- Hodgkin-Huxley-Modell
-
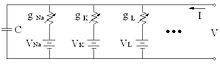 Elektrischer Schaltplan für das Hodgkin-Huxley-Modell
Elektrischer Schaltplan für das Hodgkin-Huxley-Modell
Das Hodgkin-Huxley-Modell ist das berühmteste Modell zur Simulation von Neuronen. Es wurde 1952 von Alan Lloyd Hodgkin und Andrew Fielding Huxley, ursprünglich zur Beschreibung der Entstehung von Aktionspotentialen im Riesenaxon des Tintenfisches, entwickelt. Beide Wissenschaftler erhielten zusammen mit Sir John Carew Eccles 1963 den Nobelpreis für Medizin "für ihre Entdeckungen über den Ionen-Mechanismus, der sich bei der Erregung und Hemmung in den peripheren und zentralen Bereichen der Nervenzellenmembran abspielt".
Das Modell simuliert Neuronen nahe an den biologischen Gegebenheiten, z. B. werden einzelne Aktionspotentiale (Spikes) modelliert. Dieses Modell wird auch häufig als Grundmodell, Elektrisches Modell oder Kabelmodell bezeichnet. Das Hodgkin-Huxley-Modell ist allerdings zur Modellierung großer Netzwerke nur schlecht geeignet, da es zu aufwendig ist. Stattdessen wird dann ein einfacheres Modell gewählt, zum Beispiel das FitzHugh-Nagumo-Modell.
Bei diesem Modell werden die spannungsabhängigen Membranwiderstände für die Ionenströme innerhalb einer Membran durch variable Widerstände modelliert. Die elektrische Membrankapazität C wird durch einen Kondensator simuliert. Der Gesamtmembranstrom ist die Summe der Kalium-, Natrium- und Leck- Ströme IK, INa und IL und des externen Stroms Iext (z. B. ein durch eine externe Elektrode applizierter Strom), der die Membran auflädt. Das Membranpotential V gehorcht dann der Differentialgleichung
 .
.
Die Ströme IK, INa und IL ergeben sich aus der Differenz der Membranspannung zum jeweiligen Gleichgewichtspotential E und der dazugehörigen Leitfähigkeit g:
 ,
, und
und .
.
Als Absolutwerte für die Umkehrpotentiale wurden gefunden EK=-77mV, ENa=50mV und EL=-54mV. Per Definition ist ein externer Strom (z. B. durch eine Elektrode) als positiver Einwärtsstrom definiert, während ein Membranstrom als positiver Auswärtsstrom gilt, dies erklärt die Wahl der Vorzeichen.
Die Leitfähigkeiten gK und gNa sind zeitabhängig und somit für die Entstehung eines Aktionspotentiales verantwortlich, während die Leck-leitfähigkeit gL konstant bleibt, da sich der Widerstand der Membran als unveränderlich annehmen lässt. Hodgkins und Huxleys geniale Idee war das Postulat von Gatingvariablen (engl. Gate: Tor), die die (probabilistischen) Dynamiken der Ionenkanäle nachbilden. Diese Variablen geben den Anteil der gerade geöffneten Kanäle an. Experimentelle Befunde ließen die Forscher auf drei Gatingvariablen schließen, die sie n, m und h nannten. Zusammen mit der Maximalleitfähigkeit
 eines Ions beschreibt die Gatingvariable den gerade durch die Membran fließenden Strom:
eines Ions beschreibt die Gatingvariable den gerade durch die Membran fließenden Strom: und
und
Die drei Gatingvariablen sind folgenden Dynamiken unterworfen:
 ,
, ,
, .
.
Die Funktionen αn(V),αm(V),αh(V),βn(V),βm(V),βh(V) sind jeweils exponentielle Funktionen, deren Parameter wiederum durch Experimente justiert wurden. Das sich ergebende gekoppelte, nichtlineare Differentialgleichungssystem lässt sich im Allgemeinen nicht analytisch sondern nur durch numerische Näherungsverfahren lösen.
Die postulierten Gatingvariablen haben sich später als echte Struktureigenschaft der spannungsabhängigen Ionenkanäle herausgestellt. Der Natriumkanal hat drei verschiedene Zustände, von denen nur einer Natriumionen hindurchlässt. Er unterliegt einer Aktivierung, die mit der Gatingvariablen m beschrieben wird und einer zeitlich versetzten Inaktivierung, die durch h beschrieben wird. Der Kanal kann sich in drei Zuständen befinden: geschlossen (inaktiv), geschlossen (aktiv), offen (aktiv).
Literatur
- A. L. Hodgkin, A. F. Huxley: A Quantitative Description of Membrane Current and its Application to Conduction and Excitation in Nerve. In: Journal of Physiology. 117, 1952, S. 500–544 (PMID 12991237).
Weblinks
Wikimedia Foundation.