- Selektion (Evolution)
-
Selektion (lat. selectio „Auswahl/Auslese“) ist ein grundlegender Begriff der Evolutionstheorie. Sie besteht
- als natürliche Selektion in der Reduzierung des Fortpflanzungserfolgs bestimmter Individuen einer Population mit der Folge, dass andere Individuen, die im Rückblick als „überlebenstüchtiger“ erkennbar sind, sich stärker vermehren. Die entscheidenden Einflüsse üben äußere Faktoren der Umwelt, sog. Selektionsfaktoren, aus. „Überlebenstüchtigkeit“ (Fitness) bedeutet nicht das „Überleben der Stärksten“. Sie kann auch Kooperation und Altruismus einschließen. Entscheidend ist, dass die Erbanlagen von Individuen nicht mit der gleichen Wahrscheinlichkeit weitergegeben werden.
- als sexuelle Selektion in der Auswahl von Individuen durch die Sexualpartner. Entscheidend ist, dass Erbanlagen derjenigen Merkmale weitergegeben werden, die von den Sexualpartnern bevorzugt werden.
- als künstliche Selektion in einer vom Menschen gesteuerten Zuchtwahl. Sie steigert den Fortpflanzungserfolg jener Individuen, die die vom Züchter geförderten Eigenschaften besitzen.
Die Bezeichnung natürliche Selektion wurde von Charles Darwin geprägt. Die Selektionstheorie ist ein Aspekt seiner Evolutionstheorie und wurde als wesentlicher Teil der Synthetischen Evolutionstheorie in die moderne Evolutionsbiologie übernommen. Selektion ist einer der Evolutionsfaktoren.
Natürliche Selektion
Grundlage der natürlichen Selektion ist die jeweilige Wahrscheinlichkeit, mit der Individuen ihre Erbanlagen an die Folgegeneration weitergeben. Falls die Individuen einer Population sich in einem oder mehreren Merkmalen voneinander unterscheiden (das ist in natürlichen Populationen in der Regel der Fall), bewirkt die Selektion einen unterschiedlichen Fortpflanzungserfolg, indem einige Individuen länger überleben, mehr Nachkommen produzieren können, Feinden besser entkommen oder widerstehen können, resistenter gegen Krankheiten sind, u.s.w.. Man sagt dazu, die Individuen mit höherem Fortpflanzungserfolg besitzen eine höhere Fitness. Die selektierten Merkmale, die die höhere Fitness bewirken, können genetisch bedingt (erblich) sein, oder es kann sich um umweltbedingte Varianten oder Modifikationen handeln. Evolutionär wirksam ist nur die Selektion erblicher Merkmale. Die Erbanlagen der fitteren Individuen sind dann in der Folgegeneration mit einem größeren Anteil vertreten als in der Parentalgeneration, d.h. zwangsläufig gleichzeitig, dass Individuen mit (in ihrer aktuellen Umwelt) ungünstigeren Merkmalen in der Folgegeneration mit geringerer Häufigkeit vertreten sind. Die unterschiedlichen Individuen besitzen nur in seltenen Ausnahmefällen vollkommen unterschiedliche Gene. In der Regel ist es so, dass die vererbten Unterschiede auf geringfügige Varianten desselben Gens (Allele) zurückzuführen sind. (Die meisten dieser Unterschiede betreffen sogar nur ein einzelnes Basenpaar: SNPs). Für die Populationsgenetik bedeutet Selektion deshalb dasselbe wie gerichtete (d.h. nicht zufällige) Verschiebung der Allelfrequenz in der Population.
Gegenstand der Selektion sind alle erblichen Merkmale, die zu einem Unterschied in der Fortpflanzungsrate führen können. Neben dem Tod des Individuums sind auch z.B. solche Individuen selektiv benachteiligt, die eine geringere natürliche Reproduktionsrate besitzen oder die gegenüber Artgenossen bzw. Individuen anderer Arten in der Konkurrenz benachteiligt sind. Entscheidend ist dabei nicht die Fortpflanzungsrate als solche, sondern ausschließlich der Anteil der erfolgreichen Nachkommen (d.h. derjenigen, die selbst Nachkommen hinterlassen).
Sexuelle Selektion
Die sexuelle Selektion ist ein wichtiger Sonderfall der natürlichen Selektion. Sie ergibt sich aus der Konkurrenz der Geschlechter um Fortpflanzungspartner des anderen Geschlechts innerhalb einer Art. Zahlreiche Merkmale von Arten, die mit der natürlichen Selektion zunächst nicht erklärbar sind, weil sie mit einem Überlebensnachteil für ihren Träger verbunden sind, lassen sich dadurch erklären, dass sie die Wahrscheinlichkeit ihres Trägers erhöhen, sich erfolgreich zu paaren und dadurch seine Nachkommenzahl zu steigern. Wichtig ist die sexuelle Selektion beispielsweise zur Erklärung des Sexualdimorphismus zwischen den Geschlechtern, zur Erklärung des Geschlechterverhältnisses und zur Interpretation des Verhaltens und der sozialen Systeme zahlreicher Tierarten.
Künstliche Selektion
Bei der künstlichen Selektion handelt es sich um einen Spezialfall der Selektion, nämlich die Selektion durch den Menschen. Dabei wird ein ausgewähltes Merkmal oder eine Kombination von Merkmalen in einer Population durch den Menschen gefördert. Individuen, welche diese Eigenschaften nicht aufweisen, werden von der Fortpflanzung ausgeschlossen.
Zucht
Typischerweise findet Auswahl durch Menschen bei Haustieren oder angebauten Pflanzen, aber auch im Labor oder in befischten Gewässern statt. Der Unterschied zur natürlichen Selektion besteht darin, dass die Überlebens- und Fortpflanzungskriterien vom Menschen ausgewählt werden und mit einem speziellen Ziel, meistens zur genetischen Umformung oder Verstärkung gewollter beziehungsweise Unterdrückung ungewollter Eigenschaften, verbunden sind.
Natürliche Selektion unter dem Einfluss des Menschen
Durch den überwältigenden Einfluss, den die Spezies Mensch auf unseren Planeten ausübt, ist der Mensch in jüngster Zeit zu einem wichtigen Selektionsfaktor geworden. Wenn sein Einfluss auf die betrachtete Population nicht auf bewusste Auswahl zurückgeht, sondern es sich um unbeabsichtigte Folgen seines Eingreifens handelt, handelt es sich nicht um künstliche, sondern um eine Form der "natürlichen" Selektion, auch wenn der Ausdruck hier unglücklich gewählt erscheint. Dies gilt z.B. bereits für das bekannte Lehrbuch-Beispiel des Industriemelanismus beim Birkenspanner.
