- Springschwänze
-
Springschwänze 
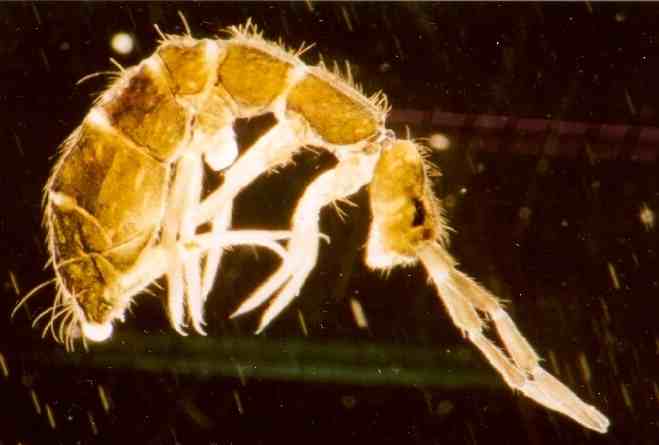
Springschwanz (Isotoma anglicana), etwa 40fach vergrößert. Unter dem Hinterleib sind Ventraltubus (zwischen den Beinen) und Furca sichtbar.
Systematik Überstamm: Urmünder (Protostomia) Überstamm: Häutungstiere (Ecdysozoa) Stamm: Gliederfüßer (Arthropoda) Unterstamm: Tracheentiere (Tracheata) Überklasse: Sechsfüßer (Hexapoda) Klasse: Springschwänze Wissenschaftlicher Name Collembola Lubbock, 1870 Ordnungen - Poduromorpha
- Entomobryomorpha
- Neelipleona
- Symphypleona
 In den Bodenporen lebende (euedaphische) Springschwänze sind wenig pigmentiert oder völlig weiß. Protaphorura armata findet sich häufig in vom Menschen beeinflussten Böden.
In den Bodenporen lebende (euedaphische) Springschwänze sind wenig pigmentiert oder völlig weiß. Protaphorura armata findet sich häufig in vom Menschen beeinflussten Böden.
 Orchesella flavescens gehört zur Familie der Entomobryidae, in der häufig artspezifische Färbungsmuster auftreten.
Orchesella flavescens gehört zur Familie der Entomobryidae, in der häufig artspezifische Färbungsmuster auftreten. Springschwänze (Anurida maritima, Neanuridae) auf küstennaher Wasseroberfläche.
Springschwänze (Anurida maritima, Neanuridae) auf küstennaher Wasseroberfläche.
Die Springschwänze (Collembola) sind eine zu den Sackkieflern (Entognatha) gehörende Klasse der Sechsfüßer (Hexapoda). Sie erreichen eine Körpergröße von 0,1 mm bis zu 17 mm und finden sich vor allem auf und in den Humusschichten nicht zu trockener Böden bis in einige Meter Tiefe und an verrottendem Pflanzenmaterial. Sie besiedeln dabei auch so unterschiedliche Habitate wie Regenwälder, küstennahe Uferbereiche[1], Sanddünen, Wüsten[2] oder Schneeflächen im Hochgebirge.[3][4]Inhaltsverzeichnis
Merkmale
Springschwänze zeigen die für die Hexapoda typische Dreiteilung des Körpers in Kopf, den beintragenden Thorax (Brustkorb) und das Abdomen (Hinterleib). Die meisten Arten sind zwischen 1 und 5 mm lang. Oberirdisch lebende (epedaphische) Arten sind eher dunkel pigmentiert, teilweise mit artspezifischen Färbungsmustern, starker Behaarung und großen Körperanhängen. Überwiegend oder ausschließlich im Boden lebende (euedaphische) Arten sind als Anpassung an ihre Lebensweise im Porengefüge des Bodens wenig gefärbt bis völlig weiß, von kompaktem Körperbau, mit kurzen Antennen, teilweise oder völlig reduzierter Sprunggabel und verringerter Augenzahl. Der Körper ist bei allen Arten nur schwach sklerotisiert, seine Oberfläche (Epicuticula) ist mit zahlreichen Mikrotuberkeln besetzt, die meist 0,3 Millimeter lang und wasserabweisend sind. Ein Tracheensystem besitzen nur einige Gruppen der Kugelspringer, die meisten Collembolen atmen ausschließlich über ihre dünnwandige Cuticula.
