- DNA-Methylierung
-
Übergeordnet epigenetische Regulation der Genexpression
Methylierung von Makromolekülen
DNA-ModifikationGene Ontology AmiGO QuickGO Bei der DNA-Methylierung handelt es sich als wichtigste epigenetische Veränderung um eine chemische Abänderung an Grundbausteinen der Erbsubstanz einer Zelle. Diese Abänderung (Modifikation) wird durch die Übertragung von Methylgruppen durch Enzyme (DNA-Methyltransferasen) auf Nukleobasen an bestimmten Stellen innerhalb der DNA hervorgerufen. Da der jeweilige Grundbaustein an der jeweiligen Stelle erhalten bleibt, ist die DNA-Methylierung keine genetische Mutation. Die DNA-Methylierung ist eine DNA-Modifikation durch Methylierung der DNA; sie kommt in sehr vielen verschiedenen (möglicherweise in allen) Lebewesen vor und hat verschiedene biologische Funktionen. Die Abfolge der DNA-Methylierung ist Teil des epigenetischen Codes einer Zelle.
Epigenetische Veränderung
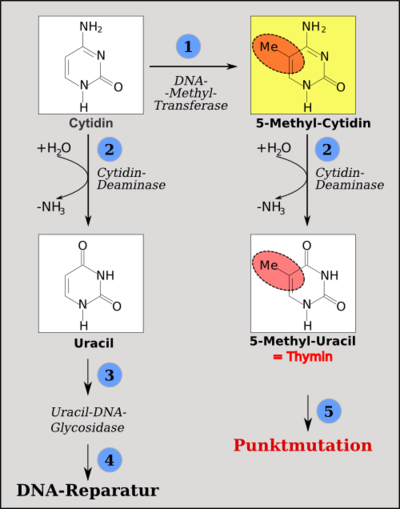 Folgen der Desaminierung: Cytidin kann durch die DNA-Methyltransferasen methyliert werden (1). Wird Cytidin desaminiert, entsteht Uracil (2). Dieses wird als DNA-fremd vom DNA-Reparatur-Apparat ausgetauscht (3). Wird 5-Methylcytidin desaminiert, entsteht Thymin (4). Dieses ist ein DNA-Baustein und die Thymin-Guanosin-Fehlpaarung wird durch Austausch von Thymin gegen Cytidin (Wiederherstellung des Ausgangszustandes) oder durch Austausch von Guanosin gegen Adenosin entfernt, was zu einer C→T-Punktmutation führt (5).
Folgen der Desaminierung: Cytidin kann durch die DNA-Methyltransferasen methyliert werden (1). Wird Cytidin desaminiert, entsteht Uracil (2). Dieses wird als DNA-fremd vom DNA-Reparatur-Apparat ausgetauscht (3). Wird 5-Methylcytidin desaminiert, entsteht Thymin (4). Dieses ist ein DNA-Baustein und die Thymin-Guanosin-Fehlpaarung wird durch Austausch von Thymin gegen Cytidin (Wiederherstellung des Ausgangszustandes) oder durch Austausch von Guanosin gegen Adenosin entfernt, was zu einer C→T-Punktmutation führt (5).
Die wichtigste epigenetische Veränderung ist die Methylierung von Cytidin-Basen der DNA.[1] Dabei werden überhaupt nur solche Cytidine methyliert, die innerhalb von Cytosin-Guanosin-Dinukleotiden angetroffen werden. Andere Cytidine werden durch die bekannten menschlichen DNA-Methyltransferasen (DNMT) nicht verändert.[2]
Während der DNA-Verdopplung vor jeder Zellteilung gibt es den alten DNA-Strang, an dem bestimmte Cytidine methyliert sind, während der neugebildete DNA-Strang noch nicht methyliert ist. Das Enzym DNMT3 methyliert jedes Cytidin in einem halbmethylierten CG/CG-Paar. Eine solche CG-Methylierung führt dazu, dass Methyl-CG-erkennende Proteine an solche meCG-Paare binden. Diese Bindung führt zur Anlagerung weiterer Proteine und zur Verdichtung der Nukleosomen (siehe weiter unten). Dadurch ist die DNA an solchen meCG-Paaren für die RNA-Polymerase nicht ablesbar und das darunterliegende Gen ist inaktiv.