Ein frappierendes Beispiel für einen solchen Selektionsfaktor stellt die industrielle Hochseefischerei dar. Durch die Überfischung von Speisefischpopulationen wird ein starker Selektionsdruck auf bestimmte Fischarten ausgeübt, wobei große und fortpflanzungsfähige Fische aus den Populationen entfernt werden. So wird künstlich das Überleben von kleineren und frühreiferen Fischen gefördert[1] Diese haben als einzige die Chance, durch die engmaschigen Netze zu entkommen und sich anschließend noch fortzupflanzen. Die Fische investieren mehr Energie in ihre Vermehrung als in das Wachstum. So waren Kabeljau aus dem Nordost-Atlantik vor 60 Jahren, als die Jagd auf sie begann, im Schnitt noch 95 cm groß, heute erreichen sie nur noch eine Körpergröße von 65 cm. Ebenso setzt die Geschlechtsreife heute schon drei Jahre früher, nämlich mit einem Alter von sechs Jahren, ein[2]. Mit Hilfe von Computermodellen, die von Ulf Dieckmann (Ökosystemforscher am Institut für Angewandte Systemanalyse IASA, Laxenburg in Österreich, http://www.iiasa.ac.at/~dieckman/) entwickelt wurden, lässt sich zeigen, dass sich Fischpopulationen unter dem Druck der Fangflotten innerhalb von 40 Jahren erheblich verändern können, indem beispielsweise die Durchschnittsgröße stark sinkt. Würde die Befischung heute gestoppt werden, so würde es Modellberechnungen zur Folge bis zu 250 Jahren dauern, bis die Fischbestände wieder die ursprüngliche Größenverteilung erreicht hätten. Dies hängt damit zusammen, dass die Natur keinen so starken Selektionsdruck ausübt wie die Fischerei. Die Frühreife der Fische wird, so Dieckmann, durch das Fehlen der größeren Fische begründet, die als Konkurrenten wirken. Der Fisch findet mehr Nahrung, wächst schneller und wird früher geschlechtsreif.
Die Modelle für das schnelle Wirken von Selektion und die damit resultierende Mikroevolution wurden in verschiedenen Labors mit Experimenten nachgestellt. Hierfür wurde mit Fischen wie Guppies und Ährenfischen experimentiert, die eine relativ kurze Generationsdauer haben. Siehe auch verschiedene Artikel von David Conover (State University of New York)[3] und David Reznick (University of California)[4]. Beide Forscher konnten zeigen, dass bei selektiver Befischung der Populationen in den Aquarien – das heißt, nur die größten Fische wurden entnommen – schon nach wenigen Generationen die Fische deutlich kleiner und weniger fruchtbar waren. Außerdem setzten sie bei gleichem Futterangebot wie die Kontrollfische weniger Fleisch an. „Rechnet man die Evolutionsrate bei den Guppies auf die Entwicklung kommerziell genutzter Fische um, so entspricht sie wenigen Jahrzehnten“, fasst Reznick seine Ergebnisse zusammen. „Die Großfischerei hat eine genetische Selektionswirkung auf die Bestände.“
Die Ebene der Selektion
Innerhalb der Biologie wird darüber gestritten, auf welche biologischen Einheiten die Selektion wirkt[5][6]. Zu unterscheiden sind hier Gruppenselektion, Individualselektion und Genselektion.
Damit Evolution funktioniert, muss die Selektion an erblichen Eigenschaften eines Individuums ansetzen. Eine Selektion nicht vererbter Eigenschaften ist zwar genauso möglich, führt aber eben nicht zur Evolution. Da Gene nicht direkt agieren können, sondern zu ihrer Aktivität einer Maschinerie oder eines "Vehikels" bedürfen, ist außerdem eine von Genen zumindest teilweise gesteuerte Überlebenseinheit erforderlich: ein Individuum. Individuen können sich zu Gruppen zusammenschließen, biologische Gruppen besitzen aber zumindest im Regelfall keine Individualität (Ausnahmen werden für einige Sonderfälle, v.a. Insektenstaaten, diskutiert). Das bedeutet, dass sich die Eigenschaften der Gruppe aus derjenigen der Individuen ergeben. Die Ebene, auf der die Selektionsfaktoren ansetzen (die Ebene der "Interaktoren"), ist damit zunächst immer die Ebene des Individuums. Damit ist das Individuum das Objekt der Selektion[7]. Dennoch kann es sinnvoll sein, je nach Fragestellung das Gen oder die Gruppe in das Zentrum des Interesses zu stellen. Hierbei geht es weniger um tatsächliche Unterschiede, sondern um unterschiedliche Blickwinkel auf dieselbe Sache. Populationsgenetiker sind z.B. besonders interessiert an der Auswirkung der Selektion auf verschiedene Allele oder Gene. Betrachten sie z.B. die Auswirkungen eines Selektionsfaktors auf die Genfrequenz, ist die Ebene des Individuums für diese Frage bedeutungslos, da es sich um eine Eigenschaft der Population handelt. Hier geht es dann weniger um den Selektionsmechanismus, sondern um seine Auswirkungen.
Die früher einflussreiche, außerhalb der Wissenschaft noch weit verbreitete Vorstellung eines Arterhaltungstriebes ist nur noch von historischem Interesse. Sie ist durch die synthetische Evolutionstheorie (auf Basis von Populationen) überwunden. Dennoch ist es natürlich weiterhin sinnvoll, ggf. auch Vergleiche auf Artebene anzustellen. Der Unterschied liegt auch hier in den Mechanismen.
Auch die Theorie der Genselektion, die Richard Dawkins in seinem einflussreichen Buch "Das egoistische Gen" formuliert hatte, wird heute nicht mehr vertreten - auch nicht von Dawkins selbst. (Jeder Evolutionsprozess ist grundsätzlich mit einer Änderung der Genfrequenzen innerhalb der Population verbunden). Der Ausdruck Genselektion (oder Gen-Level-Selektion) wird heute manchmal in einem anderen Sinne verwendet, um Selektionsvorgänge zwischen Genen innerhalb eines einzelnen Genoms zu beschreiben, z.B. im Zusammenhang mit Meiotic Drive.