Insekten haben normalerweise elf Abdominalsegmente, die Collembola hingegen nur sechs, welche auch bereits während der Embryonalentwicklung angelegt werden. Collembolen werden zu den Entognatha gezählt, da ihre Mundwerkzeuge in einer Mundtasche liegen und äußerlich nicht sichtbar sind. Charakteristisch für die meisten dieser flügellosen Tiere ist ihre Sprunggabel (Furca, manchmal auch Furcula), die sich am vierten Abdominalsegment befindet. Sie besteht aus drei Teilen: dem basalen Manubrium, den langen paarigen Dentes und terminal an diesen je einer kurzen Hakenstruktur, dem Mucro. Zwischen Manubrium und Dentes befinden sich cutinisierte 'Zähne', die genau in die Haken des Retinaculums, einer Struktur am dritten Abdominalsegment, hineinpassen und so die Furca ventral am Abdomen unter Spannung festhalten. Bei einer Reizung des Tieres löst es diese Verbindung, die Mucrones bohren sich in den Untergrund und der Collembole vollführt einen ungerichteten Sprung aus der Gefahrenzone. Die Sprungdistanz beträgt ein Vielfaches seiner eigenen Körperlänge. Bei vielen im Boden lebenden Arten ist die Furca zurückgebildet; einige sondern stattdessen bei Gefahr Wehrsekrete ab, die abschreckend auf ihre Fressfeinde wirken. [5][6] Allen Arten gemein ist der hinter den Beinen gelegene Ventraltubus (Collophor), der sich am 1. Abdominalsegment befindet und vermutlich für den Wasser- und Elektrolythaushalt eine wichtige Rolle spielt. Mit Hilfe des Ventraltubus können sich Springschwänze auch an glatten Oberflächen festhalten und fortbewegen. Von dieser Struktur leitete der englische Naturforscher John Lubbock den wissenschaftlichen Namen Collembola ab, nach dem griechischen kolla, „Leim“, und embolon, „Keil, Zapfen“.[7]
Die Komplexaugen der Springschwänze bestehen aus maximal acht Ommatidien, die oft reduziert bzw. ganz zurückgebildet sind. Zwischen der Antennenbasis und dem Ommenfeld findet sich bei den meisten Arten das Postantennalorgan, ein Sinnesorgan in einer Hautvertiefung, das der Chemorezeption dient und den Rest einer zurückgebildeten zweiten Antenne darstellt.[8] Springschwänze erreichen üblicherweise ein Alter zwischen sechs und zwölf Monaten, der Altersrekord im Labor liegt bei fünfeinhalb Jahren.[9] In dieser Zeitspanne häuten sich die meisten Arten bis zu 50 Mal. Die Geschlechtsreife erreichen sie nach etwa fünf Häutungen. Ein Weibchen legt in seinem Leben zwischen 150 und 350 Eier, der schlüpfende Nachwuchs wird seinerseits nach etwa vier Wochen geschlechtsreif.
Lebensraum, Verbreitung und Häufigkeit
Springschwänze leben am und im Boden, meist bis in zehn Zentimeter Tiefe, aber auch auf Baumrinde, auf der Wasseroberfläche, an Meeresküsten, auf Gletschern und in Nestern von Ameisen und Termiten. Sie bevorzugen hohe Luftfeuchtigkeit, manche Arten werden durch Kohlenstoffdioxid angelockt.[10]
Durch ihre wasserabweisende Cuticula können sie Überflutungen des Bodenporensystems in einer Luftblase überstehen oder auf der Wasseroberfläche manövrieren. Einige Arten wie der Schwarze Wasserspringer weiden dort gezielt Algen, Bakterien und Einzeller ab. Springschwänze können bis zu 14 Tage auf offener See treibend überleben und dabei mehrere hundert Kilometer zurücklegen.[11] Sie erlangen dadurch als Erstbesiedler neuer, steriler Landstriche Bedeutung (s. auch Surtsey). Arten wie Schnee- und Gletscherfloh leben im Lückensystem von Geröll, in Moospolstern und Felsritzen des Hochgebirges und ernähren sich von den auf die Eisflächen gewehten Koniferenpollen. Diese Arten sind noch bei Temperaturen um -5 °C aktiv. Arktische Collembolen können wenigstens vier Jahre bei -20 °C eingefroren überleben.[12]
Von mehreren Arten ist bekannt, dass sie Schwermetalle aus dem Boden aufnehmen und immobilisieren können. Unter anderem diese Fähigkeiten machen Collembolen zu wichtigen Erst- und Wiederbesiedlern gestörter oder kontaminierter Böden, etwa von überfluteten Auenböden[13] oder Abraumhalden.[14]
Die ältesten bekannten Fossilfunde von Collembolen sind etwa 400 Millionen Jahre alt. Springschwänze gehören damit zu den ältesten landlebenden Tieren überhaupt, was auch ihre weltweite Verbreitung in fast allen terrestrischen Habitaten erklärt. Dabei bevorzugen sie kühle, feuchte Umgebungsbedingungen.
