- Umkehrpotential
-

Dieser Artikel wurde aufgrund von formalen und/oder inhaltlichen Mängeln in der Qualitätssicherung Biologie zur Verbesserung eingetragen. Dies geschieht, um die Qualität der Biologie-Artikel auf ein akzeptables Niveau zu bringen. Bitte hilf mit, diesen Artikel zu verbessern! Artikel, die nicht signifikant verbessert werden, können gegebenenfalls gelöscht werden.
Lies dazu auch die näheren Informationen in den Mindestanforderungen an Kurzartikel der Biologie.
Ein Membranpotential ist in der Zellbiologie die elektrische Spannung, die zwischen der Innen- und Außenseite einer Biomembran anliegt. Man spricht deshalb auch von einem Transmembranpotential.
Ein Membranpotential (genauer eine Membranspannung = Differenz zwischen elektrischen Potentialen) kommt dann zustande, wenn Innen- und Außenseite unterschiedliche Konzentrationen geladener Teilchen (Kation und Anion) aufweisen und die trennende Membran eine selektive Leitfähigkeit für eine oder mehrere Ionensorten besitzt. Ist beispiels- und typischerweise K+ innen höher konzentriert als außen, diffundiert K+ unter Zurücklassen von negativen Ladungen so lange vorzugsweise nach außen, bis sich für K+ beim Erreichen des Kalium-Gleichgewichtspotentials gemäß der Nernst-Gleichung Diffusion nach außen und elektrostatische Anziehung nach innen gerade die Waage halten. Bei tierischen Zellen ist das Membranpotential meist eine Mischung von Gleichgewichtspotentialen verschiedener Ionensorten (hauptsächlich K+, Na+ und Cl-), wobei K+ besonders ins Gewicht fällt, sodass in der Regel das Membranpotential innen negativ ist. Das Membranpotential von intakten Pflanzenzellen und Pilzen (wahrscheinlich auch von Prokaryoten) ist wegen H+-exportierender ATPasen meist noch negativer als das Diffusionspotential für K+, dem negativsten Diffusions-Gleichgewichtspotential der meisten Zellen. Wenn sich das Membranpotential über längere Zeit nicht wesentlich ändert, spricht man vom Ruhepotential, im Gegensatz zum dynamischen Aktionspotential, das bei Pflanzen und Pilzen vermutlich osmotische Aufgaben erfüllt (s. Osmoregulation) und bei Tieren (sowie wenigen Gefäßpflanzen) schnelle Signalübertragung durch den Organismus ermöglicht.
Membranpotentiale sind bei den meisten Zelltypen und bei allen Organismen zu finden, da die Kompartimentierung (Abgrenzung von der Umwelt und Unterteilung der Zelle in voneinander abgegrenzte Reaktionsräume durch semipermeable Membranen) eine Aufrechtherhaltung und Beeinflussung von Stoffgradienten ermöglicht. So kann auch über die Membran von Zellorganellen (Mitochondrium, Chloroplast, Vakuole) ein Membranpotential gemessen werden. Dort, wie in Prokaryoten, vermittelt das Membranpotential die Konvertierung zwischen physikalischer Energie (z.B. Licht und elektrochemische Gradienten) und metabolischer Energie (typischerweise ATP), so bei Photosynthese und zellulärer Atmung.
Inhaltsverzeichnis
Bedeutung
Das Membranpotential steuert Ionenströme
Ursache für die Diffusion der Ionen entlang eines Konzentrationsgefälles ist die Brownsche Molekularbewegung. Das Membranpotential kann diese Diffusion unterstützen, wenn Konzentrations- und Potentialgefälle in dieselbe Richtung wirken. Beispiel: Ausstrom von Kalium-Ionen aus einer Nervenzelle heraus, nachdem diese depolarisiert wurde und dabei ein positives Membranpotential erhielt (siehe Aktionspotential).
Wenn das Potentialgefälle dem Konzentrationsgefälle entgegengerichtet ist, können die Ionen nur dann passiv entsprechend dem Konzentrationsgefälle diffundieren, wenn die Wirkung des Konzentrationsgefälles größer als die entgegengesetzte Wirkung des Membranpotentials ist.
