- Foraminifera
-
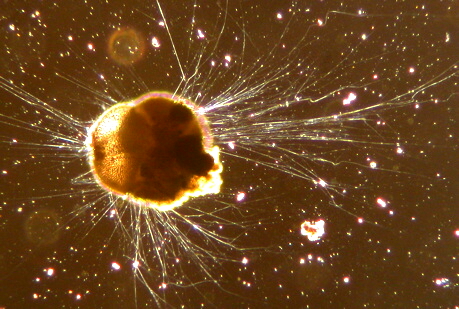
Foraminifera 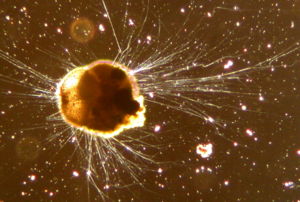
Ammonia tepida
Systematik Klassifikation: Lebewesen Domäne: Eukaryoten (Eucaryota) ohne Rang: Rhizaria ohne Rang: Foraminifera Wissenschaftlicher Name Foraminifera d'Orbigny, 1828 Die einzelligen Foraminiferen (Foraminifera), auch Kammerlinge genannt, sind ein Taxon von Amöben aus der Gruppe der Rhizaria. Sie umfasst rund 10.000 rezente und 40.000 fossil bekannte Arten und stellt damit die bei weitem größte Gruppe der Rhizaria dar.
Bis auf rund fünfzig im Süßwasser lebende Arten besiedeln alle Foraminiferen marine Lebensräume von den Küsten bis in die Tiefsee, zumeist auf dem Meeresboden (benthisch); ein kleiner Teil, die Globigerinida, zählt zum Plankton. Die Mehrheit aller Foraminiferen zeichnet sich durch ein Gehäuse aus. [1].
Die außerordentlich formenreiche Gruppe ist fossil seit dem Kambrium (rund 560 mya) nachgewiesen, molekulargenetische Untersuchungen (Molekulare Uhr) verweisen jedoch auf ein deutlich höheres Alter von 690 bis 1150 Millionen Jahren [2]. Foraminiferen dienen in der Paläontologie aufgrund ihrer gut fossil erhaltungsfähigen, oft gesteinsbildenden Schalen als Leitfossilien für Kreide, Paläogen und Neogen.
Inhaltsverzeichnis
Merkmale
Alle Arten der Foraminifera sind einzellige Lebewesen, die Durchmesser von 0,02 bis 0,05[3] Millimetern bis hin zu annähernd 20 Zentimetern und ein Alter von bis zu mehreren Monaten oder sogar einigen Jahren erreichen können [1]. Fast alle Arten besitzen ein in der Regel vielkammeriges Gehäuse. Durch dessen üblicherweise feine Perforation treten fadenförmig dünne Scheinfüßchen aus, die Pseudopodien. Die Foraminifera unterliegen einem Generationswechsel zwischen sexuellen und asexuellen Formen, ein Merkmal, das in einigen Gruppen aber modifiziert worden ist. [4]
Retikulopodien
Die Pseudopodien dienen der Verankerung im Boden sowie der Aufnahme von Teilchen des Sediments und ihrer Zusammensetzung zu einem Gehäuse. Darüber hinaus dienen sie der Sammlung von Nährstoffen, dem Fang von Beute und bei manchen Arten der extrathalamen Verdauung (extrathalam=außerhalb des Gehäuses) [4]. Sie sind fadenförmig dünn, verzweigen sich und verschmelzen wieder (Anastomosie) und bilden so ein Netzwerk sogenannter Retikulopodien, dass sich über mehrere hundert Quadratzentimeter ausbreitetn kann. An seinen jeweiligen Enden verjüngt es sich auf einen Durchmesser von weniger als einen Mikrometer. [1]
Die Retikulopodien werden durch einzeln wie auch bündelweise auftretende Mikrotubuli verfestigt, die auch Bewegungen vermitteln, darunter als Besonderheit der Foraminifera eine bidirektionale Strömung des Zytoplasmas innerhalb der Pseudopodien [1].