Springschwänze gelten als die häufigsten Sechsfüßer. In einem Quadratmeter Boden leben bis in eine Tiefe von 30 Zentimetern bis zu 400.000 Individuen. Nach den Milben sind sie damit die individuenreichste Tiergruppe der Mesofauna im Boden. Ihre Häufigkeit orientiert sich an Faktoren wie Lichtverhältnissen, Feuchtigkeit, Humusform, pH-Wert des Bodens und Nährstoffverfügbarkeit. Dementsprechend treten sie nicht gleichmäßig verteilt auf, sondern eher konzentriert in „Hot spot“-Mustern überall dort, wo sie optimale Lebensbedingungen vorfinden. Hier können sie kurzfristig Massenbestände aufbauen und etwa an warmen Wintertagen oder im zeitigen Frühjahr ein auffälliges Schwarmverhalten zeigen.[15]
Nahrung
Die meisten Arten der Springschwänze sind Detritusfresser, sie ernähren sich von zerfallenden pflanzlichen Stoffen, Exkrementen oder Aas. Es gibt neben diesen ‚Allesfressern‘ aber auch Spezialisten, die nur Algen, Pilze und Pollen fressen oder Mikroorganismen abweiden.
Ökologische und wirtschaftliche Bedeutung
 Der Luzernefloh (Sminthurus viridis) gehört zu den Kugelspringern.
Der Luzernefloh (Sminthurus viridis) gehört zu den Kugelspringern.Durch den Abbau ihrer Nahrung sind sie wesentlich an der Bildung von Humus beteiligt. Sie beseitigen dabei organische Rückstände und fördern so die Bodenfruchtbarkeit und damit das Wachstum von Pflanzen. Da sie die Reste von Pflanzen in natürlichen Dünger verwandeln, sind sie der Landwirtschaft von erheblichem Nutzen.
Nur wenige Arten, wie z. B. der Luzernefloh (Sminthurus viridis), gelten als Schädlinge für Agrarsysteme.[16] Springschwänze können gelegentlich für Monokulturen im Freiland ebenso wie für Zimmerpflanzen schädlich werden, wenn ihre eigentliche Nahrungsquelle, pflanzlicher Detritus, zur Neige geht und sie die lebenden Feinwurzeln anfressen. Durch gezieltes Abweiden von Pilzmyzelien verringern sie andererseits die Gefahr von Pilzbefall bei Samen und Keimlingen und tragen so zum Pflanzenschutz in Agrarökosystemen bei.[17] Einige Springschwanzarten reagieren empfindlich auf anthropogene Störungen im Boden und werden daher im Labor bei Standardtests zum Nachweis von Bodenkontamination eingesetzt. Insbesondere der im Labor leicht zu haltende Blumentopfspringschwanz Folsomia candida (Isotomidae) gibt als Testorganismus durch Änderungen seines Fraß- und Fortpflanzungsverhaltens oder bei Vermeidungsexperimenten Hinweise auf vorhandene Stör- und Schadsubstanzen.[18]
Der Mensch hat nur wenigen Springschwanzarten einen Trivialnamen gegeben. Zu diesen gehören neben einigen Agrarschädlingen (Beispiel Luzernefloh) vor allem Arten mit auffälligem Massenauftreten zu bestimmten Jahreszeiten (Schneefloh) oder in bestimmten Habitaten (Gletscherfloh, Schwarzer Wasserspringer, Blumentopfspringschwanz).Systematik
 Isotomurus palustris (Isotomidae) ist eine häufige Art in feuchten Habitaten.