Es ist aber auch möglich, dass die Ionen gegen das Konzentrationsgefälle, aber mit dem Potentialgefälle diffundieren, wenn die Wirkung des Potentialgefälles größer als die entgegensetzte Wirkung des Konzentrationsgefälles ist. Beispiel: Einstrom von Kalium-Ionen aus dem Apoplasten ins Innere einer Schließzelle, nachdem die Zellmembran hyperpolarisiert wurde.
Strömen nun Ionen entsprechend dem elektrochemischen Potential von einer Seite zur anderen, werden deren Konzentrationsverhältnisse geändert, wodurch sich für eine Zelle verschiedene Auswirkungen ergeben:
- Stoffgradienten können als Energiespeicher dienen: Diffundieren Ionen „freiwillig“ (ΔG < 0, siehe unten) von einer Seite einer Biomembran zur anderen, wird Energie frei, die entweder zur Synthese von ATP (Chemiosmotische Kopplung) oder zum Transport anderer Stoffe genutzt wird.
- Die Änderung der Konzentrationsverhältnisse von Ionen führt auch zur Änderung des osmotischen Potentials eines Kompartiments: Auf die Seite der höheren Ionenkonzentration strömt mit Hilfe von Aquaporinen vermehrt Wasser ein.
- Durch die Volumenzunahme werden embryonale Zellen vergrößert (Wachstum).
- Durch die Änderung des osmotischen Drucks können bei Pflanzen Bewegungen wie das Öffnen und Schließen der Spaltöffnungen oder die Blattbewegungen bei der Mimose durchgeführt werden.
- Durch einströmendes Wasser wird das Cytosol verdünnt und damit werden die Konzentrationen aller Stoffe erniedrigt. So kann für bestimmte Stoffe ein Konzentrationsgefälle entstehen, die nun ebenfalls einströmen können (Assimilat-Transport bei Pflanzen).
Potentialänderungen als Signale
Bei Tier- und Pflanzenzellen, die auf Informationsverarbeitung und –weiterleitung spezialisiert sind, wird das Membranpotential im unerregten Zustand konstant gehalten (Ruhepotential). Bei Erregung ändert sich das Membranpotential kurzzeitig (Aktionspotential) durch Änderung der Permeabilität der Zellmembran für bestimmte Ionen. Dieses Signal kann sich über die Zelle ausbreiten, verursacht in Synapsen die Ausschüttung von Transmitterstoffen und löst in Muskelzellen eine Kontraktion aus.
Änderungen des Membranpotentials
Die Beeinflussung eines Membranpotentials erfolgt dadurch, dass der Transport von Ionen durch die Membran nur über passive Kanäle möglich ist, die von Transmembranproteinen gebildet werden. Diese Kanäle sind stoffselektiv, sie lassen also nur bestimmte Ionen passieren, und teilweise auch richtungsselektiv (Gleichrichterkanäle), lassen also die Ionen nur in einer Richtung durch die Membran diffundieren. Manche dieser Kanäle können aufgrund chemischer Signale (ligandengesteuerte Kanäle) oder durch Potentialänderungen (spannungsgesteuerte Kanäle) geöffnet oder geschlossen werden. Bestimmte Kaliumkanäle sind sowohl chemisch als auch potentialgesteuert.
Die Änderung eines Membranpotentials wird durch die Änderung der Permeabilität der Membran für bestimmte Ionen (in der Regel, Kalium-, Natrium- oder Calcium-Kationen oder Chlorid-Anionen) oder durch aktive Transportproteine wie ATPasen hervorgerufen.
Dabei gibt es prinzipiell zwei Möglichkeiten:
Depolarisation und Hyperpolarisation
- Bei einer Depolarisation wird das Membranpotential erniedrigt, da Kationen auf die Seite der Membran mit dem negativen Potential oder Anionen auf die Seite mit dem positiven Potential strömen.
- Bei einer Hyperpolarisation wird das Membranpotential erhöht, da Kationen auf die Seite der Membran mit dem positiven Potential oder Anionen auf die Seite mit dem negativen Potential strömen.
