- Kelchwürmer
-
Kelchwürmer 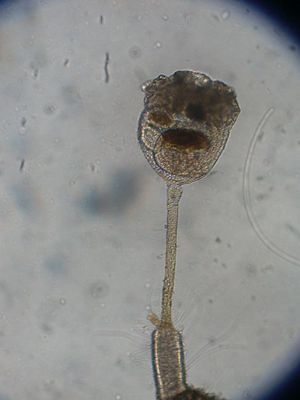
Barentsa discreta
Systematik Unterreich: Vielzellige Tiere (Metazoa) Abteilung: Gewebetiere (Eumetazoa) Unterabteilung: Bilateria Überstamm: Urmünder (Protostomia) Überstamm: Plattwurmartige (Platyzoa) Stamm: Kelchwürmer Wissenschaftlicher Name Entoprocta Nitsche, 1869 Die Kelchwürmer, auch Kelchtiere (Entoprocta (Gr.: mit innerem After), auch Kamptozoa) genannt, sind ein Tierstamm mit etwa 250 Arten. Sie leben im Wasser als sessile Filtrierer und werden 0,5 bis 5 Millimeter lang. Ihren wissenschaftlichen Namen (Entoprocta / griechisch entos = „Innere“ + proktos = „Anus“) haben sie, weil bei ihnen im Gegensatz zu den äußerlich ähnlichen Moostierchen (Bryozoa oder Ectoprocta) der Anus innerhalb des Tentakelkranzes liegt. Sie leben kommensal auf Schwämmen und anderen Lebewesen. Dabei schaden sie ihren „Wirten“ nicht.
Inhaltsverzeichnis
Merkmale
Anatomie der Adulten
Der Aufbau aller Kelchwürmer ist sehr einheitlich. Sie bestehen aus einem muskulösem Stiel, an dessen unterem Ende ein Fuß mit einer Klebedrüse ausgebildet ist, sowie einem Kelch, der den eigentlichen Körper darstellt und eine Tentakelkrone trägt. Die Größe der Einzeltiere (Zooid) beträgt im Normalfall etwa einen Millimeter, die kleinste bekannte Art ist Loxomespilon perezi mit etwa 0,1 Millimeter und die größte Barentsia robusta mit etwa sieben Millimetern Körpergröße. Die Einzeltiere sind bilateralsymmetrisch aufgebaut und ähneln bei oberflächlicher Betrachtung den Polypen der Hydrozoa, die zu den Nesseltieren gehören. Anders als bei diesen sind bei den Kelchwürmern allerdings die Tentakel nicht einziehbar, sondern können nur eingerollt werden.
Die Außenschicht der Tiere wird durch eine einschichtige Epidermis gebildet, die nach außen durch eine Gallertschicht begrenzt wird. Darunter liegt einlagig die Muskulatur, die im Kelchbereich aus schräggestellten Fasern besteht und im Stiel in eine Längsmuskulatur übergeht. Als Leibeshöhle besitzen die Tiere ein flüssigkeitsgefülltes Pseudocoel mit vereinzelten Ansammlungen von Mesenchymzellen, bei dem keine epitheliale Auskleidung des freien Raumes vorhanden ist. Dieses durchzieht den Körper vom Stiel über den Kelch bis hin zu den einzelnen Tentakeln, ein zusätzliches Gefäßsystem existiert nicht.
Im Kelch befinden sich die Organe der Tiere, wobei der U-förmige Darmkanal den Großteil der Raumes einnimmt. Der Darm besitzt keine eigene Muskulatur und ist im Innenraum von Cilien bestanden, die die Nahrung zum zentralen Abschnitt, dem Magen, transportieren. Hier erfolgt die Verdauung und auch die Aufnahme der Nährstoffe, außerdem ist das Magendach ähnlich der Leber der Wirbeltiere das Hauptspeicher- und -stoffwechselorgan und dient zusätzlich der Exkretion, indem Stoffwechselendprodukte aus der Leibeshöhle in den Darmkanal abgegeben werden. Die Mundöffnung und der After münden beide nach oben gerichtet in den Bereich des tentakelumstandenen Raums (Atrium), ebenso die der Exkretion und Osmoregulation dienenden paarigen Protonephridien sowie die Ausführgänge der sackartigen Gonaden.
Das Nervensystem ist einfach gebaut und besitzt ein einfaches, hantelförmiges Ganglion in der Schleife des Darms. Von diesem ziehen Nervenstränge zur Kelch-, zur Tentakel- und zur Stielmuskulatur. Auf der Kelchoberfläche befinden sich Mechanorezeptoren und an den Tentakelinnenseiten zusätzlich Sinnescilien, Lichtsinnesorgane besitzen die Tiere nicht.
