- Diakinese
-
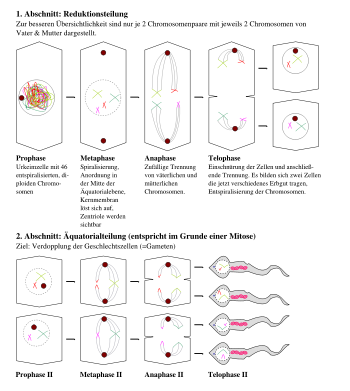 Meiose am Beispiel der Spermatogenese
Meiose am Beispiel der SpermatogeneseUnter Meiose (von griech.: meiono = vermindern, verkleinern), Reifeteilung oder Reduktionsteilung versteht man eine besondere Form der Zellkernteilung, wobei im Unterschied zur gewöhnlichen Kernteilung, der Mitose, die Anzahl der Chromosomen vom diploiden auf den haploiden Zustand reduziert wird. Dabei findet gewöhnlich auch eine Rekombination mütterlichen und väterlichen Erbguts statt. Die Meiose vollzieht sich immer in zwei Teilungsschritten (1. und 2. meiotische Teilung oder auch Meiose I und II genannt). In der Regel erfolgt nach jedem dieser Teilungsschritte eine Cytokinese (Zellteilung), was zur Bildung von 4 Einzelzellen führt, die als Keimzellen oder Gameten bezeichnet werden. Da die Zytokinese in der Regel mit der Meiose zusammenhängt, werden umgangssprachlich auch beide Vorgänge gemeinsam als Meiose bezeichnet.
Die Reduktion des Ploidiegrads (d. h. der Anzahl der Chromosomensätze) ist eine Voraussetzung für die geschlechtliche Fortpflanzung.
Inhaltsverzeichnis
Bedeutung
Die biologische Bedeutung oder der Sinn der Meiose wird darin gesehen, dass sie im Rahmen der sexuellen Fortpflanzung eine Rekombination des Erbguts mütterlichen und väterlichen Ursprungs ermöglicht. Das erhöht die genetische Vielfalt innerhalb der Population und damit die Anpassungsfähigkeit und ermöglicht eine schnellere Evolution im Vergleich zu asexuellen Organismen.
Die Rekombination wird dadurch möglich, dass durch eine Vereinigung von Zellen unterschiedlicher Herkunft verschiedene, aber homologe Gene in einem Zellkern zusammenkommen. Bei der Meiose werden die homologen Gene wieder voneinander getrennt, wobei es aber zu einem Austausch von Chromosomen und Chromosomenabschnitten zwischen den ursprünglichen haploiden Sätzen kommt. Das beruht erstens darauf, dass homologe Chromosomen zufällig den beiden Tochterzellen zugeteilt werden. Zweitens erfolgt bei den meisten Organismen (chiasmatische Meiose) im Verlauf der Meiose auch ein Austausch von Teilen homologer Chromosomen (Crossing over).
Der damit verbundene Wechsel zwischen einer haploiden und einer diploiden Phase (Kernphasenwechsel) tritt in mehreren Varianten auf. Bei den meisten höheren Organismen steht die diploide Phase ganz im Vordergrund. Nur die Gameten - oder im Falle der Blütenpflanzen die winzigen, wenigzelligen Pollen und Embryosäcke - sind haploid. Solche Organismen bezeichnet man als Diplonten. Den umgekehrten Fall repräsentieren viele Mikroorganismen, die normalerweise haploid sind und deren diploide Phase auf die Zygote beschränkt ist (Haplonten). Drittens gibt es Diplohaplonten, bei denen sich haploide und diploide Generationen abwechseln, z. B. Farne und Moose.
Phasen der Meiose
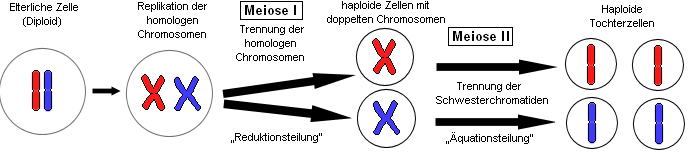 Vereinfachtes Schema der zwei Teilungsschritte der Meiose: die Reduktionsteilung und die Äquationsteilung. Das für die Rekombination wichtige Crossing over ist nicht dargestellt.