Eine mathematisch elegante Synthese der beschriebenen Selektionsmechanismen stellt die Price-Gleichung dar, in der Individual- wie auch Gruppenselektion berücksichtigt werden.
Gruppenselektion, Verwandtenselektion und Mutualismus
Während die Selektionstheorie auf die meisten morphologischen und Verhaltens-Merkmale im Prinzip problemlos anwendbar ist, ist es ein Problem, soziale Verhaltensweisen durch die Evolutionstheorie zu erklären, die nicht das jeweilige Individuum selbst, sondern andere Individuen begünstigen. Die Erklärung solches altruistischen Verhaltens stellt die Theorie seit Darwin selbst vor Probleme und wird bis heute intensiv in der Wissenschaft diskutiert[8][9][10]. Das Problem besteht auch dann, wenn das Verhalten letztlich allen, d.h. auch dem helfenden Individuum selbst, zugute kommt. Dies liegt daran, dass ein "Betrüger", der helfende Individuen nur ausnutzt, ohne selbst etwas beizutragen, immer eine höhere Fitness als ein Helfer besitzen müssste. Damit wäre Altruismus keine evolutionär stabile Strategie. Sobald durch eine Mutation ein einziger "Betrüger" in der Population entstehen würde, müsste er sich unweigerlich durchsetzen, selbst wenn am Ende alle schlechter dran wären.
Die Wissenschaft hat eine Vielzahl von Theorien aufgestellt, wie dieses Problem zu lösen sein könnte. Zu erklären ist dabei, worin der langfristige evolutionäre Vorteil der helfenden Strategie liegen kann (kurzfristig ist dies keine Frage, da auf kurze Sicht der Betrüger immer besser dran ist); man spricht von den "ultimaten" Gründen ("Warum"-Fragen). Andererseits müssen auch die Voraussetzungen und Mechanismen aufgeklärt werden, unter denen helfendes Verhalten entstehen kann, dies nennt man die "proximaten" Gründe ("Wie"-Fragen). Beide Betrachtungsebenen müssen streng getrennt werden, weil es sonst unweigerlich zu Konfusionen kommt.
Klassische Gruppenselektion
Der Evolutionsbiologe Vero Wynne-Edwards ist der Begründer einer Theorie, nach der die Evolution helfenden Verhaltens durch die Selektion nicht zwischen Individuen, sondern zwischen Gruppen von diesen zu erklären sei[11]. Nach seiner Hypothese besitzen Gruppen, bei denen die Individuen aufeinander Rücksicht nehmen, insgesamt eine höhere Fitness als Gruppen aus Individuen, bei denen jeder ausschließlich sein eigenes Interesse sucht. Solche "Egoisten"-Gruppen sterben häufiger als Ganzes aus, z.B. weil sie ihre Fortpflanzungsrate nicht an die Tragfähigkeit ihres Lebensraums anpassen ,diesen übernutzen und dann daran zugrunde gehen. Die kooperierenden Gruppen bleiben übrig. So setzt sich das Merkmal "Kooperation" schließlich insgesamt durch.
Innerhalb der Evolutionstheorie wird Wynne-Edwards Theorie der Gruppenselektion heute überwiegend abgelehnt. Theoretiker haben nachweisen können, dass der Selektionsmechanismus, den Wynne-Edwards vorschlägt, durchaus funktionieren würde. Allerdings sind dazu sehr enge Rahmenbedingungen erforderlich, die in natürlichen Populationen kaum denkbar erscheinen. Zum Beispiel macht bereits sehr moderater Austausch von Individuen zwischen Gruppen (und dadurch bewirkter Genfluss) den Mechanismus zunichte. Es ist auch nur in wenigen Fällen gelungen, natürliche Populationen ausfindig zu machen, bei denen der Mechanismus plausibel wäre. Meist kann man sie mit einer der konkurrierenden Theorien besser erklären.
Verwandtenselektion
Die Theorie der Verwandtenselektion (engl.: kin selection) geht auf die Biologen John Maynard Smith und William D. Hamilton zurück. Nach dieser Theorie wird helfendes Verhalten nur dann evolutionär begünstigt, wenn die Individuen, denen geholfen wird, ebenfalls Träger des "Helfer-Gens" sind, welches das Hilfsverhalten genetisch determiniert. In diesem Fall erhöht sich die Genfrequenz des Helfergens in der Folgegeneration nicht nur durch die Nachkommen des Helfers selbst, sondern auch durch diejenigen der Geholfenen. Dadurch kann dieses Gen sich in der Population letztlich durchsetzen. Bei der Anwendung der Hypothese muss man den Begriff der Fitness umdefinieren, da sich der entscheidende Selektionsvorteil nicht beim Vergleich verschiedener Individuen erweist, sondern in sehr vielen Individuen jeweils zum Teil verwirklicht ist, für dies hat sich der Ausdruck "inklusive Fitness" eingebürgert. Die Theorie der Verwandtenselektion legt es deshalb nahe, den Vorteil nicht der beteiligten Individuen, sondern ihrer jeweiligen Gene ins Zentrum der Betrachtung zu stellen. In besonders prägnater Form hat das der Biologe Richard Dawkins mit seiner Theorie vom "egoistischen Gen", durchgeführt, die in wesentlichen Teilen auf das Konzept der Verwandtenselektion zurückgeht.
Die Frequenz des Helfergens kann sich dann besonders leicht in der Population erhöhen, wenn Helfer ausschließlich Verwandten helfen (da diese ja einen großen Anteil ihrer Gene mit dem Helfer zwangsläufig gemeinsam haben), im Idealfall noch abgestuft nach dem Grad der Verwandtschaft (gemäß Hamiltons Regel). Dies ist die Verwandtenselektion im engeren Sinne. Daneben ist aber auch eine Verwandtenselektion im weiteren Sinne zu berücksichtigen. Dabei kann der Helfer z.B. allen benachbarten Individuen gleichermaßen helfen, wenn sich Individuen der Art nur selten und wenig weit ausbreiten - in diesem Fall ist jeder Nachbar mit genügend hoher Wahrscheinlichkeit ein Verwandter, ohne dass dies dem Helfer bekannt sein müsste. Daneben ist es theoretisch durchaus plausibel, dass nach demselben Mechanismus Träger des Helfergens sich einfach gegenseitig helfen, ohne dass ihre Verwandtschaft bekannt sein müsste, wenn sie sich gegenseitig erkennen können. Nach einem frühen Gedankenexperiment könnten sie z.B. alle einen grünen Bart tragen. Danach ist der Mechanismus als "Grüner-Bart-Effekt" in die Wissenschaft eingegangen.