Methylierte Cytidine sind anfällig für Desaminierung, dabei verlieren die Cytidine die Aminogruppe an Position 4 des Ringes. Ein desaminiertes, nichtmetyliertes Cytidin ist ein Uracil. Dieses ist keine der vier normalen DNA-Basen Adenin, Cytosin, Guanin oder Thymin. Daher wird ein Uracil in der DNA als Fehler erkannt und schnellstens ausgetauscht. Wird aber ein 5-Methylcytidin desaminiert, entsteht daraus ein Thymin, das ein DNA-Baustein ist. Hier kann der DNA-Reparaturapparat nicht erkennen, ob das Thymin oder das gegenüberliegende Guanin falsch eingebaut ist. Daher bleibt die Umwandlung eines Methylcytidins in ein Thymin erhalten und wird, wenn diese Methylierung in einer Keimzelle stattgefunden hat, auch vererbt.
Wenn man auszählt, wie häufig Zweierpaare von Nukleotiden in einer Nukleinsäure vorkommen, stellt man fest, dass alle Dinukleotide, z.B. Adenin-Thymin (AT), Thymin-Guanidin (TG), Guanidin-Thymin (GT) und die anderen 12 Paare etwa gleich häufig sind. Nur das Cytidin-Guanidin-Paar (CG) kommt wesentlich seltener vor. Der Grund dafür ist genau, dass das Cytidin methyliert werden kann, eine Desaminierung stattfand, die nicht repariert wurde, und daher die Häufigkeit von CG verringert wird. Dieser Vorgang ist auch nicht abgeschlossen, sondern könnte immer wieder neu vonstatten gehen.
Die erhalten gebliebenen CG-Dinukleotide treten gehäuft vor allem in den Genbereichen auf, die für die Steuerung von Genen zuständig sind, den Promotoren. Man spricht von einer CpG-Insel (Cytosin phosphat Guanin - Insel). Würden hier CG- in TG-Paare umgewandelt, könnten Zellfunktionen verändert werden oder verloren gehen. Wenn eine solche Veränderung die Existenz der Zelle bzw. des Embryos gefährdet, findet Selektion gegen die Veränderung statt und die Veränderung wird nicht vererbt. Der Methylierungsgrad in solchen CpG-Inseln ist häufig ein Maß für die Aktivität, mit der das zugehörige Gen transkribiert wird: Je geringer die Methylierung, desto eher wird das zugehörige Gen abgelesen. Eine verstärkte Methylierung in dem Promotor eines Genes verhindert also die Aktivität in diesem Gen.
DNA-Methyltransferasen (DNMT)
Bislang sind drei menschliche DNA-Methyltransferasen bekannt: DNMT1, DNMT3a und DNMT3b (DNMT2 methyliert RNA). Für die Erhaltungs-Methylierung (Maintenance-Methylierung) bei der Zellteilung ist DNMT1 zuständig. DNMT3a und DNMT3b methylieren die CG-Dimere, die aufgrund von Zelldifferenzierungen neu methyliert werden (de-novo-Methylierung). Mutationen im DNMT3b-Gen auf Chromosom 20 führen zum Immunschwäche/zentromere Instabilität/Gesichtsausdrucks-Anomalie-Syndrom (ICF). An methylierte DNA kann sich das Methyl-bindende Protein (MeCP) anlagern. Dieses wiederum ist Keim für weitere Proteinanlagerungen, die schließlich auch zur Modifizierung von Histonen führen. Kondensiertes Histon in Zusammenarbeit mit dem durch MeCP ausgelösten Proteinkomplex führt zur Inaktivierung eines Chromosomenabschnittes.
DNA-Methylase
Auch das Methyl-abspaltende Enzym DNA-Methylase wurde identifiziert.[3] Es war als Methyl-CpG-Domäne-bindendes Protein 2 (MBD2) schon früher beschrieben worden. Damit ist die Methylierung von DNA keine Einbahnstraße, sondern der Methylierungszustand kann zellfunktionsabhängig geregelt werden. Eine solche Situation nennt man plastisch.