Isotomurus palustris (Isotomidae) ist eine häufige Art in feuchten Habitaten.Die Springschwänze (Collembola) wurden traditionell zu den Insekten gerechnet und hier mit anderen flügellosen Taxa als „Urinsekten“ (Apterygota) an die Basis der Insecta gestellt. Mittlerweile werden die Apterygota als paraphyletisch und die Springschwänze als eigene Klasse innerhalb der Sechsfüßer (Hexapoda) angesehen. Da sie mehrere gemeinsame Merkmale aufweisen, gelten die Beintastler (Protura) als ihre Schwestergruppe. Nach einem dieser Merkmale, dem Fehlen von Hinterleibsanhängen, werden beide Gruppen zusammen als Ellipura (griechisch έλλειψις, „Fehlen“, und ουρά, „Schwanz“) bezeichnet. Diese Einteilung geht auf Willi Hennig zurück,[19] ebenso wie die Zuordnung der Doppelschwänze als Schwestergruppe der Ellipura. Protura, Collembola und Diplura werden als Klasse der Entognatha zusammengefasst, weil nur bei diesen drei Gruppen die Mundwerkzeuge sich in der Kopfkapsel befinden statt außerhalb wie bei den meisten Insekten. Neue Fossilfunde und molekulargenetische Untersuchungen stellen diese traditionelle Sicht in Frage. Auf der Basis mitochondrialer Gensequenzen wird eine engere Verwandtschaft der Collembola mit den Kiemenfußkrebsen [20] oder den Kieferfußkrebsen[21] postuliert. Nach diesen Modellen hätte sich der segmentierte Körperbau der Hexapoda mehrfach unabhängig voneinander entwickelt. Untersuchungen ribosomaler Proteinsequenzen stützen hingegen eine engere Verwandtschaft der Collembola mit den geflügelten Insekten, gefolgt von den Kiemenfußkrebsen und den Höheren Krebsen als nächstverwandten Taxa, was mit den morphologisch basierten Stammbäumen eher übereinstimmen würde.[22] Eine neuere Untersuchung auf der Basis ribosomaler Sequenzen, einer vergrößerten Anzahl untersuchter Taxa und einer optimierten Merkmalsgewichtung stützt teilweise die Stellung der Collembola als monophyletisches Taxon innerhalb der Hexapoda und stellt ihnen die Protura und Diplura als gemeinsame Schwestergruppe („Nonoculata“) zur Seite, ohne aber diese Aufteilung in allen untersuchten Rechenmodellen bestätigen zu können.[23] Bis zur Einbeziehung weiterer RNA-unabhängiger Merkmale in die Stammbaumanalyse bleibt die tatsächliche Stellung der Collembola innerhalb der Gliederfüßer unklar.
Ursprünglich unterteilte man die Collembola in die langgestreckten Arthropleona und die eher rundlichen Symphypleona (Kugelspringer) mit ihren typischen verschmolzenen Abdominalsegmenten.[24] Die neuere Systematik ersetzt die Unterklasse Arthropleona durch Poduromorpha und Entomobryomorpha, die als eigene Ordnungen neben die Neelipleona und Symphypleona gestellt werden.[25] Andere Systematiker sehen die Neelipleona als den Sminthuroidea zugehörig. Die Proportionen der Thorakal- und Abdominalsegmente werden zur Unterteilung der Großgruppen nach wie vor verwendet: bei den Entomobryomorpha ist das Tergit des ersten Thoraxsegments verkürzt, verschwindet weitgehend zwischen Kopf und zweitem Thoraxsegment und trägt keine Borstenhaare (Chaetae bzw. Setae), bei den Poduromorpha ist es normal entwickelt und trägt mindestens eine Reihe von Chaetae. Innerhalb der Entomobryomorpha ist bei den Entomobryidae das vierte Abdominaltergit deutlich länger als das dritte, bei Isotomidae hingegen sind beide etwa gleich lang. Bei den Symphypleona wird die kugelige Hinterleibsstruktur hauptsächlich von den Abdominalsegmenten gebildet, bei den Neelipleona (bzw. Neelida) hauptsächlich von den Thoraxsegmenten.
