- CpG-Inseln
-
CpG-Inseln (engl.: CpG-islands) sind genomische Regionen mit statistisch erhöhter CpG-Dinukleotid-Dichte. Diese Dichte wird auf die Einzelnukleotid- und Dinukleotidfrequenzen im gesamten betrachteten Genomausschnitt bezogen. CpG bedeutet Cytosin-phosphatidyl-Guanosin. Das p wird mit angegeben, um GC von CG innerhalb von Phosphonukleosidketten (=DNA-Molekülen) besser zu unterscheiden (siehe auch 5'-Ende, 3'-Ende). CpG-Inseln sind definiert als DNA-Abschnitte von 0,5kb bis 2kb Länge, die einen erhöhten GC-Gehalt von über 60% aufweisen, dabei liegt der GC-Gehalt des Gesamtgenoms bei 40%. CpG-Inseln entstehen durch Mechanismen, die mit der Nutzung der Erbsubstanz als Informationsträger zu tun haben. Dadurch sind CpG-Inseln wichtige Markierungen, die z. B. für die Genetik, Medizin und Bioinformatik Bedeutung haben.
Sie sind nicht zu verwechseln mit der GC-Box, die 60–100 bp vor Beginn des Transkripts liegt.
Inhaltsverzeichnis
Entstehung und Bedeutung
Bei Säugetieren sind je nach Spezies zwischen 2% und 7% der Cytosine einer Zelle methyliert. Meist sind es die Cytosine aus 5'-CpG-3' Dinukleotiden, die auf beiden komplementären DNA-Strängen eine Methylgruppe tragen, wodurch ein palindromisches Methylierungsmuster entsteht. Sind zwei Cytosine in dieser Konstellation methyliert, bewirken sie zusammen eine Veränderung der dreidimensionalen Struktur in der großen Furche der Doppelstrang-DNA. Ein Großteil (etwa 60-90%) aller CpGs des Genoms von Säugetieren sind methyliert, wobei unmethylierte CpG-Dinukleotide vorwiegend innerhalb von CpG-Inseln anzutreffen sind.
Der durchschnittliche GC-Gehalt beim Menschen beträgt 41%, womit das Dinukleotid CpG rechnerisch mit einer Häufigkeit von 4% im Genom vorliegen sollte. Tatsächlich sind aber CpG-Dinukleotide mit 0,8% stark unterrepräsentiert, was hauptsächlich mit der relativ spontanen Reaktion von methyl-Cytosin zu Thymin durch Desaminierung zu erklären ist (s. Erklärung und Abbildung weiter unten). Damit ist die CpG-Dinukleotiddichte in CpG-Inseln 10-20 mal höher als in anderen Bereichen des durchschnittlichen Genoms von Wirbeltieren.
CpG-Inseln werden auch zur Regulation der Expression (des Ablesens) der Gene verwendet, und sind damit ein Mechanismus der epigenetischen Genregulation. Methylierung der CpG-Inseln eines Genes bedeutet, dass diese Gene nicht abgelesen werden (Gen-Silencing). Circa 60 % aller menschlichen Gene haben CpG-Inseln in ihren Promotorbereichen.
Methylierung von CpG-Inseln spielt sowohl in der Entstehung von Krebs (als Mechanismus zum Abschalten von Tumorsuppressorgenen) als auch beim genomischen Imprinting eine Rolle.
Die beiden Cytosine in einem CpG-Dinukleotid sind im menschlichen Genom meist methyliert (DNA-Methylierung). In einigen Bereichen wird die Methylierung dauerhaft unterdrückt. Häufig sind diese Bereiche CpG-Inseln und liegen oft vor Genen (den sogenannten Promotorbereichen). Die CpG dieser Regionen sind meist nicht methyliert und entgehen dadurch einem Mutationsdruck, der nachfolgend beschrieben wird:Cytosine sind chemisch labil. Sie können in der Zelle einer oxidativen Desaminierung (aus -NH2 wird =O ) unterliegen. Aus methyliertem Cytosin wird dabei Thymin, aus unmethyliertem Cytosin (z. B. in den CpG-Inseln) wird Uracil. Während Thymidin eine "normale" Nukleobase der DNA ist, gehört Uracil nicht in die DNA. Uracil -eigentlich eine RNA-Base- wird sehr gut erkannt und durch Cytosin ersetzt. Die DNA-Reparaturmechanismen der Zelle nehmen das auf dem gegenüberliegenden DNA-Strang vorhandene Guanosin als Grundlage der Fehlerkorrektur. In den methylierten CpG-Dinukleotiden entsteht durch die Desaminierung hingegen Thymidin. Dieser "Fehler" wird wesentlich häufiger toleriert als Uracil und führt zu einer dauerhaften Mutation.
Das folgende Schema zeigt die möglichen Mutationen durch Desaminierung und die Folgen durch Reparatur der DNA bzw. durch dauerhafte Etablierung von Mutationen.