DNA-Methylierung und Schutz vor fremder DNA
DNA ist weit verbreitet. Zellen unterschiedlicher Arten können in unmittelbarer Nachbarschaft existieren. Sowohl bei der Nahrungsaufnahme einer Zelle (z. B. Phagozytose) wie auch bei parasexuellen und sexuellen Prozessen kommt es zur Aufnahme der DNA von einer (lebenden oder toten) Zelle in eine andere Zelle. Darüber hinaus sind viele Zellen in der Lage, unter bestimmten Umständen Fremd-DNA leicht aufnehmen zu können (Zellkompetenz).
Da eine lebende Zelle ihre Integrität nur erhalten kann, wenn die genetische Information sinnvoll ist, sollte sie in der Lage sein, fremde DNA zu erkennen und zu eliminieren. Dies wird häufig durch ein System aus zwei Enzymgruppen gewährleistet: Die DNA-Methyltransferasen und die Restriktionsendonukleasen.
Die Methyltransferasen erkennen eine (meist kurze) DNA-Sequenz und hängen eine Methylgruppe an eine definierte Nukleobase. Dadurch entsteht ein sogenanntes Methylierungsmuster. Die Restriktionsenzyme erkennen ebenfalls eine (meist kurze) DNA-Sequenz und trennen die DNA an definierten Stellen zwischen Phosphat und Desoxyribose. Viele Restriktionsenzyme sind methylierungssensitiv. Das heißt, sie zerschneiden die DNA nur, wenn an bestimmten Stellen Methylierungen vorliegen oder wenn an bestimmten Stellen keine Methylierungen vorhanden sind.
Das System aus Methyltransferasen und Restriktionsenzymen ist in einer lebenden Zelle so abgestimmt, dass die eigene DNA nicht zerschnitten wird. Fremde DNA, die von außen in die betrachtete Zelle gelangt, hat jedoch in den allermeisten Fällen ein anderes Methylierungsmuster. Daher wird sie mit hoher Wahrscheinlichkeit von den Restriktionsenzymen sowie anderen Nukleasen verdaut. In seltenen Fällen wird fremde DNA nicht oder nur zum Teil verdaut und dauerhaft in die zelleigene DNA integriert. Eine Integration fremder DNA wird auch als horizontaler Gentransfer bezeichnet und ist ein Motor der Evolution.
Nachfolgend wird ein einfaches System aus Methyltransferase und Restriktionsenzym als Beispiel erläutert.
Beispiel für DNA-Methylierung und DNA-Restriktion
Das Zusammenwirken von DNA-Methylierung und DNA-Restriktion (Spaltung von DNA) soll anhand der Enzyme DpnM (DNA-Methyltransferase) und DpnII (Restriktionsenzym) beschrieben werden. Die Enzyme stammen aus dem Bakterium Diplococcus pneumoniae. Die Methyltransferase DpnM sorgt dafür, dass die palindromische Sequenz GATC im Adenosin methyliert wird:
m --GATC-- --CTAG-- mDadurch kann „frische DNA“ die gerade neu entstanden ist, von der alten DNA, die als Vorlage gedient hat, unterschieden werden:
m --GATC-- --CTAG--Das ist für die korrekte Reparatur von Fehlern während der DNA-Replikation wichtig. Der sogenannte hemimethylierte Zustand (eine Seite ist methyliert, die andere nicht) wird nachfolgend durch die Methyltransferasen – wie z. B. DpnM – durch Methylierung aufgehoben. Sollte DNA einer anderen Art in die D. pneumoniae-Zelle gelangen, so ist diese DNA in der Sequenz GATC meist nicht methyliert:
--GATC-- --CTAG--Diese doppelsträngige DNA wird mit großer Wahrscheinlichkeit vom Restriktionsenzym DpnII zerschnitten. Neben anderen Prozessen führt das dazu, dass fremde DNA eher als Nahrung und weniger als Erbsubstanz dient. Dieses ist außerdem ein Mechanismus, mit dem sich Bakterien vor Bakteriophagen schützen – indem sie die eingebrachte DNA in kleine Stücke schneiden.