Reziprokität
Vor allem für das Studium des menschlichen Verhaltens sind Modelle entwickelt worden, die auf der Spieltheorie beruhen, in besonderer Weise auf dem sog. Gefangenendilemma. In die Biologie eingeführt wurde das Konzept durch Robert Trivers [12]. Demnach könnte sich eine Disposition für Helfen in einer Population auch zwischen nicht verwandten Individuen einfach dadurch durchsetzen, dass der Helfer nur denjenigen hift, von denen er im Gegenzug selbst Hilfe erhält (Hilfe auf Gegenseitigkeit, nach dem Fachausdruck Reziprokität). Für diese Hypothese sind Sanktionen und Bestrafungen besonders wichtig, durch die ein Helfer diejenigen bestraft, die die Hilfe nicht durch Gegenhilfe "zurückzahlen". Im Gegensatz zu vielen biologischen Ansätzen stehen im Zentrum des Interesses hier nicht Verhaltensweisen selbst (die unterschiedlich determiniert oder motiviert sein können), sondern dadurch vermittelte Interaktionen, die in Form einer Interaktionsmatrix miteinander verknüpft werden, ohne dass die Motivation des Akteurs bekannt sein müsste oder auch nur eine besondere Rolle spielt.
Das Prinzip der Reziprokität scheint außerhalb der Art Mensch nach empirischen Studien allerdings nur sehr wenig verbreitet zu sein, so dass seine Erklärungskraft heute trotz theoretischer Plausibilität meist als gering eingeschätzt wird.
Neue Gruppenselektion und Multilevel-Selektion
Eine Gruppe von Evolutionsbiologen hat das Konzept der Gruppenselektion mit gegenüber der ursprünglichen Fassung etwas veränderter Definition erneut in die Debatte eingebracht, vor allem, um Fälle erklären zu können, bei denen aus ihrer Sicht das Modell der Verwandten-Selektion unzureichend ist. Prominenteste Vertreter dieser Theorie ist David Sloan Wilson [13]. (Sein bekannterer Namensvetter Edward Osborne Wilson hat sich der Theorie angeschlossen[14].) Nach der Theorie wirkt die natürliche Selektion auf verschiedenen Ebenen (Leveln) gleichzeitig: das Individuum, die soziale Gruppe und die Population können demnach gleichermaßen Selektionslevel darstellen. Im Gegensatz zur klassischen Gruppenselektion (s.o.) wirkt dabei die Selektion aber vor allem innerhalb der Population, nicht so sehr durch Selektion ganzer Populationen gegeneinander. Der Theorie gemäß kann man die Gesamtfitness in einen Anteil aufteilen, der zwischen den Individuen einer Population wirkt, und einen anderen, der den relativen Selektionsvorteil zweier Populationen gegeneinander beschreibt. Der Selektionswert für das einzelne Individuum ergibt sich demnach über die Summe der Selektionswerte auf den unterschiedlichen Leveln.
Verschiedene mathematisch arbeitende Theoretiker haben gezeigt, dass die Theorien der Verwandtenselektion und diejenigen der Multilvel-Selektion zu ganz überwiegenden Teilen lediglich verschiedene Wege darstellen, denselben Sachverhalt konzeptionell zu fassen. Das bedeutet, dass man die zugrunde liegenden mathematischen Modelle ineinander überführen kann. Die Multilevel-Selektion ist demnach eine alternative Fassung der Verwandtenselektion (im weiteren Sinne!)[15].
Ökologischer Ansatz
Für die Ökologie besteht zwischen Akteuren, die derselben Art angehören, und Interaktionen zwischen Akteuren verschiedener Spezies kein prinzipieller Unterschied, wodurch sich der Blickwinkel erweitert. Hilft ein Individuum einem anderen, das zu einer anderen Art gehört, in beiderseitigem Interesse, entspricht dies der Definition von Mutualismus, der damit nicht von der Kooperation zwischen Artgenossen prinzipiell verschieden ist.
Im Zentrum der ökologischen Betrachtung stehen die Mechanismen, die die (auf andere Weise begründeten) Vorteile einer kooperativen Strategie realisieren können. Betrachtet werden z.B. in Gruppen zusammenlebende Arten im Vergleich mit ihren einzeln lebenden Verwandten. Die Forschung hat Bedingungen identifizieren können, unter denen die Bildung von Gruppen ökologisch vorteilhaft ist. Besteht z.B. ein einfacher Zusammenhang zwischen Gruppengröße und Erfolg in aggressiven Begegnungen, ist es vorteilhaft, auch Nicht-Verwandte in die Gruppe aufzunehmen[16]. Unter diesen Bedingungen ist es vorteilhaft für den Einzelnen, in den Zusammenhalt der Gruppe zu investieren. Allerdings müssen dann (genetische und/oder soziale) Mechanismen zum Ausschluss von Betrügern entwickelt werden. Je nach Umweltbedingungen kann es sich aber unterschiedlich auszahlen, zu kooperieren oder nicht[17].
Wirkungsweise der Selektion
Die Selektion kann sich auf jedes erbliche Merkmal eines Individuums auswirken, das, direkt oder indirekt, Auswirkungen auf dessen Fortpflanzungsrate haben kann. Durch die Wirkung der Selektion optimierte Merkmale nennt man Adaptationen.
Sobald sich Individuen unterscheiden, ist grundsätzlich immer eine Selektion zwischen ihnen wirksam. Wenn kein anderer Faktor begrenzend wirkt, richtet sich diese auf die Fortpflanzungsrate selbst. In diesem Fall wird auf diejenigen Individuen selektiert, die die meisten Nachkommen produzieren können. Normalerweise wird das Wachstum aber über kurz oder lang immer durch Konkurrenz um Ressourcen begrenzt sein oder durch andere antagonistische Beziehungen wie z.B. Prädation beschränkt werden. Die Selektion wirkt sich dann auf diejenigen Merkmale am stärksten aus, die am stärksten wachstumsbegrenzend wirken. Dadurch können andere Merkmale. zumindest vorübergehend, von der Selektion weitestgehend ausgenommen bleiben.