Diffusionsgleichgewicht
Werden die Ionenkanäle nicht rechtzeitig geschlossen, kommt es zum Diffusionsgleichgewicht, da sich durch die Diffusion die Konzentrations- und Potentialverhältnisse ändern. Dieses Gleichgewicht kann dann erreicht sein, wenn innen und außen die gleichen Ionenkonzentrationen vorliegen und das Membranpotential bei 0 mV liegt. Es sind aber auch Gleichgewichtslagen möglich, bei welchen das Membranpotential von Null verschieden ist und zwischen Innen und Außen verschiedene Konzentrationsverhältnisse vorliegen (siehe Ruhepotential).
Repolarisation
Die Rückkehr des Membranpotentials in den Ausgangszustand (bei Nervenzellen das Ruhepotential) wird Repolarisation genannt. Wann die Repolarisation eintritt und wie schnell diese abläuft, ist von der Funktion der jeweiligen Zelltypen abhängig. Bei Nervenzellen der Metazoa dauern Depolarisations- und Repolarisationsphase eines Aktionspotentials jeweils 2 ms, bei bestimmten Zellen der Pflanzen, die ein Aktionspotential erzeugen können, können beide Phasen mehrere Sekunden betragen.
Bei Nervenzellen erfolgt die Repolarisationsphase unmittelbar nach der Depolarisationsphase, beim Aktionspotential durch Öffnung spannungsgesteuerter Kalium-Kanäle innerhalb von 2 ms, bei graduierten Potentialen innerhalb von 40 ms bis 4000 ms durch Ionenpumpen, welche die eingeströmten Ionen wieder zurückpumpen.
Graduiertes Potential und Aktionspotential
Nervenzellen codieren Information in Gestalt von kurzfristigen Potentialänderungen. Diese lassen sich in zwei Gruppen unterteilen, die unterschiedliche Eigenschaften und Funktionen haben:
- graduierte Potentiale treten bei Sinneszellen (Sensorpotential, Rezeptorpotential oder Generatorpotential genannt) und an den postsynaptischen Membranen (Postsynaptisches Potential, PSP) auf. Beispiele: Membran des Nervenzellkörpers bei chemischen Synapsen oder an der Motorischen Endplatte bei Nervenzell-Muskelzell-Kontakten.
- Aktionspotentiale werden am Axonhügel und am Axon einer Nervenzelle oder an der subsynaptischen Membran von Muskelzellen erzeugt.
Vergleich in der Übersicht:
graduiertes Potential Aktionspotential 1 amplitudenmoduliert frequenzmoduliert 2 graduierte Amplitude konstante Amplitude 3 nicht refraktär refraktär 4 Summation möglich keine Summation möglich („Alles-oder-Nichts-Prinzip“) 5 Ausbreitung passiv mit Amplitudenabfall Ausbreitung aktiv mit Erhalt der Amplitude 6 keine Auslöseschwelle definierte Auslöseschwelle 7 Depolarisation oder Hyperpolarisation mit anschließender Repolarisation nur Depolarisation mit anschließender Repolarisation 8 unspezifische Kationenkanäle schnelle, spannungsgesteuerte Natrium-Ionenkanäle 9 Dauer 40 bis 4000 ms Dauer 4 ms Beispiele
- Die Schließzellen des Spaltöffnungsapparates von Pflanzenblättern halten den Zustand der Hyperpolarisation so lange aufrecht, wie sie belichtet werden. Solange pumpt eine ATPase Protonen aus der Zelle heraus. Ohne Licht stellt sie ihre Tätigkeit ein und Kalium- und Chlorid-Ionen strömen entsprechend dem chemiosmotischen Potential nach außen, wodurch das Membranpotential wieder erhöht wird (Repolarisation).
Grundlagen
Diffusionspotential
- Modellversuch
Eine Kammer, die mit destilliertem Wasser gefüllt ist, wird durch eine omnipermeable Membran, zum Beispiel ein Filterpapier, in zwei Halbzellen unterteilt. In jeder Halbzelle befindet sich eine Elektrode, die beiden Elektroden sind durch ein Spannungsmessgerät miteinander verbunden. Wird in einer der beiden Halbzellen (zum Beispiel der rechten) Kochsalz (NaCl) aufgelöst, beobachtet man zunächst einen Anstieg der Spannung, die dann mit der Zeit allmählich wieder auf Null Volt sinkt.