DNA-Methylierung und Fehlerkorrektur bei der DNA-Neusynthese
Die identische Verdopplung der Desoxyribonukleinsäure (DNA-Replikation) ist eine wesentliche Voraussetzung für die Zellteilung und damit für die Vermehrung. Die DNA-Replikation wird dadurch gewährleistet, dass Enzyme (die DNA-abhängigen DNA-Polymerasen) den bereits vorhandenen alten Strang „lesen“ und dabei den neuen Strang „schreiben“. Dabei können Fehler auftreten. Die Fehlerstellen können durch die DNA-Reparatursysteme einer Zelle erkannt werden, da dort keine komplementäre Basenpaarung vorliegt. Allerdings bliebe unklar, welche der beiden Möglichkeiten die richtige sei, wenn sich alter und neuer DNA-Strang nicht unterschieden. Da der alte Strang jedoch methyliert ist, der neue aber noch nicht, ist eine Unterscheidung möglich. Die DNA-Reparatursysteme können das Methylierungsmuster zur Fehlerkorrektur nutzen. Das wird im Folgenden anhand der Methylierung des CpG-Dinukleotids verdeutlicht.
In den Zellen des Menschen wie auch anderer Säugetiere ist eine kurze Folge aus zwei Grundbausteinen (Nukleosiden) die Grundlage für eine DNA-Methylierung: Cytidin – Phosphorsäure – Guanosin. CpG ist in der Reihenfolge der Synthese von Nukleinsäuren angegeben. Das Phosphat stammt vom Baustein Guanosintriphosphat, der für die DNA-Synthese verwendet wurde. In der Aufeinanderfolge der beiden Nukleobasen Cytosin–Guanin wird das Cytosin methyliert. Bis auf einige Bereiche passiert dies fast in der gesamten menschlichen Erbsubstanz.
1. Vor der DNA-Replikation sind im betrachteten Beispielabschnitt die CpG-Dinukleotide in beiden Strängen am Cytosin methyliert:
m m ...pApApCpTpCpGpTpGpCpApApApCpGpGpTpT ... | | | | | | | | | | | | | | | | | ... TpTpGpApGpCpApCpGpTpTpTpGpCpCpApAp... m m2. Während der DNA-Replikation kommt es zum Fehler: Statt eines Thymidintriphosphat-Moleküls wird ein Cytidintriphosphat verwendet. Dadurch wird an der entsprechenden Stelle die komplementäre Basenpaarung aufgehoben:
m m ...pApApCpTpCpGpTpGpCpApApApCpGpGpTpT ... | | | | | | | | | | | CpApCpGpCpTpTpGpCpCpApAp...3. Nach Abschluss der DNA-Replikation im Beispielabschnitt liegt der DNA-Doppelstrang hemimethyliert vor. Das heißt, der alte Strang ist methyliert, der neue nicht.
m m ...pApApCpTpCpGpTpGpCpApApApCpGpGpTpT ... | | | | | | | | | | | | | | | | ... TpTpGpApGpCpApCpGpCpTpTpGpCpCpApAp...4. Die Stelle der Fehlpaarung wird erkannt. Von den beiden Möglichkeiten Adenosinmonophosphat und Cytidinmonophosphat wird das Cytidinmonophosphat ausgeschnitten, das im nichtmethylierten Strang liegt. Tymidintriphosphat wird verwendet, um die Lücke zu schließen:
m m ...pApApCpTpCpGpTpGpCpApApApCpGpGpTpT ... | | | | | | | | | | | | | | | | ... TpTpGpApGpCpApCpGp TpTpGpCpCpApAp... pppT5. Der reparierte DNA-Doppelstrang liegt im hemimethylierten Zustand vor:
m m ...pApApCpTpCpGpTpGpCpApApApCpGpGpTpT ... | | | | | | | | | | | | | | | | | ... TpTpGpApGpCpApCpGpTpTpTpGpCpCpApAp...6. Durch die Übertragung von Methylgruppen auf die Nukleobase Cytosin in den CpG-Dinukleotiden wird der Grundzustand wiederhergestellt:
m m ...pApApCpTpCpGpTpGpCpApApApCpGpGpTpT ... | | | | | | | | | | | | | | | | | ... TpTpGpApGpCpApCpGpTpTpTpGpCpCpApAp... m mDNA-Methylierung und die Nutzung der DNA als Informationsträger
DNA-Methylierungen sind Markierungen, die es der lebenden Zelle gestatten, Bereiche innerhalb der DNA für verschiedene Prozesse selektiv zu nutzen. Die Markierung von DNA kann ähnlich wie Textformatierungen in einem Buch betrachtet werden: Wenn in einem Lexikon ein Stichwort hervorgehoben dargestellt ist, hat es für den Leser eine andere Bedeutung als dasselbe Wort im Fließtext. Es existieren mehrere (sich überschneidende) Möglichkeiten, wie DNA-Methylierungen die Art der Interpretation von Information variieren, welche in der Bausteinabfolge der DNA gespeichert ist.