Damit die Selektion Evolution bewirken kann, muss das selektierte Merkmal eine genetische Variabilität besitzen. Fehlt diese, führt auch starker Selektionsdruck nicht zur Evolution. In der Regel bewirkt die Selektion immer eine Verkleinerung der Variabilität (es gibt Ausnahmen, vor allem die disruptive Selektion. vgl. u.). Starke Selektion führt in evolutionär kurzer Zeit zwangsläufig dazu, dass nach und nach alle Allele mit Auswirkungen auf das selektierte Merkmal fixiert werden (d.h. entweder in allen oder in keinem Individuum vorkommen), wodurch eine weitere Veränderung des Merkmals immer schwerer wird.
Da Individuen einer Population in der Regel untereinander zwangsläufig recht ähnliche Bedürfnisse haben und einem gleich gerichteten Selektionsdruck unterliegen werden, ist der entscheidende Faktor die Konkurrenz zwischen Individuen derselben Art (genannt: intraspezifische Konkurrenz). Die Artgenossen sind allerdings bei sexuell fortpflanzenden Arten nicht nur Konkurrenten, sondern auch Paarungspartner. Durch die Vermischung des Erbguts bei der Fortpflanzung (Rekombination) können bei verschiedenen Individuen innerhalb der Population erworbene Adapatationen miteinander kombiniert werden und so die Fitness schneller erhöhen. Zunächst wirkt die Durchmischung aber (als Genfluss) homogenisierend und damit einer sehr schnellen selektiven Anpassung entgegen. Wichtigster Effekt ist aber, dass sich die Variabilität erhöht, wodurch die Selektion immer neue Ansatzpunkte bekommt.
Die drei bekannten Formen der Selektion – natürliche, sexuelle und künstliche Selektion – können jeweils in drei Typen auftreten: als stabilisierende, gerichtete oder disruptive Selektion.
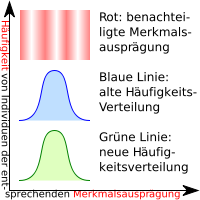
Rechts befindet sich eine Legende für die Grafiken, die die einzelnen Selektionsarten (Selektionstypen) illustrieren.
Stabilisierende Selektion
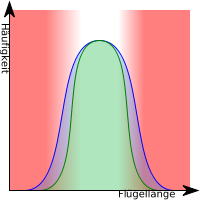
Stabilisierende Selektion (oder selektive Stabilisation/Stabilisierung bzw. selektive Retention) findet statt, wenn die Individuen einer Population über viele Generationen hinweg unter konstanten Umweltbedingungen leben. Individuen, deren Merkmale nahe am Mittelwert der Population liegen, zeigen eine höhere Fitness. Extreme bzw. vom Mittelwert abweichende Phänotypen können sich nicht durchsetzen. Somit führt stabilisierende Selektion zu einer geringeren phänotypischen Variabilität.
Ein Beispiel ist die Flügelgestaltung einiger Vogelarten. Längere oder kürzere Flügel haben schlechtere aerodynamische Eigenschaften als die mit Ideallänge, was zu Nachteilen bei Futterbeschaffung und Fluchtgeschwindigkeit führt.
Transformierende oder direktionale Selektion
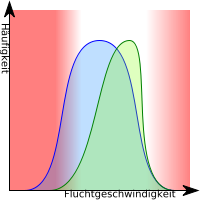
Transformierende, dynamische, direktionale, verschiebende oder gerichtete Selektion liegt vor, wenn die Träger eines Merkmals, das am Rand des Merkmalsspektrums der Population liegt, begünstigt werden. Muss sich z. B. eine Population an neue Umweltfaktoren anpassen, werden Individuen bevorzugt, deren Merkmale bereits vorher zufällig am besten auf die veränderte Umgebung gepasst haben (Präadaption) und/oder die sich besser an die neuen Bedingungen anpassen. Dies führt zu einer Veränderung des Genpools. Eine sehr starke direktionale Selektion kommt durch gezielte Züchtung zustande.
- Eine Population zeigt bestimmte Variabilität in der Ausprägung eines bestimmten Merkmals.
- Extreme Merkmalsausprägung, durch Selektionsdruck aufgrund veränderter Umweltbedingungen
- Eine Merkmalsausprägung bekommt Selektionsvorteile gegenüber dem anderen Extrem.
- Ständige Anpassung an sich ändernde Umweltbedingungen.
Zum Beispiel haben Kleintiere mit höherer Fluchtgeschwindigkeit bessere Überlebensquoten, was eine selektionsbedingte Geschwindigkeitssteigerung zur Folge haben kann .
Disruptive Selektion
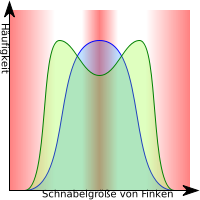
Bei der disruptiven (aufspaltenden) Selektion werden die Formen, die am häufigsten vorkommen, zurückgedrängt, z. B. aufgrund von Parasiten, Fressfeinden oder ansteckenden Krankheiten. Individuen, die seltene Merkmale besitzen, haben dann einen Vorteil (zum Beispiel die besonders kleinen und die besonders großen Individuen). Diese Individuen können durch ihre spezifischen Merkmale sog. ökologische Nischen besetzen, was Ihnen einen evolutionären Vorteil, beispielsweise bei der Nahrungsbeschaffung, bringen kann. Sie sind selektionsbegünstigt. Der Selektionsdruck sorgt für eine geringere Häufigkeit der Tiere mit „durchschnittlichen“ Merkmalen; diejenigen mit den extremen Phänotypen sind selektionsbegünstigt. Man spricht hier auch von Polymorphismus.
Der Fachbegriff für die Besetzung der ökologischen Nischen bei neuer Artenbildung durch disruptive Selektion lautet adaptive Radiation. Eine solche disruptive Selektion kann zu einer bimodalen Häufigkeitsverteilung führen und hierdurch zu einer Aufspaltung der Populationen in letztlich zwei getrennte Arten.
Ein klassisches Beispiel sind die sog. Darwinfinken deren selektionsbedingte Artaufspaltung bereits von zahlreichen Evolutionsforschern untersucht wurde.
Selektion auf Genebene
Für die Wirkung der Selektion auf der Ebene der Gene selbst ist eine andere Einteilung gebräuchlich. Selektion auf Genebene bedeutet in anderen Worten Änderung der Allelfrequenz. Hierbei können verschiedene Prozesse ablaufen, die jeweils in unterschiedlichen Situationen bedeutsam sind.