1. 2. 3. | Methyliert: | m | m a) --CpG-- Desaminierung --TpG-- häufig --CpG-- | -> --CpG-- --GpC-- --GpC-- --GpC-- | --GpC-- m m m | m | | b) selten --TpG-- | -> --TpG-- --ApC-- | --ApC-- m | Unmethyliert: | | c) --CpG-- Desaminierung --UpG-- sehr häufig --CpG-- | --GpC-- --GpC-- --GpC-- | | | | d) sehr selten --UpG-- | -> --TpG-- --ApC-- | --ApC-- |Legende zum Schema: Dargestellt sind zwei CpG-Dinukleotide, von denen eins sich in einem methylierten Bereich befindet [a) und b)], während das andere in einem unmethylierten Bereich -z. B. einer CpG-Insel- lokalisiert ist [c) und d)]. Die "auffälligen" Nukleobasen sind fett vervorgehoben.
1. Eine Desaminierung führt zu einem neuen Dinukleotid, in welchem die komplementäre Basenpaarung aufgehoben ist.
2. Für die nachfolgende Wiederherstellung der komplementären Basenpaarung stehen jeweils zwei Varianten zur Verfügung, die mit unterschiedlicher Wahrscheinlichkeit verlaufen. Der Unterschied zwischen a) und b) mit "häufig" und "selten" kommt dadurch zustande, dass der gegenüberliegende Strang eine Methylierung des CpG aufweist. Dadurch wird dieser Strang in diesem Bereich vom DNA-Reparatursystem als "älterer", konservierter Strang verstanden. Der größere Unterschied zwischen c) und d) mit "sehr häufig" und "sehr selten" geht darauf zurück, dass Uracil keine DNA-Base ist.
3. Im Anschluss an die mutativen Ereignisse werden gegebenenfalls falsche Methylierungen oder Nukleobasen ersetzt.
Bioinformatische Analyse
Auffinden von CpG-Inseln mit Hilfe von Markov-Ketten
Bezeichnet
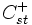 die Anzahl der st-Paare auf CpG-Inseln und
die Anzahl der st-Paare auf CpG-Inseln und 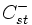 sonst (nicht CpG-Inseln) mit
sonst (nicht CpG-Inseln) mit 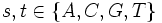 . Die Übergangswahrscheinlichkeiten werden über Maximum Likelihood berechnet:
. Die Übergangswahrscheinlichkeiten werden über Maximum Likelihood berechnet: 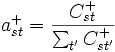 und
und 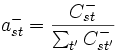 Die Bestimmung basiert auf Sequenzabschnitten, von denen man weiß, ob es sich um CpG-Inseln handelt oder nicht. Gegeben sei nun eine unbekannte Sequenz X. Frage: "Handelt es sich um eine CpG-Insel?" Bezeichnungen:
Die Bestimmung basiert auf Sequenzabschnitten, von denen man weiß, ob es sich um CpG-Inseln handelt oder nicht. Gegeben sei nun eine unbekannte Sequenz X. Frage: "Handelt es sich um eine CpG-Insel?" Bezeichnungen:- P(+|X) Wahrscheinlichkeit, dass X CpG-Insel
- P(-|X) Wahrscheinlichkeit, dass X keine CpG-Insel
Zusätzlich wird eine Score-Funktion definiert:
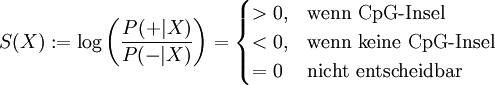
Als "Prior" wird die Gesamtlänge aller CpG-Inseln relativ zur Gesamtlänge des Genoms verwendet.
Auffinden von CpG-Inseln mit Hilfe des Hidden Markov Modells
Als sichtbare Zustände bezeichnet man hierbei die Basen (G,C,A,T) an den jeweiligen Stellen in der DNA-Sequenz. Der nicht-sichtbare Zustand sagt etwas darüber aus, ob diese Base Teil einer CpG-Insel ist oder nicht (+,-). Es gibt 4 mögliche Übergangswahrscheinlichkeiten:
ast = P(Zi = t | Zi + 1 = s)
 .
.Jeder versteckte Zustand s erzeugt mit einen Emissionswahrscheinlichkeit es(b) einen sichtbaren Zustand b (eine Base):
es(b) = P(Xi = b | Zi = s)
Die Wahrscheinlichkeit, dass ein sichtbarer Zustand von einem versteckten Zustand emittiert wurde ergibt sich demnach aus: P(Z | X) = P(X | Z) * P(Z) / P(X)
mit:
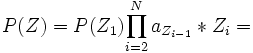
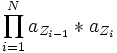 (s. Markov-Kette)
(s. Markov-Kette)Damit ergibt sich:
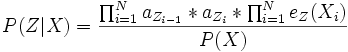
Da der Aufwand zur Maximierung von P(Z | X) mit der Länge der Sequenz exponentiell steigt, eignet sich der rekursive Viterbi-Algorithmus zur Lösung des Problems.
Wikimedia Foundation.