DNA-Methylierung und Genregulation
In einem Bereich vor einem Gen (stromaufwärts, upstream) sind häufig Stellen vorhanden, die sich hinsichtlich ihres Methylierungsmusters von der Umgebung unterscheiden. Dabei kann in vielen Fällen der Methylierungsgrad in unterschiedlichen Situationen variieren. Dadurch wird eine selektive Lesehäufigkeit des dahinterliegenden Gens möglich, was man als Genregulation oder differenzielle Genexpression bezeichnet. Beispiele für solche Bereiche, die selektiv methyliert sein können, sind CpG-Inseln.
DNA-Methylierung und Imprinting (genomische Prägung)
Die genomische Prägung ist ein Spezialfall einer differenziellen Genexpression, welche in der Regel durch DNA-Methylierung gesteuert wird. Durch unterschiedliche DNA-Methylierungsmuster in den männlichen und weiblichen Keimzellen können väterliche und mütterliche Allele unterschieden werden. Bei Genen, die dem Imprinting unterliegen, wird nur das mütterliche oder väterliche Allel genutzt. Dadurch ist eine geschlechtsspezifische Ausprägung von phänotypischen Merkmalen möglich.
DNA-Methylierung bei Bakterien
 N6-Methyl-Adenin
N6-Methyl-AdeninBesonders bei Bakterien hat die Adenin-Methylierung eine wichtige Rolle bei der Fehlerkorrektur der frisch replizierten DNA.[4][5] Innerhalb von GATC-Tetrameren wird das Adenin an der 6-Aminogruppe methyliert (vgl. Bild rechts). Manchmal paart ein Thymin mit einem Cytidin anstelle eines Guanins und wird bei der DNA-Verdopplung irrtümlich eingebaut. Diese und andere Fehlpaarungen können von einem Komplex gefunden werden, der den DNA-Strang absucht und eine Fehlerkorrektur (proof-reading) einleitet. Hierbei wird der DNA-Strang um den Fehler herausgeschnitten, der keine methylierten Adenine aufweist. Das ausgeschnittene Stück wird schließlich neu synthetisiert.
Ist das proof-reading abgeschlossen, werden die Adenine im neuen Strang methyliert.
DNA-Methylierung bei Eukaryoten
Anders als bei Bakterien benutzt der DNA-Reparatur-Apparat der Eukaryoten nicht hemimethylierte GATC-Palindrome, um den neuen von dem alten Strang zu unterscheiden, sondern methylierte CG-Dimere. Bei C→T-Punktmutationen außerhalb von CpG-Inseln kann es aber zu Irrtümern kommen, wenn das nächste CG zu weit entfernt ist. Wahrscheinlich sind deshalb die methylierten CG-Dimere nur innerhalb von CpG-Inseln stabil.
CG-Methylierung spielt bei der Promotor-Inaktivierung, der Chromatin-Kondensierung, dem genomischen Imprinting und der X-Chromosom-Inaktivierung eine wichtige Rolle [6] Im Gegensatz zu Hefen, Fliegen wie Drosophila oder Nematoden wie Caenorhabditis elegans finden sich bei Pflanzen und bei den Vertebraten näher verwandten Echinodermata CH3CG-Methylierungen. .
Biologische Funktionen von DNA-Methylierungen
Da die DNA-Methylierung die Nutzung der DNA gestaltet, ohne die Abfolge der Grundbausteine (DNA-Sequenz) zu verändern, ist sie Gegenstand der Epigenetik. Bisher sind im Wesentlichen folgende Bedeutungen der Methylierung von Nukleobasen innerhalb doppelsträngiger DNA bekannt:
- Bei Prokaryoten
- Schutz vor fremder DNA: Unterscheidung zelleigener DNA von solcher, die von außen in die Zelle gelangt ist.