Anatomie der Larven
Die Larve der Kelchwürmer hat einen Durchmesser von durchschnittlich nur 50 bis 100 Mikrometern und zeigt eine Reihe von Ähnlichkeiten zum weit verbreiteten Trochophora-Larventyp. Unterschiede bestehen unter anderem in der kuppelförmigen Ausbildung der oberen Hälfte (Episphäre) mit einer Scheitelplatte am oberen, apikalen Ende. Aufgrund dieser Kuppelform wurde der Name „Tholophora-Larve“ (von gr. tholos=Kuppeldach, Gewölbe) vorgeschlagen (Salvini-Plawen 1980). Am unteren Rand der Episphäre zieht ein Wimpernkranz entlang, der während der anfänglichen schwimmenden (pelagischen) Lebensweise der Larve als Motor der Fortbewegung dient. Auf der Episphäre ist bei den meisten Arten ein Sinnesorgan ausgebildet, welches aus einem Wimpernbüschel und bei später einzellebenden Tieren einem Paar Lichtsinnesorgane (Ocellen) besteht (Praeoralorgan). Bei den später solitären Formen ist das Praeoralorgan paarig, bei koloniebildenden Formen unpaar.
Die untere Hälfte der Larve, die Hyposphäre, ist während der Schwimmphase vollständig in die Episphäre eingestülpt und somit von außen kaum sichtbar. Dies war der Hauptgrund für die anfängliche Gleichsetzung mit dem Trochophora-Larventyp. Erst als klar wurde, dass in einer zweiten, bodenlebenden (benthischen) Phase die ausgestülpte Hyposphäre als Kriechsohle eingesetzt wird, wurde die Gleichsetzung mit der Trochophora relativiert (bei einigen Kamptozoen-Arten fehlt allerdings diese zweite bodenlebende Phase). Die Mundöffnung liegt am Rand der Hyposphäre unterhalb des Wimperkranzes und führt über einen U-förmigen Darmkanal zum ebenfalls an der Hyposphäre liegenden Anus. Außerdem besitzt die Tholophora Protonephridien, die der Exkretion dienen.
Lebensweise
Kelchwürmer leben als Filtrierer, wobei sie mit Hilfe ihrer Tentakel Schwebstoffe aus dem Wasser filtern. Diese sondern einen klebrigen Schleim ab, an dem die Partikel kleben bleiben, über die Cilien der Tentakel werden diese der Mundöffnung zugeführt. Durch Kontraktion der Längsmuskulaturstränge führen die Einzeltiere Pendelbewegungen ihres Stieles sowie charakteristische Nickbewegungen des Kelches durch, um die Filtrationsleistung zu steigern.
Die meisten Arten leben solitär und sind mit ihrem Fuß am Boden befestigt, etwa ein Drittel der Arten bilden jedoch auch Kolonien mit mehreren Individuen, die über Äste aus ihren Stielen miteinander verbunden sind
Fortpflanzung und Entwicklung
Kelchwürmer können sich sowohl sexuell als auch asexuell fortpflanzen. Während die Pedicellinidae grundsätzlich simultan zwittrig sind, sie besitzen also dauerhaft sowohl männliche als auch weibliche Geschlechtsorgane, stellen die Loxosomatidae protandrische Zwitter dar, bei ihnen entwickeln sich also zuerst männliche und anschließend weibliche Geschlechtsorgane.
Die Barentsidae sind getrenntgeschlechtlich, innerhalb einer Kolonie kommen dabei beide Geschlechter vor. Dabei bilden sich grundsätzlich immer zuerst Männchen mit Hoden aus, die Entwicklung weiblicher Zooide ist abhängig von der Präsenz bereits vollentwickelter Männchen in der Kolonie. Die Lebenszeit der Einzeltiere beträgt etwa sechs Wochen.
Asexuelle Fortpflanzung
Die asexuelle Vermehrung erfolgt dabei durch Knospung der ektodermalen Kelchwand, bei der aus einem Einzeltier mehrere genetisch mit diesem identische Nachkommen resultieren. Die Knospen entstehen in der Kelchwand im Bereich des Mundes oder, bei koloniebildenden Formen, am Stiel. Erst nach der abgeschlossenen Knospung wandern mütterliche Mesenchymzellen in das neu entstandene Tier und bilden die Muskulatur sowie die Bindegewebe.
Bei den koloniebildenden Arten bleiben die Jungtiere über den Stiel mit dem Muttertier verbunden und bilden einen verbindenden Stolo durch Längenwachstum an der Verbindungsstelle. Als Ergebnis bilden sich mattenförmige oder stark verzweigte, bäumchenförmige Kolonien, die bei einigen Arten wie etwa Pedicellinopsis fructiosa Höhen von bis zu 2,5 Zentimeter mit Tausenden einzelnen Zooiden erreichen.