- Erklärung:
Aufgrund des Konzentrationsgefälles diffundieren Natrium-Kationen und Chlorid-Anionen solange durch die Membran, bis in beiden Halbzellen die gleiche Konzentration an vorliegt. Das Natrium-Kation hat einen kleineren Durchmesser als das Chlorid-Anion, es kann leichter durch das Porensystem der Membran diffundieren. Deshalb steigt zu Beginn des Versuchs ihre Konzentration in der linken Halbzelle schneller als die der Anionen. Damit bildet sich ein Spannungsunterschied zwischen den beiden Halbzellen aus: links sind mehr positive, rechts mehr negative Ladungen.
Die Diffusionsgeschwindigkeit der Kationen wird aber gebremst. Einerseits wird das Konzentrationsgefälle schwächer, andererseits müssen die Kationen gegen das sich aufbauende Potentialgefälle diffundieren.
Dagegen wird die Diffusionsgeschwindigkeit der Anionen durch das Potentialgefälle erhöht.
Gleichgewichtspotential
- Modellversuch
Eine Kammer, die mit destilliertem Wasser gefüllt ist, wird durch eine selektive, semipermeable Membran, die nur das Kation passieren lässt, in zwei Halbzellen unterteilt. In jeder Halbzelle befindet sich eine Elektrode, die beiden Elektroden sind durch ein Spannungsmessgerät miteinander verbunden. Wird in einer der beiden Halbzellen (zum Beispiel der rechten) Kochsalz (NaCl) aufgelöst, beobachtet man zunächst einen Anstieg der Spannung, die dann erhalten bleibt.
- Erklärung:
Aufgrund des Konzentrationsgefälles diffundieren die Natrium-Kationen durch die Membran. Aufgrund der Ladungstrennung baut sich ein Potentialgefälle auf: die Innenseite der Membran (linke Kammer) wird positiv, die Außenseite (rechte Kammer) negativ. Die Diffusionsgeschwindigkeit der Kationen wird aber gebremst. Einerseits wird ihr Konzentrationsgefälle schwächer, andererseits müssen die Kationen gegen das sich aufbauende Potentialgefälle diffundieren.
Das Diffusionsgleichgewicht ist dann erreicht, wenn die treibende Kraft des Konzentrationsgefälles für die Diffusion nach innen genauso groß ist, wie die treibende Kraft des Potentialgefälles für die Diffusion nach außen.
Im Gleichgewicht sind die Konzentrationen der Ionen innen von der Konzentration außen verschieden, deshalb ist eine Potentialdifferenz messbar.
Umkehrpotential
Als Umkehrpotential bezeichnet man das Membranpotential, bei dem die Ionenflüsse im Gleichgewicht sind. Das heißt, durch den entsprechenden Kanal findet kein Nettostromfluss von Ionen statt.
Eine Abweichung des Membranpotentials von diesem „Gleichgewichtspotential“ des Ions verursacht eine treibende, elektromotorische Kraft, welche das Ion entweder in oder aus der Zelle treibt. (Bei jedem Überschreiten des Umkehrpotentials ändert sich die Stromrichtung über der Membran).
Mathematische Formalismen
Die Freie Enthalpie der Diffusion
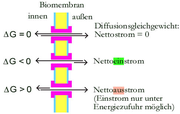
An der Freien Enthalpie ΔG kann abgelesen werden, ob Teilchen bei einem gegebenen Konzentrations- und Potentialverhältnis durch eine Membran transportiert werden können:
- Ist ΔG = 0, liegt Gleichgewicht vor, die Menge der durch die Membran in ein Richtung diffundierenden Teilchen ist gleich der Menge der durch die Membran zurück diffundierenden Teilchen.
- Ist ΔG < 0 läuft der Transport in eine Richtung freiwillig ab, wobei Energie frei wird.
- Ist ΔG > 0 können die Teilchen nur unter Energieaufwand transportiert werden.