Zunächst sind prinzipiell zwei Wege zu unterscheiden, wie sich Selektion auf die Allelfrequenz auswirken kann:
- negative Selektion (auch: reinigende Selektion; engl.: purifying selection). Sie besteht in der Entfernung nachteiliger Allele. Die Rolle der negativen Selektion wird vor allem im Zusammenhang mit der neutralen Theorie der molekularen Evolution deutlich. Dieser (heute weitgehend anerkannten) Theorie zufolge ist der Einfluss der Gendrift in kleinen und mittelgroßen Populationen so groß, dass die zahlreich auftretenden neutralen Mutationen (d.h. diejenigen ohne Auswirkung auf die Fitness des Phänotyps, häufig sogar ohne Auswirkungen irgendeiner Art) häufig durch Zufall fixiert werden. Diese Mutationen unterliegen also nicht der Selektion. Die Selektion macht sich demnach vor allem durch die Auslese schädlicher Mutationen, d.h. negative Selektion, bemerkbar. Die Theorie leugnet nicht die Bedeutung positiver Selektion, sagt aber voraus, dass sie im Verhältnis viel seltener ist.
- positive Selektion. Sie bewirkt die Auswahl bestimmter Allele.
Positive Selektion tritt im Wesentlichen in zwei Formen auf:
- gerichtete Selektion (engl.: directional selection). Bei dieser wird ein Allel ausgelesen und damit häufiger. Dies führt letztlich zur Fixierung des Allels. Gerichtete Selektion vermindert also die Variabilität, auf Genebene spricht man von Polymorphismus. Zum Nachweis zurückliegender gerichteter Selektion auf Genebene sind indirekte Methoden etabliert. So wird z.B. bei Punktmutationen die Rate von nicht synonymen Substitutionen mit derjenigen von synonymen verglichen (durch die Redundanz des genetischen Codes führen nur bestimmte Nukleotid-Substitutionen zur Veränderung der Aminosäurensequenz im Protein, andere führen zu synonymen Sequenzen und sind damit "stumm"). Bei gerichteter Selektion sollte diese höher liegen als nach dem Zufall zu erwarten. Eine andere Methode versucht, sich den Effekt der gerichteten Selektion auf den Polymorphismus von Allelen zunutze zu machen. Vergleicht man den Polymorphismus im Genom an verschiedenen Loci, sollte er in Bereichen, die starker gerichteter Selektion unterliegen, geringer sein. Dieser Effekt wird "genetic sweep" (in etwa: genetisches Auswischen) genannt. Genetic sweep führt zu auffallend geringem Polymorphismus in Abschnitten der DNA unterschiedlicher Länge, die das selektierte Gen enthalten. Der Abschnitt ist gewöhnlich länger als das Gen selbst, weil die Selektion nicht am Gen selbst ansetzen kann, sondern nur an einem zufällig durch Rekombinationsvorgänge begrenzten DNA-Abschnitt, der das Gen enthält.
- ausgleichende Selektion (engl.: balancing selection). Bei dieser wird ein Allel je nach seiner Frequenz unterschiedlich ausgelesen. Wenn es selten ist, wird es selektiv bevorzugt. Wird es häufiger, wird es hingegen selektiv benachteiligt. Ausgleichende Selektion erhält also den Polymorphismus. Für das Wirken ausgleichender Selektion sind im Wesentlichen zwei Prozesse bedeutsam: Einerseits können auf bestimmten Genloci heterozygote Individuen gegenüber homozygoten bevorzugt sein (engl.: overdominance). Ein wichtiges Beispiel dafür ist die Evolution des Immunsystems der Wirbeltiere. Für die Vielfalt der Antikörper, die die Erkennung beinahe jedes fremden Organismus ermöglichen, sind vor allem die MHC-Gene verantwortlich. Heterozygote Individuen besitzen logischerweise mehr MHC-Gene als homozygote. Sie haben ein leistungsfähigeres Immunsystem und sind daher bei der Selektion bevorzugt. Der Vorteil liegt nicht darin, dass einzelne MHC-Gene "besser" oder "schlechter" wären, gefördert wird einfach ihre Vielfalt. Dadurch werden seltene Gensequenzen von der Selektion gefördert[18]. Ein weiteres Beispiel mit ähnlicher Grundlage ist die Evolution der Selbstinkompatibilität bei Pflanzen. Derselbe Effekt kann auch dadurch eintreten, dass ein Allel im heterozygoten Fall günstige Eigenschaften vermittelt, welches bei Homozygotie schädlich oder gar lethal ist. Berühmt geworden ist dieser Fall bei der Sichelzellenanämie, einer menschlichen Erbkrankheit, deren Gen im heterozygoten Fall Resistenz gegenüber Malaria bewirkt und dadurch in Malariagebieten selektiv in evolutionär kurzer Zeit stark gefördert worden ist. Andererseits kann ein seltenes Allel gelegentlich auch direkt bevorzugt sein, z.B. wenn Prädatoren in der Umwelt häufige Beute gegenüber seltener bevorzugen. Ein Beispiel, bei dem seltene Merkmalskombinationen dadurch direkt gefördert wurden, zeigte sich beim Guppy. Bei dieser Fischart existieren vor allem von den Männchen zahlreiche, extrem verschiedene Farbmorphen. Es konnte nachgewiesen werden, dass Männchen mit selteneren Farbmustern seltener von Räubern gefressen werden[19] Daneben unterliegt dasselbe Merkmal allerdings auch noch der Wirkung der sexuellen Selektion. Bei vielen Tierarten bevorzugen die Weibchen gezielt Männchen mit ungewöhnlichen und seltenen Merkmalen gegenüber vertrauten Formen, beim Guppy werden z.B. Männchen mit seltenen Farbmustern signifikant als Paarungspartner bevorzugt[20]. Dieser Effekt kann so weit gehen, dass Individuen mit künstlichen, vom Menschen angebrachten Markierungen (z.B. für verhaltensbiologische Experimente) größere Paarungschancen besitzen, wie bei Zebrafinken beobachtet[21].
Auswirkungen anderer Variationen auf Genebene
Mutationen in der proteincodierenden Sequenz von Genen und dadurch erzeugte Allele sind der am besten untersuchte Mechanismus, der Variationen bereitstellt, auf deren Existenz das Wirken der Selektion bei der Gestaltung von Adaptationen angewiesen ist. Es gibt allerdings umfangreiche theoretische wie empirische Hinweise auf weitere Mechanismen. Wichtige Beispiele sind:
- Mutationen in genregulierenden Sequenzen
- Genduplikation (durch Mutationen bei der Rekombination oder Meiose) mit Wandel der Funktion einer der Kopien
- Genrekrutierung: Übernahme neuer Funktionen für existierende Gene
- Exon shuffling
- Einfügung von Transposons in bestehende Gene.