Aufbau der Gehäuse
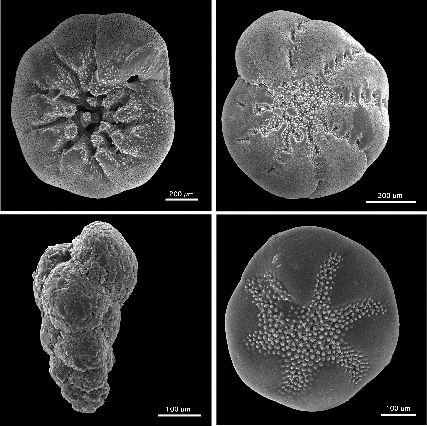
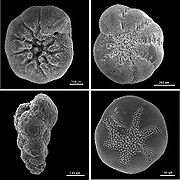 Schalen rezenter benthischer Foraminiferen aus Nordamerika, (im Uhrzeigersinn links oben beginnend: Ammonia beccarii, Elphidium excavatum clavatum, Buccella frigida, Eggerella advena, REM-Aufnahme, verschiedene Maßstäbe)
Schalen rezenter benthischer Foraminiferen aus Nordamerika, (im Uhrzeigersinn links oben beginnend: Ammonia beccarii, Elphidium excavatum clavatum, Buccella frigida, Eggerella advena, REM-Aufnahme, verschiedene Maßstäbe)Die Gehäuse der Foraminiferen können sehr verschieden gebaut sein, der Reichtum ihres Aufbaus dient bis in die Gegenwart als diagnostisches Merkmal zur Unterscheidung der Taxa. Gehäuse können aus nur einer Kammer bestehen (monothalam), aus einer Reihe von miteinander verbundenen Kammern (polythalam) oder -sehr selten- ganz fehlen (athalam). [1]
Als Baustoff für die Gehäuse dient entweder sekretorisch ausgeschiedener Kalk (zumeist Kalzit, selten Aragonit) oder Opal (SiO2 x n H2O) oder ein organisches Material namens Tektin, ein dem Keratin verwandtes Protein. Bei sogenannt agglutinierenden Foraminiferen werden Sandkörner oder andere Fremdkörper aus dem Sediment aufgenommen und miteinander verklebt.
Ökologie der Foraminiferen
Foraminiferen sind vorwiegend Bewohner des Meeres, nur ganz wenige Arten finden sich im Frischwasser. Im Meer leben sie entweder auf oder im Meeresboden (benthische Lebensweise) oder -seltener- frei in der Wassersäule schwebend (planktische Lebensweise). Am Boden können sie fest mit dem Substrat verbunden sein (sessilobenthisch) oder frei umherschweifen (vagilobenthisch).