Die Freie Enthalpie kann auch als Maß des elektrochemischen Potentials angesehen werden, das sich aus den beiden Komponenten
- chemisches Potential (entspricht dem Konzentrationsgefälle) und
- elektrisches Potential (entspricht Potentialgefälle) zusammensetzt.
Chemisches Potential - Neutrale Teilchen
Für den Transport von außen nach innen (Import) gilt die Formel
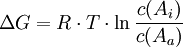
Erläuterung:
- R: Allgemeine Gaskonstante R = 8,3143 J·mol-1·K-1
- T: Temperatur in Kelvin
- c(Ai), c(Aa): Stoffmengen-Konzentrationen des Stoffes A innen, außen
- ln: natürlicher Logarithmus
Für T = 298 K und Benutzung des dekadischen Logarithmus vereinfacht sich die Gleichung zu
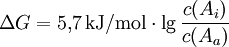
- Wenn die Konzentration des Stoffes A innen genau so groß ist wie außen, ist ΔG = 0, es liegt Konzentrationsausgleich vor und es findet kein Stofftransport statt.
- Ist die Konzentration innen größer als außen, ist ΔG > 0, es findet kein passiver („freiwilliger“) Stofftransport von außen nach innen statt.
- Ist die Konzentration außen größer als innen, ist ΔG < 0, es findet Stofftransport von außen nach innen statt.
Elektrisches Potential - Geladene Teilchen
Anteil des Ladungstransportes an der Freien Enthalpie:
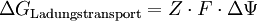
Erläuterung:
- Z: Die Ladungszahl Z entspricht der Ionenladung des zu transportierenden Teilchens. Sie ist für Kationen positiv, für Anionen negativ.
- F: Faraday-Konstante F = 96485 C·mol-1
- ΔΨ: Das Membranpotential ΔΨ lässt sich durch die Goldman-Gleichung (siehe unten) berechnen.
Elektrochemisches Potential
Für den Import geladener Teilchen gilt die Formel
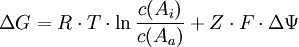
 passiver Transport durch geöffnete Ionenkanäle
passiver Transport durch geöffnete IonenkanäleGleichgewichtspotential
Für den Gleichgewichtsfall (ΔG = 0) lässt sich das Gleichgewichtspotential ΔΨ0 für ein Ion nach folgender Formel berechnen:
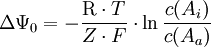
für Z = 1 (bei Na+, K+) und T = 298 K ergibt sich bei Benutzung des dekadischen Logarithmus die vereinfachte Gleichung
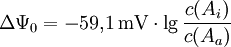
Beispiel für ein Membranpotential (Mischpotential) von –53 mV bei 298 K:
Ionensorte ceq(Aaußen) ceq(Ainnen) ΔΨ0 ΔG für einen Transport von außen nach innen Na+ 400 mmol/l 20 mmol/l +76 mV +2,3 kJ/mol K+ 50 mmol/l 440 mmol/l -55 mV -10,5 kJ/mol Cl- 108 560 -43 mV -11,0 kJ/mol Goldman-Hodgkin-Katz-Gleichung
Mittels der Goldman-Gleichung lässt sich das Membran-Gleichgewichtspotential ΔΨ berechnen:
![\Delta \Psi = \frac {R T}{F} \cdot \ln \frac {\Sigma[P_\mathrm{ka} \cdot c_\mathrm{ka-a}] + \Sigma [P_\mathrm{an} \cdot c_\mathrm{an-i}]}{\Sigma [P_\mathrm{ka} \cdot c_\mathrm{ka-i}] + \Sigma [P_\mathrm{an} \cdot c_\mathrm{an-a}]}](/pictures/dewiki/56/8a8482fca525b5286ef28e43b00df63d.png)
Erläuterung:
- P: Permeabilität der Kanäle für Anionen (an) und Kationen (ka)
- c: Konzentration der Anionen und Kationen innerhalb (-i) oder außerhalb (-a) der Zelle
Siehe auch
- Ionenkanal, Transmembranprotein, Kaliumkanal, Transport (Biologie),
- Depolarisation (Physiologie), Hyperpolarisation (Biologie)
- Ruhemembranpotential, Aktionspotential
Weblinks
Wikimedia Foundation.