Vereinfachtes Schema der zwei Teilungsschritte der Meiose: die Reduktionsteilung und die Äquationsteilung. Das für die Rekombination wichtige Crossing over ist nicht dargestellt.Die Meiose läuft in zwei Teilschritten ab:
- Die Reduktionsteilung, auch 1. meiotische Teilung, 1. Reifeteilung oder einfach Meiose I. Hier wird der Chromosomensatz der diploiden Zelle reduziert, wobei die Chromosomen auf verschiedene Arten miteinander rekombiniert werden können. Schon nach der Reduktionsteilung weisen die Zellen nur noch einen einzigen Chromosomensatz auf, müssen aber noch eine Teilung durchlaufen. Dies liegt daran, dass die Chromosomen immer noch zwei Chromatiden haben, da sich ja nur der Chromosomensatz, nicht die Chromosomen selbst getrennt haben. So wird nach der Reduktionsteilung auch keine Replikation angestoßen und nach einer sehr kurzen Interphase folgt
- Die Äquationsteilung, auch als 2. meiotische Teilung, 2. Reifeteilung oder Meiose II bezeichnet. Diese Phase ähnelt nun einer normalen Mitose, nur dass hier ein haploider Chromosomensatz vorliegt. Ansonsten werden die Chromosomen in den normalen Zellkernteilungsphasen voneinander getrennt. Da die Meiose zwei Teilungsschritte durchläuft und jeder einzelne Schritt meist mit einer abschließenden Zytokinese zwei Tochterzellen entstehen lässt, liegen nach der abgeschlossenen Meiose vier haploide Tochterzellen vor.
Reduktionsteilung - Meiose I
Die 1. meiotische Teilung beginnt mit der Prophase I, die im Vergleich zur Prophase der Mitose stark verlängert ist: Sie kann über Wochen, Monate, oder im Falle der Oozyte beim Menschen sogar einige Dekaden andauern (Oozyte verweilt bis zur Ovulation im Diktyotän). Diese Phase wird in fünf Stadien unterteilt:
- Im Leptotän (von gr. leptos, dünn und lat. taenia, das Band) beginnen die Chromosomen sehr langsam zu kondensieren. Sie sind bis zum Ende der Prophase I an den Ärmchen-Enden, den so genannten Telomeren, an der inneren Zellkernmembran befestigt.
- Das Zygotän (von gr. zygon, das Joch) ist durch die Paarung der homologen Chromosomen gekennzeichnet. Hier liegt nun ein klar sichtbares Chromatin vor. Die Chromosomenpaarung, auch Synapsis, verläuft reißverschlussartig, wobei sich zwischen beiden Chromosomensträngen ein so genannter synaptonemaler Komplex bildet, der beide Stränge zusammenhält.
- Im Pachytän (von gr. pachys, derb) kommt es zu einer weiteren Kondensation der gepaarten Chromosomen. In dieser Phase findet im häufigsten Fall der chiasmatischen Meiose das Crossing-over statt, wodurch der Austausch homologer Chromatiden-Abschnitte zwischen den gepaarten Chromosomen eingeleitet wird.
- Im Diplotän (von gr. diploos, doppelt) werden die homologen Chromosomen voneinander getrennt, wobei die rekombinierten Stellen über so genannte Chiasmata noch miteinander verbunden bleiben. Bei einigen Organismen findet währenddessen noch Transkription statt. Jene Bereiche werden als Schleifen symmetrisch ausgelagert, weswegen man diese Chromosomen nach ihrem Aussehen als Lampenbürstenchromosomen bezeichnet. Die menschliche Oozyte verweilt hier in einer besonderen Phase, dem Diktyotän. Dieses Stadium kann bis zu ca. 50 Jahren andauern, da die Eizellen schon bei Geburt ausgebildet sind, sich aber erst kurz vor dem jeweiligen Eisprung weiter differenzieren.