Die Zahl tatsächlich auf dem Planeten vorhandener Collembolenarten wurde auf 50.000 geschätzt[26], weltweit wurden bislang 8000 Arten beschrieben.[27] Angaben über die Zahl beschriebener Arten schwanken stark, was nicht zuletzt damit zusammenhängt, dass viele Arten und Artengruppen sehr schwer zu bestimmen und auseinanderzuhalten sind. Komplexe Genitalstrukturen, wie sie bei vielen anderen Hexapoda zur Taxierung herangezogen werden, sind bei den Collembola nicht vorhanden. Bei Arten einiger Collembolengattungen (z. B. Isotomurus, Orchesella) kann das Färbungsmuster für die Artidentifikation verwendet werden, während bereits bei nahe verwandten Gattungen die entsprechenden Muster sehr stark variieren. Bei den paläarktisch weitverbreiteten Familien der Hypogastruridae und Isotomidae werden Ökomorphosen beobachtet, d. h. ungünstige Umweltbedingungen, besonders Dürre und Hitze, können die morphologische Gestaltung der Körperanhänge, die Beborstung einzelner Körpersegmente und der Mundwerkzeuge beeinflussen.[28][29] Bei Isotomiden, die kalte Klimata bewohnen, treten solche Ökomorphosen als Cyclomorphosen regelmäßig auf, Winter- und Sommerformen unterscheiden sich in Form und Gestalt, v. a. der Furca und der Beine. Da Ausprägung und Proportionen derartiger Merkmale für die Artdiagnose häufig herangezogen werden, bleibt die Zahl tatsächlich vorhandener Arten mit Unsicherheiten behaftet. Beispielsweise wurde die Art Mesaphorura krausbaueri Börner, 1901 noch 1960 als gut abgegrenzte Art aufgefasst, danach als Pulk schwer unterscheidbarer Varietäten oder kryptischer Arten erkannt und zunächst behelfsweise als „Tullbergia krausbaueri-Gruppe“ bezeichnet. Genauere methodische Ansätze[30][31] unterteilen mittlerweile diese Gruppe allein in der Paläarktis in acht Genera und 43 valide Arten. Der ähnlich problematische Protaphorura armata-Komplex umfasst je nach Autor eine[32], dreizehn[33] oder bis zu 44 Arten nebst diversen Unterarten.[34]
Nach wie vor werden sowohl neue Arten beschrieben als auch bei bereits beschriebenen der Artstatus durch Anwendung genauerer Methoden exakter festgelegt, um aus dem bislang oft ungenau definierten Übergangsbereich zwischen Artenkomplex, Art, Farbmorphe, Unterart und Varietät valide biologische Arten herauszuschälen. Die Einführung neuer taxonomischer Merkmale erleichtert die hierbei notwendige Unterscheidung zwischen inner- und zwischenartlicher Variabilität. Während Färbungsmuster aufgrund ihrer hohen Variabilität an Bedeutung verlieren, wird die Chaetotaxie, das ist das Vorhandensein oder Fehlen bestimmter Borsten auf definierten Positionen eines Körpersegments, immer häufiger erfolgreich zur Artabgrenzung herangezogen; die Chaetotaxie der ersten Häutungsstadien auch zur Klärung der Verwandtschaftsbeziehungen der höheren Collembolentaxa.[35] Die Feinstruktur der Mundwerkzeuge liefert ebenfalls wichtige Bestimmungsmerkmale, erfordert allerdings sorgfältiges Präparieren. Molekularbiologische Methoden werden seit den 1970er Jahren erfolgreich zur Auflösung kryptischer Artenkomplexe eingesetzt, sie führten aber auch zu einem anhaltenden Disput um die tatsächliche Stellung der Collembola innerhalb oder außerhalb der Hexapoda.[36][37]
Siehe auch
Quellen
Einzelnachweise
- ↑ McMeechan, F.K.; Manica, A.; Foster, W.A. (2000). Rhythms of activity and foraging in the intertidal insect Anurida maritima: Coping with the tide. Journal of the Marine Biological Association of the United Kingdom, 80, 189–190.
- ↑ Suhardjono, Y.R.; Greenslade, P. (1994). Folsomides arnoldi n.sp. (Isotomidae): A new Collembolan abundant in arid Australia, with a redescription of Folsomides denisi (Womersley). Proceedings of the Linnean Society of New South Wales, 114, 21–27.
- ↑ Yosii, R. (1966). Snow Collembola of the Siachen Glacier in Karakoram. Results of the Kyoto University Scientific Expedition to the Karakoram and Hindukush, 8, 407–410.