- Fehlerkorrektur bei der DNA-Synthese: Unterscheidung des ursprünglichen (methylierten) DNA-Strangs vom neusynthetisierten Strang, in welchem die Nukleobasen noch nicht methyliert sind.
- Bei Eukaryoten
- Nutzung der DNA als Informationsträger: Markierung von aktiven und inaktiven Bereichen der DNA.
Medizinische Bedeutung
Da fehlerhafte DNA-Methylierungen auf Zellebene reduzierte oder erhöhte Genaktivität bedingen, und diese Aktivitätsveränderungen meist stabil an Tochterzellen vererbt werden, sind sie auf Organismenebene häufig auch Ursache für Krankheiten. So weisen z. B. Tumorzellen oft Methylierungsmuster auf, die von denjenigen gesunder Geweben signifikant abweichen. Ein Tumor kann dabei sowohl als Folge zu starker Methylierung (Hypomethylierung-Hypermethylierung) von upstream DNA-Bereichen entstehen, als auch bei verringertem Methylierungsgrad [7]. Der regulatorische Bereich vor jedem Gen (Promotorbereich) besteht aus verschiedenen typischen DNA-Sequenzen, die spezielle Bindungsstellen für unterschiedliche Enzyme darstellen. Meistens blockiert eine hypermethylierte upstream DNA den Zugang transkriptionsaktiver Faktoren und Enzyme, wodurch die Genaktivität des nachfolgenden Gens supprimiert wird.
Die DNA-Bereiche, die für die Methylierung von besonderer Bedeutung sind, heißen CpG-Inseln. Ihr GC-Gehalt beträgt etwa 60 % (Gesamtgenom: ca. 40 %), und in diesen Abschnitten liegt das Dinukleotid Cytosin-Guanin (5'-CpG-3') im Vergleich zum restlichen Genom mit zehn- bis zwanzigmal erhöhter Frequenz vor. CpG-Inseln dienen in der humangenetischen Forschung oft der Zuordnung von Genen zu genetischen Erkrankungen. Die Gene und die durch DNA-Methylierung gesteuerten Bereiche vor dem jeweiligen Gen können für die Diagnose von vererbbaren Erkrankungen mit molekulargenetischen Methoden eingesetzt werden.
Eine Therapie von Erkrankungen durch eine gezielte Beeinflussung der DNA-Methylierung ist bisher und auf absehbare Zeit nicht möglich – u. a. auch deshalb, weil zu wenig über das ‚richtige‘ Methylierungsmuster gesunder Gewebe bekannt ist. Derzeit gibt es nur experimentelle In vitro-Ansätze, durch sogenannte Zinkfingerproteine (spezielle Klasse von Proteinen, die um ein zentrales Zink-Ion DNA-bindende Domänen besitzen und mit Methylasen oder Demethylasen gekoppelt sein können), um so gezielt bestimmte Sequenzen modifizieren zu können.
Regulation der DNA-Methylierung in Tumoren
- Die Analyse der DNA-Methylierung von Tumorzellen hat ergeben, dass in Tumorzellen häufig die Gene für sogenannte Tumorsuppressorproteine im Vergleich zu Normalzellen methyliert sind.
- So ist in der akuten myeloischen Leukämie (AML) häufig die CG-Insel des P15-Proteins (auch CDKN2B oder ink4b genannt) methyliert.
- P15 ist ein hemmender Regulator des Zellzyklus.
- Nach Bildung von meCG in der CG-Insel von P15 wird dessen Transkription und die Biosynthese des P15-Proteins eingestellt.
- Beim Zellzyklus-Regulator P53 ist in 50 % aller menschlichen Tumoren das P53-Gen hypermethyliert und damit inaktiviert.[1]
- Da P53 das proof-reading kontrolliert, wird durch Ausschalten von P53 die Fehlerkontrolle aufgegeben und Mutationen können sich anhäufen, die zur Ausschaltung weiterer Tumorsuppressor-Gene oder zur Aktivierung zellwachstums-fördernder Proteine führen können.
Andererseits ist in Tumorzellen die globale DNA-Methylierung geringer als in Normalzellen. Das führt man darauf zurück, dass das in Normalzellen hochmethylierte Heterochromatin (vor allem die Zentromer-Region) in Tumorzellen geringer methyliert ist.