Eine Übersicht über diese Fälle gibt[22]. Sogar eine völlige Neuentstehung von Genen aus funktionsloser DNA erscheint (wenn auch sehr selten) möglich. Neuere Evolutionstheorien wie die evolutionäre Entwicklungsbiologie stellen diese Vorgänge ins Zentrum ihres Interesses, auch wenn die empirischen Belege nach wie vor umstritten sind[23]. Auch wenn die besprochenen Fälle möglicherweise seltener auftreten (oder zumindest weniger gut erforscht sind) als Mutationen der proteincodierenden Sequenzen, sind ihre möglichen Auswirkungen auf die Evolution beträchtlich. Einige Forscher haben daraus die Folgerung abgeleitet, dass das Wirken der Selektion als entscheidender Triebkraft der Evolution möglicherweise überschätzt worden ist. Das Wirken der Selektion auf solche seltenen Ereignisse ist viel schwieriger im Labor zu erforschen als das der recht gut verstandenen Auswirkungen "klassischer" Mutationen. Trotz ihrer möglicherweise großen Bedeutung für die Evolution liegen daher noch keine abgeschlossenen Theorien vor.
Selektion messen
Es ist im Prinzip möglich, die Wirkung der Selektion in einer Population direkt zu messen[24]. Folgendes ist für eine Messung erforderlich:
- ein Merkmal, das innerhalb der Population variiert, von dem ein selektiver Effekt angenommen wird
- der Effekt dieses Merkmals auf die Fitness. Das Maß für die Fitness ist, wie immer, die jeweilige Fortpflanzungsrate. Notwendig ist also, den Fortpflanzungserfolg der Individuen zu messen.
Trägt man die Variation des Merkmals gegenüber dem Fortpflanzungserfolg in einem Graphen auf, können sich folgende Zusammenhänge ergeben:
- keine Selektion wirksam: Es besteht kein Zusammenhang zwischen dem gemessenen Merkmal und dem Fortpflanzungserfolg. Der Graph ergibt eine Gerade parallel zur X-Achse (real durch Messfehler und natürliche Variabilität nicht perfekt verwirklicht).
- gerichtete Selektion wirksam: Der Fortpflanzungserfolg steigt linear mit dem Merkmal an (oder fällt linear ab). Es ergibt sich im Graphen eine Gerade mit einer Steigung, die als Maß für die Stärke der Selektion dienen kann.
Ist eine der anderen Formen der Selektion wirksam, besteht zwischen Merkmal und Fortpflanzungserfolg kein linearer Zusammenhang mehr.
- bei stabilisierender Selektion ergibt sich eine Kurve, die bis zu einem bestimmtem Maximum ansteigt (der optimalen Merkmalsausprägung), um danach wieder abzufallen.
- bei disruptiver Selektion ergibt sich eine Kurve, die bis zu einem Minimum abfällt, um danach wieder anzusteigen.
Beide Formen ergeben also ähnliche Verteilungen, wobei nur das Vorzeichen vertauscht ist. Gegenüber der linear wirkenden gerichteten Selektion sind zur mathematischen Beschreibung der Kurve höherdimensionale (Quadratische) Terme erforderlich.
Obwohl also die Messung der Selektion vom Prinzip her zunächst relativ einfach erscheint, ergeben sich in der Praxis zahlreiche Fallstricke, durch die die tatsächliche Messung zu einer anspruchsvollen Aufgabe wird. Zunächst ist es in natürlichen Populationen extrem schwierig und (je nach Lebensdauer der untersuchten Art) auch zeitaufwendig, die reale Fortpflanzungsrate über die gesamte Lebensdauer zu messen. Über kürzere Fristen zu messen ist nicht zulässig, weil der Fortpflanzungserfolg bei beinahe jeder Art bekanntermaßen vom Lebensalter abhängig ist. Im Labor zu messen ist zwar einfacher, aber es ist normalerweise nicht zulässig, die Ergebnisse auf das Freiland zu übertragen, da die Fitness von der jeweiligen Umwelt abhängig ist. Um das Problem zu umgehen, wird häufig ein leichter messbares Kriterium verwendet, welches im untersuchten Fall einen wesentlichen Anteil der Fitness erklären kann, z.B. die Überlebensrate. Ein noch größeres Problem besteht darin, dass das gemessene Merkmal möglicherweise gar nicht direkt für den beobachteten Effekt kausal verantwortlich ist, sondern es mit dem eigentlich entscheidenden Merkmal korreliert. In der Praxis ist beinahe immer davon auszugehen, dass die Selektion auf zahlreiche Merkmale gleichzeitig einwirkt und das ein bestimmter Effekt von zahlreichen Merkmalen jeweils zum Teil verursacht wird. Um den Anteil des betrachteten Merkmals herauszulösen, ist dessen partielle Regression zu bestimmen[25]
Inzwischen ist bei zahlreichen natürlichen Populationen nach der beschriebenen Methode versucht worden, die Stärke der einwirkenden Selektion zu messen[26]. Die dabei erzielten Resultate sind aber in den meisten Fällen noch nicht aussagekräftig genug, weil die Größe der gemessenen Populationen noch zu klein war[27]. Zusätzlich zeigte sich, dass die Stärke der Selektion scheinbar in den größeren Studien immer kleiner wurde. Dies deutet auf ein Publikations-Vorurteil ("Schubladen-Effekt"), weil Studien, die, vielleicht nur zufällig, einen signifikanten Zusammenhang zwischen Effekt und Merkmal zu belegen scheinen, häufiger publiziert und zur Publikation angenommen werden als solche ohne diesen, wodurch das Resultat in der Wahrnehmung verzerrt wird.
Selektion in nicht lebenden Systemen
Manfred Eigen hat in seinen Untersuchungen zur Selbstorganisation als Ursprung des Lebens das Konzept der Quasispezies entwickelt. Dabei hat er das Konzept der Selektion auf chemische Systeme auf der Früherde vor Entstehung des Lebens übertragen[28]. Nach Eigens Theorie ist eine Evolution unausweichlich, sobald autokatalytische Makromoleküle und eine Energiequelle vorhanden sind. Selektion ist für ihn ein aus physikalischen Größen ableitbares Optimalprinzip.