- ↑ Hemmer, W. (2000). Onychiurus (Protaphorura) kolenatii n.sp. (Collembola), eine fakultativ rotgefärbte Onychuridenart von hochalpinen Schneefeldern der Ostalpen. Vorarlberger Naturschau, 8, 145–152. http://www.landesmuseum.at/pdf_frei_remote/VNFE_8_0145-0152.pdf
- ↑ Dettner, K., Scheuerlein, A., Fabian, P., Schulz, S. & Francke, W. (1996). Chemical defense of giant springtail Tetrodontophora bielanensis (Waga) (Insecta: Collembola). Journal of Chemical Ecology, 22, 1051-1074.
- ↑ Messner, C., Walther, J., Dettner, K. & Schulz, S. (2000). Chemical deterrents in podurid Collembola. Pedobiologia, 44, 210-220.
- ↑ Lubbock, J. (1873). Monograph of the Collembola and Thysanura. London: The Ray Society.
- ↑ Lawrence, P. N. (1999). From whence and whither the Collembola? Crustaceana, 72, 1110-1122.
- ↑ Hopkin, S. P. (2002). Collembola. In: Lal, R., Encyclopedia of Soil Science (pp. 207-210). New York: Marcel Dekker Inc.
- ↑ Dunger, W. & Fiedler, H.J. (1989). Methoden der Bodenbiologie. Jena: Gustav Fischer Verlag.
- ↑ Coulson, S. J., Hodkinson, I. D., Webb, N. R. & Harrison, J. A. (2002). Survival of terrestrial soil-dwelling arthropods on and in seawater: implications for trans-oceanic dispersal. Functional Ecology, 16 (3): 353-356.
- ↑ Coulson, S. J. & Birkemoe, T. (2000). Long-term cold tolerance in Arctic invertebrates: recovery after 4 years at below -20%C. Canadian Journal of Zoology, 78 (11): 2055-2058.
- ↑ Russell, D., Hauth, A. & Fox, O. (2004). Community dynamics of soil Collembola in floodplains of the Upper Rhine Valley. Pedobiologia, 48 (5-6): 527-536.
- ↑ Rusek, J. (2004). Collembola succession on deposits from a chemical factory. Pedobiologia, 48 (5-6): 519-524.
- ↑ Zettel, J. & Zettel, U. (2008). Manche mögen's kalt: die Biologie des «Schneeflohs» Ceratophysella sigillata (Uzel, 1891), einer winteraktiven Springschwanzart (Collembola: Hypogastruridae). Mitteilungen der Naturforschenden Gesellschaft in Bern, 65, 79-110.
- ↑ Bishop, A. L., Harris, A. M. & McKenzie, H. J. (2001). Distribution and ecology of the lucerne flea, Sminthurus viridis (L.) (Collembola: Sminthuridae), in irrigated lucerne in the Hunter dairying region of New South Wales. Australian Journal of Entomology, 40, 49-55.
- ↑ Sabatini, M. A. & Innocenti, G. (2001). Effects of Collembola on plant-pathogenic fungus interactions in simple experimental systems. Biology and Fertility of Soils, 33, 62-66.
- ↑ Fountain, M. T. & Hopkin, S. P. (2005). Folsomia candida (Collembola): A "Standard" Soil Arthropod. Annual Review of Entomology, 50, 201-222.
- ↑ Hennig, W. (1969). Die Stammesgeschichte der Insekten. Waldemar Kramer, Frankfurt am Main.
- ↑ Nardi, F. Spinsanti, G., Boore, J. L., Carapelli, A., Dallai, R., Frati, F. (2003). Hexapod Origins: Monophyletic or Paraphyletic? Science, 299(5614), 1887-1889.
- ↑ Cook, C.E., Yue, Q., Akam, M. (2005). Mitochondrial genomes suggest that hexapods and crustaceans are mutually paraphyletic. Proceedings of the royal Society of London Series B, 272, 1295-1304
- ↑ Timmermans, M. J. T. N., Roelofs, D., Mariën, J., van Straalen, N. M. (2008). Revealing pancrustacean relationships: Phylogenetic analysis of ribosomal protein genes places Collembola (springtails) in a monophyletic Hexapoda and reinforces the discrepancy between mitochondrial and nuclear DNA markers. BMC Evolutionary Biology, 8 (83).