Die Regenerationsfähigkeit der Kamptozoen ist relativ beschränkt und betrifft nur die Neubildung verletzter Tentakel. Bei den koloniebildenden Formen können die Kelche aus dem Blastoderm des Stieles regeneriert werden, die Stiele können also als Überdauerungsstadien dienen (vor allem bei Barentsiidae). Einige Kolonien bilden zudem stark retardierte Knospen aus, die erst zu echten Zooiden werden, wenn die restliche Kolonie abgestorben ist.
Sexuelle Fortpflanzung
Bei der sexuellen Fortpflanzung geben die männlichen Tiere ihre Spermien in das freie Wasser ab, von dort werden sie von den weiblichen Tieren eingestrudelt. Die Befruchtung findet im Ovar der Weibchen statt, von dort wandern die befruchteten Zygoten in das Atrium und lagern sich in spezielle paarige Bruttaschen ein, die beidseitig des Enddarms liegen.
Die Embryonalentwicklung verläuft über eine Spiralfurchung, bei der sich Quartette von jeweils zwei kleineren (Mikromere) und zwei größeren Zellen (Makromere) bilden und spiralig gegeneinander lagern. Aus den 4d-Zellen entwickeln sich Telomesoblasten, die die Ursprungszellen des gesamten mesodermalen Gewebes sind – Es handelt sich also um die für die Spiralia typische Spiralquartett-4d-Furchung. Aufgrund dieses Merkmales muss eine nähere Verwandtschaft der Kelchwürmer mit anderen Spiraliern in Betracht gezogen werden, diskutiert werden hier vor allem Ringelwürmer und Weichtiere (siehe Abschnitt Systematik).
Verbreitung und Lebensraum
Kelchwürmer sind weltweit in den Gewässern der Küstengebiete zu finden. Die meisten Arten sind marin, die weitverbreitete Art Urnatella gracilis lebt als einzige Art im Süßwasser, sie kam ursprünglich wahrscheinlich nur in Nordamerika vor und wurde von dort nach Europa und Asien eingeschleppt.
Die Einzeltiere siedeln sehr häufig auf anderen wirbellosen Tieren wie Schwämmen, Ringelwürmern, Stachelhäutern und seltener auch auf Krebstieren, wo sie von der Verwirbelung des Wassers durch die Bewegung und Nahrungsaufnahme der größeren Tiere profitieren. Kolonien leben auf allen Substraten, die einer Strömung ausgesetzt sind, dabei vor allem auf Hydroidstöcken, Muschelschalen und Gesteinsflächen.
Stammesgeschichte
Aufgrund ihrer Kleinheit sowie der fehlenden Hartsubstanzen ihres Körpers ist der stammesgeschichtliche Nachweis der Kelchwürmer nur sehr lückenhaft. Die ältesten bekannten Formen stammen aus dem späten Jura und wurden in England gefunden.
Systematik
Die externe Systematik der Kelchwürmer ist bislang nicht vollständig geklärt. Ursprünglich wurden sie von Nitsche vollständig den Moostierchen (Bryozoa) zugeordnet, von den meisten Bearbeitern wurde diese Einordnung jedoch abgelehnt. Auf der Basis von ontogenetischen Beobachtungen werden sie heute noch von einzelnen Autoren, vor allem von Nielsen 1979[1], in die verwandtschaftliche Nähe der Bryozoa gestellt. Ax 1999[2] schlägt dagegen als Alternative eine Einordnung in die Verwandtschaft der Weichtiere (Mollusca) vor und benennt das entstehende Taxon als Lacunifera, wobei er den Feinaufbau der Cuticula, den Aufbau der Leibeshöhle als Lakunensystem sowie den Feinaufbau der Cilien als Argumente anführt. Ein weiterer Vorteil dieser Hypothese wird darin gesehen, dass ein evolutionärer Zusammenhang zwischen den beschalten, trochophoraartigen Larven mancher Weichtiere und den ähnlich erscheinenden Kriechlarven der Kelchwürmer hergestellt werden kann (im Extremfall wird sogar der Kriechfuß der Weichtiere von der Kriechsohle der Kelchwurm-Larve hergeleitet!).
Molekularbiologisch konnte allerdings weder eine Nähe der Kelchwürmer zu den Moostierchen, noch die Ax’sche „Lacunifera“-Hypothese bestätigt werden (vgl. auch weiter unten).