Literatur
- Douglas J. Futuyma (1990): Evolutionsbiologie. Springer Verlag. ISBN 3764322004
- Charles Darwin: Die Entstehung der Arten
- Richard Dawkins (1992): Der blinde Uhrmacher Kindler Verlag. ISBN 3463400782. orig: (1986) The blind Watchmaker.
- Ernst Mayr (1994): ...und Darwin hat doch recht Piper: München. orig. (1991): One Long Argument: Charles Darwin and the Genesis of Modern Evolutionary Thought.
- George C. Williams (1996): Adapatation and natural selection. Princeton University Press. ISBN 0691026157
Siehe auch
Weblinks
- Selektion und Fitness. Definitionen und theoretische Grundlagen
- Arten biologischer Selektion
- Robert Brandon: Natural Selection, in: Stanford Encyclopedia of Philosophy (englisch, inklusive Literaturangaben)
- Link zu einem Artikel von Ulf Dieckmann, siehe auch „Rezente Beispiele für künstliche Selektion“ / „Unexpected Discontinuities in Life-History Evolution under Size-dependent Mortality“: [2]
Einzelnachweise
- ↑ Richard Law (2000): Fishing, selection, and phenotypic evolution. ICES Journal of Marine Science, 57: 659–668.
- ↑ Esben Moland Olsen, George R. Lilly, Mikko Heino, M. Joanne Morgan, John Brattey, Ulf Dieckmann (2005): Assessing changes in age and size at maturation in collapsing populations of Atlantic cod (Gadus morhua). Canadian Journal of Fishery and Aquatic Sciences 62: 811–823.
- ↑ David O. Conover & Stephan B. Munch (2002): Sustaining fisheries yields over evolutionary time scales. Science Vol. 297 no. 5578: 94-96.
- ↑ David N Reznick & Cameron K Ghalambor (2005): Can commercial fishing cause evolution? Answers from guppies (Poecilia reticulata). Canadian Journal of Fisheries and Aquatic Sciences 62(4): 791-801.
- ↑ Eine Übersicht der Debatte in E. Lloyd: Units and Levels of Selection. The Stanford Encyclopedia of Philosophy (Winter 2010 Edition), Edward N. Zalta (Hrsg.) [1]
- ↑ Samir Okasha (2006): Evolution and the Levels of Selection. Oxford University Press. ISBN 978-0-19-926797-2.
- ↑ E. Mayr (1997): The objects of selection. Proceedings of the National Academy of Science USA 94: 2091–2094.
- ↑ S. A. West, A. S. Griffin, A. Gardner (2007): Social semantics: altruism, cooperation, mutualism, strong reciprocity and group selection. Journal of Evolutionary Biology 20(2: 415-32
- ↑ R. Bshary & R. Bergmüller (2008): Distinguishing four fundamental approaches to the evolution of helping. Journal of Evolutionary Biology 21: 405–420.
- ↑ S. F. Brosnan & R. Bshary (2010): Cooperation and deception: from evolution to mechanisms. Philosophical Transactions of the Royal Society Series B 365: 2593-2598.
- ↑ V. C. Wynne-Edwards (1962): Animal Dispersion in Relation to Social Behaviour. Oliver and Boyd, (Edinburgh).
- ↑ R. L. Trivers (1971): The evolution of reciprocal altruism. Quarterly Revue of Biology 46: 35–57.
- ↑ D. S. Wilson (1975): A theory of group selection. Proceedings of the National Acadademy of Science USA 72: 143–146.
- ↑ D. S. Wilson & E. O. Wilson (2007): Rethinking the theoretical foundations of sociobiology. Quarterly Revue of Biology 82: 327-348.
- ↑ Eine klare Herleitung (auf Basis der Price-Gleichung) z.B. in A. Gardner, S. A. West, N. H. Barton (2007): The Relation between Multilocus Population Genetics and Social Evolution Theory. American Naturalist 169(2): 207-226 + 2 app.
- ↑ T. Clutton-Brock (2009): Cooperation between non-kin in animal societies. Nature 462: 51-57.
- ↑ vgl. z. B.: J. Field, A. Cronin, C. Bridge (2006): Future fitness and helping in social queues. In: Nature, Band 441, S. 214–217.
- ↑ P. C. Doherty & R. M. Zinkernagel (1975): Enhanced immunological surveillance in mice heterozygous at the H-2 gene complex. Nature 256: 50–52.
- ↑ R. Olendorf, F. H. Rodd, D. Punzalan, A. E. Houde, C. Hurt, D. N. Reznick, K. A. Hughes (2006): Frequency-dependent survival in natural guppy populations. Nature 441: 633-636.
- ↑ K. A. Hughes, L. Du, F.H. Rodd, D. N. Reznick (1999): Familiarity leads to female mate preference for novel males in the guppy, Poecilia reticulata. Animal behaviour 58: 907-916.
- ↑ Nancy Burley(1988): Wild zebra finches have band-colour preferences. Animal behaviour 36(4):1235-1237.
- ↑ Manyuan Long, Esther Betrán, Kevin Thornton, Wen Wang (2007): The origin of new genes: glimpses from the young and old. Nature Reviews Genetics 4: 865-875.
- ↑ Hopi E. Hoekstra & Jerry A. Coyne (2007): The locus of evolution: evo devo and the genetics of adaptation. Evolution 61(5): 995–1016.
- ↑ Übersicht in: John A. Endler (1986): Natural selection in the wild. Princeton University Press (Monographs in Population Biology). ISBN 9780691083872.
- ↑ Russell Lande & Stevan J. Arnold (1983): The measurement of selection on correlated characters. Evolution 37(6): 1210-1226.
- ↑ ein Übersicht in: J. G. Kingsolver, H. E. Hoekstra, J. M. Hoekstra,, D. Berrigan, S. N. Vignieri, C. E. Hill,, A. Hoang, P. Gibert, P. Beerli (2001): The strength of phenotypic selection in natural populations. American Naturalist 157(3): 245-261.
- ↑ E. I. Hersch & P. C. Phillips (2004): Power and potential bias in field studies of natural selection. Evolution 58(3): 479–485.
- ↑ Manfred Eigen (1971): Selforganization of matter and the evolution of biological macromolecules. Naturwissenschaften 58(10): 465-523.




Wikimedia Foundation.