlage setzt sich gegen die rezessive weiße durch.
(3) F2 Generation: Dominante (rot) und rezessive (weiße) Erscheinungsformen zeigen ein Verhältnis von 3:1.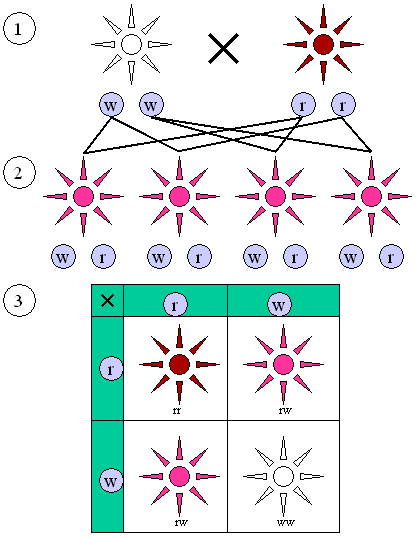 Abbildung 2: Intermediärer Erbgang wie er z. B. bei der Blütenfarbe der Wunderblume Mirabilis jalapa auftritt.
Abbildung 2: Intermediärer Erbgang wie er z. B. bei der Blütenfarbe der Wunderblume Mirabilis jalapa auftritt.
(1) Elterngeneration mit reinerbigen Anlagen (w/w oder r/r).
(2) F1 Generation:Alle Individuen sehen gleich aus, Die „roten“ und „weißen“ Erbanlagen ergeben eine rosa Blütenfarbe.
(3) F2 Generation. Rote, rosa und weiße Blütenfarben treten mit einem 1:2:1 Verhältnis auf.Regel 3: Unabhängigkeitsregel/Neukombinationsregel
Diese Regel beschreibt das Vererbungsverhalten von zwei Merkmalen (z. B. Samenfarbe und Samenform) bei der Kreuzung reinerbiger Individuen und deren Nachkommen. Beide Merkmale werden unabhängig (daher der Name der Regel) voneinander vererbt, wobei ab der F2-Generation („Enkel“) neue, reinerbige Kombinationen auftreten (siehe Abbildung 3).
 Abbildung 3: Zwei Merkmale (weißes/braunes Haar und kurzer/langer Schwanz, wobei „braun“ und „kurz“ dominant sein sollen) zeigen in der F2-Generation im Phänotyp ein Verhältnis von 9:3:3:1. (S = kurz (short), s = lang, B = braun, b = weiß)
Abbildung 3: Zwei Merkmale (weißes/braunes Haar und kurzer/langer Schwanz, wobei „braun“ und „kurz“ dominant sein sollen) zeigen in der F2-Generation im Phänotyp ein Verhältnis von 9:3:3:1. (S = kurz (short), s = lang, B = braun, b = weiß)
oben: Eltern-Generation,
mitte: F1-Generation,
unten: F2-Generation
Ergebnis:
9 × kurzer Schwanz, braunes Haar
3 × langer Schwanz, braunes Haar
3 × kurzer Schwanz, weißes Haar
1 × langer Schwanz, weißes Haar
Zu beachten ist, das es sich bei beiden Merkmalen um dominant-rezessive Erbgänge handelt. Werden ein oder beide Merkmale intermediär vererbt, so werden die Phänotypen nicht im Verhältnis 9:3:3:1 ausgebildet. Im Genotyp bleiben die Verhältnisse bei allen Erbformen gleich. Alle Verhältnisse lassen sich durch Ausrechnen der obigen Matrix (Punnett-Quadrat) leicht ermitteln.Diese Regel gilt allerdings nur dann, wenn sich die für die Merkmale verantwortlichen Gene auf verschiedenen Chromosomen befinden oder wenn sie auf dem gleichen Chromosom so weit voneinander entfernt liegen, dass sie während der Meiose durch Crossing over regelmäßig getrennt voneinander vererbt werden (polygene Erbgänge). Befinden sich Gene auf dem gleichen Chromosom nahe beieinander, so werden sie in Kopplungsgruppen vererbt.
Genetische Hintergründe
Mendel kannte weder den Begriff der Gene noch den der Chromosomen. Die stoffliche Grundlage der Erbanlagen war ihm somit nicht bekannt. Erst 1904 wurde durch Walter Sutton und Theodor Boveri die Chromosomentheorie der Vererbung begründet. Mit Hilfe dieser Theorie können die Mendelregeln widerspruchsfrei erklärt werden.