- Mit der Diakinese (von gr. diakinein, auseinander bewegen) endet die RNA-Synthese und die volle Kondensation beginnt. Zudem löst sich nun die Zellkernhülle auf.
In der Prometaphase versammeln sich die gepaarten Chromosomen in der Äquatorialebene des Spindelapparats, womit die Metaphase I erreicht wird. Dabei können im Lichtmikroskop Überkreuzungen benachbarter Nichtschwesterchromatiden sichtbar werden, die Chiasmata. Die für die korrekte Anordnung und spätere Trennung (Segregation) der Chromosomen notwendigen Verbindungen mit den Spindelfasern (Kinetochore) werden dabei nur auf einer Seite der Schwesterchromatiden gebildet. Daher werden in der Anaphase komplette Chromosomen, nicht Chromatiden, zu den beiden Spindelpolen gezogen.
In der Telophase I liegt dann an jedem Pol jeweils nur noch ein Chromosom (mit zwei Chromatiden) jedes Typs. Es ist also zu einer Reduktion der Chromosomenzahl auf einen haploiden Satz gekommen.
Äquationsteilung - Meiose II
Die 2. meiotische Teilung läuft ähnlich wie die Mitose ab. Während der Teilungsphasen (Metaphase II, Anaphase II, Telophase II) werden die beiden Chromatiden jedes Chromosoms voneinander getrennt, so dass die resultierenden vier Keimzellen jeweils ein Chromosom jedes Typs erhalten.
Nach Abschluss der Meiose besitzen also alle Gameten den benötigten haploiden Chromosomensatz. Sie sind damit für den Befruchtungsvorgang vorbereitet.
Besonderheiten bei Säugetieren
Bei Frauen werden so genannte weibliche Ur-Keimzellen (bzw. Primärfollikel) nur kurz nach der Geburt gebildet (in den ersten 6 Wochen) und dann in den Eierstöcken gespeichert. Während der Pubertät bekommen die Primärfollikel eine weitere mehrschichtige Hülle (Epithelgewebe). Der so entstandene Sekundärfollikel wandelt sich dann durch Umlagerungen von Epithelgewebe zu einem Tertiärfollikel um, (während des Eisprungs in aktiver Form auch Graaf’scher Follikel genannt). Bei dieser Reifung wird ein Follikel einmal pro Menstruationszyklus durch Wachstum und v.a. Wassereinlagerungen bis zu 6 mm groß und gleichzeitig in die Lage versetzt auch Hormone (anfangs v.a Östrogen, nach dem Eisprung dann Progesteron) zu produzieren.
Kurz vor dem Eisprung beginnt nun erst die erste Reifeteilung. Während die Bildung von Spermien genau nach dem erklärten Muster verläuft, teilt sich die Eizelle bereits in der ersten Reifeteilung asymmetrisch in eine große, plasmahaltige Eizelle und eine deutlich kleinere: Das so genannte Polkörperchen. Bei der Bildung der Eizellen findet in der ersten Reifeteilung also eine ungleiche Verteilung des Zytoplasmas statt. Beide Zellen enthalten natürlich einen haploiden Chromosomensatz.
Die zweite Reifeteilung beginnt erst nach dem Ei- bzw. Follikelsprung. In der zweiten Reifeteilung teilt sich die Eizelle wiederum asymmetrisch, wodurch nun zwei Polkörperchen entstanden sind, die sich nicht mehr weiter teilen und später absterben. Sie wird durch Kontakt mit Spermien ausgelöst. Kommt es nicht zur Befruchtung, geht die Eizelle zugrunde.
Weiterhin findet man bei fast allen Säugetieren auch „provozierte Follikelsprünge“. Diese spontane Follikelabgabe (=Ovulation) kann durch Stress, Fieber, Nervosität, hormonelle Schwankungen, Schlafmangel usw. ausgelöst werden. Bei Hasen und Kaninchen kann der Begattungsakt selbst zur spontanen Ovulation reizen. Bei Katzen reicht ein mechanischer Reiz am Gebärmutterhals.
Literatur
- Bernard John, Jonathan B.L. Bard, Peter W. Barlow: Meiosis, Cambridge University Press, 2006
Weblinks

Wikimedia Foundation.