- ↑ von Reumont, B. M., Meusemann, K., Szucsich, N. U., Dell'Ampio, E., Gowri-Shankar, V., Bartel, D., Simon, S., Letsch, H. O., Stocsits, R. R., Luan, Y. X., Wägele, J. W., Pass, G., Hadrys, H. & Misof, B. (2009). Can comprehensive background knowledge be incorporated into substitution models to improve phylogenetic analyses? A case study on major arthropod relationships. BMC Evolutionary Biology, 9 (119).
- ↑ Börner, C. (1901). Zur Kenntnis der Apterygoten-Fauna von Bremen und der Nachbardistrikte. Abhandlungen des Naturwissenschaftlichen Vereins zu Bremen, 17, 1-141.
- ↑ Deharveng, L. (2004). Recent advances in Collembola systematics. Pedobiologia, 48, 415-433.
- ↑ Hopkin, S. P. (1997). Biology of the Springtails (Insecta: Collembola). Oxford: Oxford University Press.
- ↑ http://www.collembola.org/, Version 1.iv. 2010, Abruf 15.iv.2010.
- ↑ Cassagnau, P. (1971). Les différents types d'ecomorphose chez les Collemboles Isotomidae. Revue d'Ecologie et de Biologie du Sol, 8, 55-57.
- ↑ Bourgeois, A. (1973). Polymorphisme et épitoquie chez Ceratophysella tuberculata (Collembole, Hypogastruridae), Revue d'Ecologie et de Biologie du Sol, 10, 589-601.
- ↑ Rusek, J. (1971). Zur Taxonomie der Tullbergia (Mesaphorura) krausbaueri (Börner) und ihrer Verwandten (Collembola). Acta Entomologica Bohemoslovaca, 68, 188-206.
- ↑ Dunger, W. (1991). Ökologische Prüfung von Morphospecies der „Tullbergia krausbaueri-Gruppe“ (Insecta: Collembola). Mitteilungen aus dem Museum für Naturkunde in Berlin, 67 (1): 131-140.
- ↑ Bödvarsson, H. (1970). Studies of Onychiurus armatus (Tullberg) and Folsomia quadrioculata (Tullberg) (Collembola). Opuscula Entomologica, Supplementum, 182 pp.
- ↑ Pomorski, R. (1990). Morphological-systematic studies on the variability of pseudocelli and some morphological characters in Onychiurus of the „armatus-group“ (Collembola, Onychiuridae) Part II. On synonyms within the armatus-group, with special reference to diagnostic characters. Annales Zoologici, 43, 535-575.
- ↑ Gisin, H. (1960). Collembolenfauna Europas. Genf, Musée D'Histoire Naturelle, 311 pp.
- ↑ Deharveng, L. (2004). Recent advances in Collembola systematics. Pedobiologia, 48, 415-433.
- ↑ Nardi, F. Spinsanti, G., Boore, J. L., Carapelli, A., Dallai, R., Frati, F. (2003). Hexapod Origins: Monophyletic or Paraphyletic? Science, 299(5614), 1887-1889.
- ↑ Delsuc, F., Phillips, M.J., Penny, D. (2003). Comment on "Hexapod Origins: Monophyletic or Paraphyletic?", Science, 301(5639), 1482.
Literatur
- Dunger, W. (1983). Tiere im Boden. Westarp Wissenschaften, 4. Aufl. 2008. ISBN 3-89432-424-4
- Dunger, W. (1996). Ordnung Collembola, Springschwänze. In: Westheide, W., Rieger, R., Spezielle Zoologie. Teil 1. Einzeller und wirbellose Tiere (pp. 71-86). Stuttgart u.a.: Gustav Fischer Verlag. ISBN 3-437-20515-3
- Hopkin, S. P. (1997). Biology of the Springtails (Insecta: Collembola). Oxford: Oxford University Press. ISBN 0-19-854084-1
Weblinks
-
 Commons: Springschwänze – Album mit Bildern und/oder Videos und Audiodateien
Commons: Springschwänze – Album mit Bildern und/oder Videos und Audiodateien - http://www.collembola.org :"To keep things simple, let's suppose the taxonomical speciation rate remains stable at the last 10-year speciation rate. Taking into account that the estimated number of Collembola species is about 50,000 (Hopkin 1998:118), collembolists will continue to describe new taxa for about 595 years..." - wissenschaftliche, sehr umfangreiche Seite über Springschwänze
- Pressebericht zu den Vorfahren der Springschwänze





Wikimedia Foundation.