Wenn man die Kriechlarven der Kelchwürmer als modifizierte Trochophoralarven betrachtet, sind bezüglich der Herkunft mindestens zwei Evolutionsmodelle zu formulieren, in denen jeweils eine Abstammung von annelidenartigen Vorformen (also Ringelwürmern im weitesten Sinne) angenommen wird. Aufgrund der Organisation der Kelchwürmer als coelomlose Tiere mit Spiralfurchung und keiner erkennbaren Gliederung wird manchmal vermutet, dass die Kelchwürmer sich direkt auf Trochophorae zurückführen lassen, die im Jugendstadium bereits adult wurden (progenetische Trochophorae). Eine zweite Version sieht in den Kelchwürmern schrittweise verzwergte und umgewandelte Ringelwürmer, die Veränderungen der Larvenform würden nach diesem Modell parallel zur Umwandlung der Adultformen abgelaufen sein. Eine gewisse Nähe zu den Ringelwürmern wurde durch erste molekulargenetische Untersuchungen der rRNA (Mackay et al. 1995[3]) bestätigt, allerdings zeigen auch viele andere Lophotrochozoa-Linien molekulargenetische Ähnlichkeiten mit den Ringelwürmern (darunter auch die Weichtiere). Neueren genetischen Untersuchungen zufolge bilden Kelchwürmer die Schwestergruppe einer anderen sessilen Zwergform, nämlich der erst 1995 beschriebenen Cycliophora (Halanych 2004). Damit wäre also die Annahme eines Schwestergruppenverhältnisses zu den Moostierchen oder Weichtieren zurückgewiesen, während die Abstammung von ringelwurmartigen Vorformen in der Diskussion bleibt.
Es gibt etwa 150 Arten der Kelchwürmer, die in vier Familien eingeordnet werden. Die solitären und als ursprünglich angesehenen Loxosomatidae enthalten dabei etwa zwei Drittel der bekannten Arten und werden den anderen Familien als basale Schwestergruppe Solitaria gegenüber gestellt. Die Vertreter der anderen Familien bilden Kolonien und werden als Coloniales zusammengefasst, hier wiederum werden die Astolonata (Loxokalypodidae) den Stolonata gegenübergestellt. Als phylogenetisches System resultiert daraus:
Kelchwürmer Coloniales Astolonata (Loxokalypodidae)
Stolonata Barentsiidae
Pedicellinidae
Solitaria (Loxosomatidae)
Die einzelnen Taxa enthalten folgende Gattungen:
- Loxosomatidae: Loxosoma, Loxosomella, Loxomitra, Loxosomespilon, Loxocore
- Loxokalypodidae: Loxokalypus
- Barentsiidae: Barentsia, Urnatella, Pedicellinopsis, Pseudopedicellina, Coriella
- Pedicellinidae: Pedicellina, Pedicellina, Myosoma, Chitaspis, Loxosomatoides
Literatur
- Peter Emschermann (2004): Kamptozoa (Entoprocta), Kelchwürmer, in: W. Westheide, R. Rieger (Hrsg.): „Spezielle Zoologie Teil 1: Einzeller und Wirbellose Tiere“; 2.Aufl. Gustav Fischer Verlag, Stuttgart, Jena. ISBN 3-437-20515-3
- Peter Ax (1999): Das System der Metazoa: ein Lehrbuch der phylogenetischen Systematik, Gustav Fischer Verlag, Stuttgart. ISBN 3-437-35528-7
- Luitfried von Salvini-Plawen (1980): Was ist eine Trochophora? Eine Analyse der Larventypen mariner Protostomier. Zool. Jb. Anatomie 103: 389-423
- Kenneth M. Halanych (2004): The new view of animal phylogeny. Annu. Rev. Ecol. Evol. Syst. 35: 229-256
Weblinks
 Commons: Entoprocta – Sammlung von Bildern, Videos und Audiodateien
Commons: Entoprocta – Sammlung von Bildern, Videos und AudiodateienQuellen
- ↑ Nielsen C.: Larval ciliary bands and metazoan phylogeny, Fortschritte der Zoologischen Systematischen Evolutionsforschung 1, 1979
- ↑ Ax P: Das System der Metazoa: ein Lehrbuch der phylogenetischen Systematik, G. Fischer Verlag, Stuttgart 1999
- ↑ Mackay LY, Winnepennickx B, de Wachter R, Backeljau T, Emschermann P, Garey J: 18S rRNA suggests that Entoprocta are protostomes, unrelated to Ectoprocta, Journal of Molecular Evolution 41, 1995


Dieser Artikel wurde in die Liste der lesenswerten Artikel aufgenommen. Kategorien:- Wikipedia:Lesenswert
- Vielzellige Tiere
Wikimedia Foundation.