In den Körperzellen treten die Chromosomen paarweise auf (Ausnahmen bilden viele Pflanzen und niedere Tiere). So besitzt der Mensch 23 Chromosomenpaare, also 46 Chromosomen. Man spricht von einem doppelten oder diploiden Chromosomensatz. Je ein Chromosomensatz stammt vom Vater und einer von der Mutter. Die Chromosomen eines Paares werden auch als homologe Chromosomen bezeichnet. In einer Körperzelle sind die Erbanlagen pro Merkmal somit immer doppelt vorhanden.
Bei der Bildung der Geschlechtszellen werden die homologen Chromosomenpaare in der Meiose getrennt. In einem Ei bzw. einem Spermium (Geschlechtszelle) befindet sich also nur der einfache Chromosomensatz, die Erbanlagen pro Merkmal (Genabschnitte) sind somit immer nur einmal vorhanden. Dies erklärt die Spaltungsregel.
Bei der Befruchtung, also der Verschmelzung von Ei und Spermium, bringen beide Geschlechtszellen jeweils eine Erbanlage pro Merkmal mit. Die durch diese Verschmelzung entstehende Körperzelle (Zygote) hat also wieder den doppelten Chromosomensatz. Die Erbanlagen können so neu kombiniert werden (Unabhängigkeitsregel, freie Kombination der Gene).
Der Nutzen dieser Rekombination – also der sexuellen Vermehrung – liegt darin, dass mehr verschiedene Genkombinationen zur Evolution beitragen. Ohne sexuelle Vermehrung gäbe es von jedem Lebewesen nur weitgehend identische Kopien. Mit sexueller Vermehrung gibt es eine hohe Bandbreite an Nachkommen, und damit an Überlebensstrategien und -möglichkeiten. Dazu kommt, dass Fehler oder Schwächen im Erbgut bei sexueller Vermehrung sehr ungleichmäßig auf die Nachkommen verteilt sind, sodass einige Nachkommen deutlich weniger Probleme haben als ihre Eltern, und damit ein Ausgleich für Fehler beim Kopieren der Erbinformationen geschaffen wird. Außerdem ermöglicht die sexuelle Vermehrung die Übernahme der besten Eigenschaften zweier zeitweise getrennter Populationen in eine gemeinsame Population und damit auch eine schnellere Aneignung von Fähigkeiten, die woanders vorher schon entstanden sind. Insbesondere bei neuen Genen (z. B. Malariaresistenz) tritt auch das Phänomen auf, dass ein Gen mit Nebenwirkungen (in diesem Fall Sichelzellenanämie) nicht alle ihre Träger krank macht, sodass der Vorteil trotz Nachteilen („Unausgereiftheit“ der Mutation) vererbt werden kann. Unter „natürlichen“ Bedingungen (Selektion zugunsten der Gesünderen) sollten weitere Mutationen nach einigen Generationen dazu führen, dass die Nachteile weniger werden. Der technologische Fortschritt verlagert diese Entwicklung aber mehr und mehr in die Medizin.
Anwendung
Die mendelschen Regeln werden insbesondere in der Tier- und Pflanzenzucht angewendet, z. B. bei der Zucht von Hybriden. Sie können auch für Abstammungsgutachten verwendet werden, z. B. um nachzuweisen, dass bestimmte Menschen nicht als Elternteil eines bestimmten Kindes in Frage kommen.
Einzelnachweise
- ↑ Gregor Mendel: Versuche über Pflanzenhybriden. Verhandlungen des Naturforschenden Vereines in Brünn. Bd. IV. 1866. S. 3–47.
- ↑ Abiturwissen Biologie. Bibliographische Informationen der Deutschen Bibliothek, 2004. ISBN 3-411-00222-0.
Literatur
- H. Frederik Nijhout: Der Kontext macht’s!. Spektrum der Wissenschaft, April 2005, S. 70–77. ISSN 0170-2971.
Siehe auch
Weblinks
- Versuche über Pflanzen-Hybriden (1865): Mendels Original-Veröffentlichung (vorgelegt in den Sitzungen vom 8. Februar und 8. März 1865) bei mendelweb.org, Roger B. Blumberg
- Vererbungslehre im Bezug auf Zierfische bei Das Zierfischverzeichnis, Anika Börries
- Computersimulation zu den mendelschen Regeln von Hermann Mendel

Wikimedia Foundation.