- Kin-Selektion
-
Die Verwandtenselektion (kin selection, Sippenselektion) ist eine Erweiterung des Begriffs der natürlichen Selektion und wird in der Evolutionsbiologie und der Soziobiologie gebraucht. Im Rahmen der Gesamtfitness-Theorie erklärt sie die Vererbung von kooperativem und „altruistischem“ Verhalten. Wenn Tiere Verwandten dabei helfen, ihre Jungen aufzuziehen, fördert dies die Weitergabe ihres 'eigenen' Erbgutes. Das Ausmaß an altruistischem Verhalten richtet sich nach dem Grad der Verwandtschaft. Je enger Tiere miteinander verwandt sind, desto höher ist die Wahrscheinlichkeit, durch Verwandtenhilfe 'eigene' Gene in die nächste Generation weiterzugeben und desto häufiger ist altruistisches Verhalten anzutreffen. Die Theorie der Verwandtenselektion wurde von John Maynard Smith (1964) und William D. Hamilton entwickelt.
Inhaltsverzeichnis
Gesamtfitness
Gesamtfitness = direkte Fitness + indirekte Fitness
Die genetische Gesamtfitness (engl. "inclusive fitness"), der genetische Erfolg eines Lebewesens, misst sich an der Anzahl der eigenen Gene, die an die nachfolgende Generation weitergegeben wird. Sie setzt sich zusammen aus der direkten Fitness, der Anzahl der Gene, die durch eigene Nachkommen weitergegeben wird, und der indirekten Fitness, der Anzahl der eigenen Gene, die über Verwandte an die nächste Generation weitergegeben wird. Ein Individuum, das die Fortpflanzungschancen eines nahen Verwandten erhöht, bewirkt so eine Erhöhung seiner eigenen Gesamtfitness (= genetischer Erfolg).
Dieser Altruismus ist nur dann erfolgreich und breitet sich in Populationen aus, wenn der Nutzen für denjenigen, der das altruistische Verhalten zeigt, größer ist als die Kosten, die er dafür aufbringen muss (Hamiltons Regel).
Mathematisch ausgedrückt muss das Verhältnis von Nutzen (B) zu Kosten (C) größer sein als 1 dividiert durch den Verwandtschaftsgrad.
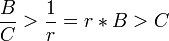
mit B: Nutzen (benefit); C: Kosten (cost); r: Verwandtschaftskoeffizient (relatedness)
Unter Einbeziehung der verschiedenen Verwandtschaftsgrade zum Empfänger und zu den eigenen Nachkommen ergibt sich folgende Formel (Hamilton's rule):
rB * B > rC * C
rB: Verwandtschaftsgrad des Gebers zum Empfänger; rC: Verwandtschaftsgrad des Gebers zu den eigenen Nachkommen
Beispiel 1: Ein Tier, das auf zwei eigene Nachkommen verzichtet (C = 2), dafür aber einem Geschwister (Verwandtschaftsgrad zwischen Geschwistern bei diploiden Organismen (r = 0.5)) hilft, fünf zusätzliche Nachkommen (B = 5) zu produzieren, hat eine höhere Gesamtfitness als ein Tier, das „egoistisch“ nicht hilft. r * B > C ; 0,25 * 5 > 2 * 0,5
Beispiel 2: Viele Arbeiterinnen bei Insektenvölkern verzichten auf eigene Nachkommen und opfern für die Verteidigung des Volkes sogar ihr Leben. Die Verwandtenselektion gibt dafür eine plausible Erklärung. Aufgrund der ungewöhnlichen Haplodiploidie hymenopterer sozialer Insekten (Ameisen, Bienen und Wespen) ergibt sich bei Vollschwestern eines Nestes ein Verwandtschaftskoeffizient von durchschnittlich 0,75 miteinander, mit ihren Vollbrüdern 0,25. Mit ihren eigenen Nachkommen sind diese Arbeiterinnen jedoch nur zu 50 % (r = 0,5), also weniger als mit den Schwestern, verwandt. Als Folge ist es für Arbeiterinnen sozialer Hymenopteren genetisch vorteilhafter, eigene Schwestern als Töchter aufzuziehen. Dadurch verbreiten sie ihre Gene mehr, als wenn sie eigene Nachkommen hätten.
Beispiel 3: Wenn ein Mensch sein Leben opfert, aber zwei Geschwister dafür überleben, macht das für seine Gene keinen Unterschied; rettet er drei Geschwister, ist das für seine Gene ein Gewinn. Aus Sicht der Gesamtfitness sollte eine Person ihr Leben opfern, wenn sie dadurch mehr als zwei ihrer Kinder, vier Neffen oder acht Cousins rettet, da ein Kind 50%, ein Neffe 25% und ein Cousin 12.5% der Gene mit ihr gemeinsam hat.
Gruppenstruktur
Die Verwandtenselektion macht auch Aussagen zu Gruppenstruktur von Populationen. Wann bildet sich eine ungleiche Rangordnung heraus, bei der sich nur die Ranghöchsten fortpflanzen? Um seine Gesamtfitness zu maximieren sollte das Individuum je nach den Umweltbedingungen den Schwerpunkt auf die Erhöhung der direkten Fitness oder die Erhöhung der indirekten Fitness legen.
Sind die Umweltbedingungen günstig und der mögliche Fortpflanzungserfolg groß, dann sollte ein Individuum abwandern und seine Gene durch eigene Nachkommen verbreiten. Die Hierarchieunterschiede innerhalb einer Gruppe sind dann gering.
Sind die Umweltbedingungen ungünstig und der mögliche Fortpflanzungserfolg gering, dann sollte das Individuum zu Hause bleiben. Es bilden sich dann hierarchische Gruppenstrukturen mit starker Rangordnung heraus.
Konkurrierende / weiterführende Konzepte
- Gruppenselektion 1962 Vero Wynne-Edwards
- Reziproker Altruismus‚ 'Wie du mir, so ich dir', tit for tat; Robert L. Trivers
- Optimal-Skew-Theorie, Laurent Keller, H. Kern Reeve
- Eine mathematisch elegante Synthese der Verwandtenselektion (oder allgemeiner der individuellen Selektion unter Berücksichtigung genetischer Verwandtschaften) und der Gruppenselektion stellt die Price-Gleichung dar, in der Individual- wie auch Gruppenselektion berücksichtigt werden.
Siehe auch
Wikimedia Foundation.