Seitdem man den Einfluss der Hypermethylierung auf das Tumorwachstum identifiziert hat, hat man nach Wegen gesucht, um durch Demethylierung die im Entstehen begriffenen bzw. auch schon existierende Tumore wieder der Zellzykluskontrolle zu unterwerfen:[8]
- Cytosin-ähnliche Substanzen wie Azacytosin oder Aza-Desoxy-Cytosin werden in Patienten mit Akuter Myeloischer Leukämie infundiert.
- Diese Stoffe werden in Zellen aufgenommen, deren DNA verdoppelt wird.
- Azacytosin kann in der Zelle in Aza-Desoxy-Cytosin umgewandelt werden.
- Aza-Desoxy-Cytosin wird anstelle von Cytosin in DNA eingebaut.
- Die DNMT3, die die hemimethylierten CGs methylieren will, bindet an das Aza-Analog.
- Der Austausch von Kohlenstoff gegen Stickstoff bewirkt, dass das Enzym bei dem enzymatischen Methyltransfer an der DNA hängenbleibt und keine weiteren Reaktionen durchführen kann.
- Mit diesem Verfahren werden die DNMT3 inaktiviert und eliminiert. Eine Methylierung findet nicht mehr statt.
- Nach der nächsten Zellteilung ist die DNA weniger methyliert. Wenn von dieser De-Methylierung z.B. das P53- oder das P15-Gen betroffen sind, findet wieder Zellzykluskontrolle statt.
- Das Tumorwachstum ist damit unterbunden.
Es wurden klinische Studien veröffentlicht, in denen bei menschlichen Patienten ein hemmender Effekt von Aza-Desoxy-Cytosin auf Tumorentwicklung gezeigt werden konnte.[9] Die Forscher nennen ihr Verfahren Epigenetische Therapie.
Für die Behandlung des Myelo-Dysplastischen Syndroms, das sich häufig zu einer Akuten Myeloischen Leukämie entwickelt, wurde 5-Aza-2'-Desoxy-Cytosin unter dem Namen Dacogen von der FDA im Jahre 2006 als Medikament freigegeben Dacogen-Freigabe durch die FDA. Ein anderer Name für diese Substanz ist Decitabin.
Abgrenzung und Zusammenhang der Begriffe Methylierung, Modifikation, Mutation und Vererbung
Die Methylierung ist eine universelle chemische Abwandlung von Molekülen. Im Bereich anorganischer und organischer Chemie spricht man bei solchen Abwandlungen eher von Derivaten, im Bereich der biologischen Makromoleküle eher von Modifikationen. So können außer den Nukleobasen in der DNA auch Proteine durch Methyltransferasen methyliert werden. Der Begriff Modifikation wird in der Biologie mehrdeutig gebraucht. Zum einen betrifft er die bereits erwähnte Modifikation von Makromolekülen – vor allem DNA –, zum anderen die phänotypische Modifikation. Von (phänotypischer) Modifikation spricht man, wenn sich die Eigenschaften eines Lebewesens durch geänderte Umweltbedingungen ändern (geänderter Phänotyp), ohne dass die Erbsubstanz in der Abfolge ihrer Grundbausteine verändert wurde (unveränderter Genotyp). Somit sind die Modifikation von Makromolekülen und die phänotypische Modifikation verschieden. Allerdings haben beide Begriffe die Gemeinsamkeit, sich auf solche Veränderungen am Lebewesen zu beziehen, welche ohne grundsätzliche Änderung der Erbsubstanz vonstatten gingen. Keine grundsätzliche Änderung heißt in diesem Fall, dass keine Änderung der Abfolge der Grundbausteine der Desoxyribonukleinsäure erfolgte. Die DNA-Modifikation und die phänotypische Modifikation sind daher keine Mutationen. DNA-Modifikationen können Mutationen nach sich ziehen. So steigt die Wahrscheinlichkeit, dass ein Cytosin zum Thymin umgewandelt wird (Punktmutation), wenn dieses Cytosin methyliert ist (siehe CpG-Inseln).
DNA-Modifikationen können auch phänotypische Modifikationen nach sich ziehen: Veränderte Umweltbedingungen führen über die Signaltransduktion zum veränderten Methylierungsmuster der DNA in bestimmten Bereichen (DNA-Modifikation); dadurch wird die Nutzung der Gene verändert (differenzielle Genexpression); das führt zu einer Änderung der Eigenschaften der Lebewesens (phänotypische Modifikation). Mutationen sind per Definition vererbbar. DNA-Modifikationen sind nicht oder nur im begrenzten Maße vererbbar. Die genomische Prägung ist ein Grenzfall. Hier kommt es durch DNA-Methylierung zu einer Unterscheidung väterlicher und mütterlicher Allele. Da nur ein Allel (eine Gen-Kopie) aktiv ist, kommt es zur Weitergabe der Ausprägung dieses Allels in die nächste Generation. Es handelt sich hierbei jedoch nicht um genetische Vererbung (Biologie) im engeren Sinne.
Literatur
- Shen L, Waterland RA: Methods of DNA methylation analysis. In: Curr Opin Clin Nutr Metab Care. 10, Nr. 5, September 2007, S. 576–81. doi:10.1097/MCO.0b013e3282bf6f43. PMID 17693740.
- Beck S, Rakyan VK: The methylome: approaches for global DNA methylation profiling. In: Trends Genet.. 24, Nr. 5, Mai 2008, S. 231–7. doi:10.1016/j.tig.2008.01.006. PMID 18325624.
- Shames DS, Minna JD, Gazdar AF: DNA methylation in health, disease, and cancer. (PDF) In: Curr. Mol. Med.. 7, Nr. 1, Februar 2007, S. 85–102. PMID 17311535.
- Jowaed A, Schmitt I, Kaut O, Wüllner U: Methylation regulates alpha-synuclein expression and is decreased in Parkinson's disease patients' brains. In: J. Neurosci.. 30, Nr. 18, Mai 2010, S. 6355–9. doi:10.1523/JNEUROSCI.6119-09.2010. PMID 20445061.
Einzelnachweise
- ↑ a b Jeltsch, A. (2002): Beyond Watson and Crick: DNA methylation and molecular enzymology of DNA methyltransferases. In: ChemBioChem 3(4); 274–293; PMID 11933228
- ↑ Jeltsch, A. (2006): Molecular enzymology of mammalian DNA methyltransferases. In: Curr Top Microbiol Immunol. 301; 203–225; PMID 16570849
- ↑ Cervoni, N. et al. (1999): DNA demethylase is a processive enzyme. In: J. Biol Chem. 274(13); 8363–8366; PMID 10085064; PDF (freier Volltextzugriff, engl)
- ↑ Barras, F. und Marinus, MG. (1989): The great GATC: DNA methylation in E. coli. In: Trends Genet. 5(5); 139–143; PMID 2667217
- ↑ Marinus, MG. und Casadesus, J. (2009): Roles of DNA adenine methylation in host-pathogen interactions: mismatch repair, transcriptional regulation, and more. In: FEMS Microbiol Rev. 33(3); 488–503; PMID 19175412; doi:10.1111/j.1574-6976.2008.00159.x
- ↑ Mathieu, O. et al. (2002): Methylation of a euchromatin-heterochromatin transition region in Arabidopsis thaliana chromosome 5 left arm. In: Chromosome Research 10(6); 455–466; PMID 12489828; doi:10.1023/A:1020936229771
- ↑ Daura-Oller E, Cabre M, Montero MA, Paternain JL, Romeu A: Specific gene hypomethylation and cancer: new insights into coding region feature trends. In: Bioinformation. 3, Nr. 8, 2009, S. 340–3. PMID 19707296. Volltext bei PMC: 2720671.
- ↑ Momparler, RL. und Bovenzi, V. (2000): DNA methylation and cancer. In: J. Cell Physiol. 183(2); 145–154; PMID 10737890
- ↑ Wijermans, PV. et al. (2005): An epigenetic approach to the treatment of advanced MDS; the experience with the DNA demethylating agent 5-aza-2'-deoxycytidine (decitabine) in 177 patients. In: Ann. Hematol. 84(Suppl1); 9–17; PMID 16211386; doi:0.1007/s00277-005-0012-1
Kategorien:- Biologischer Prozess
- Genexpression
- Epigenetik

Wikimedia Foundation.