Energiehaushalt
Foraminiferen decken ihren Energiebedarf auf vielfältige Weisen. Es gibt fleisch-, pflanzen- oder allesfressende Arten, die sich beispielsweise als Weidegänger, Destruenten, Filtrierer, Parasiten oder über photosynthetisierende Endosymbionten ernähren. Dessen ungeachtet sind viele Arten in ihrer Ernährung auf einzelne Beutegruppen spezialisiert. [4]
Arten, die in Tiefen leben, die noch vom Licht erreicht werden, leben meist von wenigen, ausgewählten Algen, ergänzt durch Bakterien, Hefen, Pilze und kleine Tiere. Lichtlos lebende Arten hingegen fungieren als Zersetzer des Detritus insbesondere phytoplanktonischen Ursprungs, Arten der Tiefsee leben dabei von bereits stärker zersetztem Material. [4]
Endosymbiontische Photosynthese
In den Scheinfüßchen sowie im Zytoplasma planktonischer wie auch großer benthischer Foraminiferen der Tropen finden sich pro Individuum bis zu dreitausend Zoochlorellae bzw. Zooxanthellae als Photosynthese betreibende Endosymbionten, die äußerstenfalls 75 bis 80%[5] des Gehäusevolumens einnehmen können. Je nach Familie handelt es sich dabei um Rotalgen, Chlorophyten, Dinoflagellaten oder Diatomeen. Diese Endosymbionten sind integriert in die Stoffwechselprozesse ihrer Wirte, die durchweg transparente Schalen mit Bauweisen aufweisen, die eine effiziente Nutzung des Lichteinfalls ermöglichen. Diese werden zwar bei asexueller Vermehrung durch Zellteilung mit den Abkömmlingen "geteilt", von sexuellen Abkömmlingen müssen sie aber jeweils neu erworben werden. [1]
Die Foraminiferen "ernten" diese Endosymbionten regelrecht ab und nutzen sie als Nahrungsergänzung oder im Falle von Nahrungsmangel als Reserve. Zugleich gibt es Hinweise auf die Lieferung von Stärke und anderen organischen Substanzen an den Wirt. [5]
Paläontologie
Das verwesungsresistente Gehäuse kann nach dem Absterben der Zelle fossilisieren und auf diese Weise erhalten bleiben. Anhand von fossilen Foraminiferen-Vergesellschaftungen kann man die Umweltbedingungen vergangener Zeiten rekonstruieren und die sie enthaltenden Gesteine relativ datieren (Biostratigraphie)[6] Ab der Kreidezeit stellen planktonische Foraminiferen aufgrund ihrer marinen Lebensweise und somit fast weltweiter Verbreitung wichtige Leitfossilien. Einige fossile Formen traten in solchen Mengen auf, dass sie gesteinsbildend wurden, so die Globigerinida (Globigerinen), die Fusulinen (Fusulinida) und die Nummuliten (Nummulitidae).
Forschungsgeschichte
Foraminiferen wurden vom Menschen lange nur durch ihre fossilierte Schalen wahrgenommen. Frühen Autoren ging selbst das Verständnis ihres Ursprungs als Gehäuse eines Lebewesens noch ab, so deutete Strabo die Nummuliten im Kalk der Pyramiden von Gizeh als Kotreste der Arbeiterschaft und noch im 16. bis 18. Jahrhundert wurde meist davon ausgangen, es handele sich schlicht um Steine. [5]
Antoni van Leeuwenhoek dokumentierte 1700 den Fund einer Foraminiferenschale, „nicht größer als ein Sandkorn“, im Magen einer Garnele. Eine von ihm angefertigte Zeichnung lässt sich sicher als eine Art der Gattung Elphidium bestimmen und er verstand sie als Weichtier. [7] Unabhängig davon äusserte Johann Jakob Scheuchzer zu gleicher Zeit, dass „diese Steine wahrhaft Schnecken und nicht in der Erden durch ich weiß nicht was vor einen Archeum gebildet worden“ [5]. Zwar war ihre systematische Zuordnung falsch, immerhin jedoch war nunmehr der Charakter der Foraminiferen als Lebewesen erkannt.
Die Pionierarbeit von Janus Plancus, der 1739 einige Foraminiferen vom Strand von Rimini beschrieb, diente Carl von Linné 1758 als Grundlage für seine Platzierung der Arten bei den Perlbooten (Nautilus). Spätere Arbeiten um die Mitte des 18. Jahrhunderts (Gualtieri 1742, Lesser 1744, Ledermüller 1761, Martini 1769) folgten ihm darin oder gliederten sie in die heute zu den Geradhörnern gehörende Gattung Orthoceras ein. Als eigenständige systematische Gruppe wurden sie jedoch erst 1826 durch die Erstbeschreibung der Foraminiferen von Alcide d'Orbigny manifest. Sah er sie ursprünglich noch als Ordnung der Kopffüßer, so erhob er sie 1839 zu einer eigenen Klasse. Dem Voraus gingen die ersten Erkenntnisse zur Biologie der Foraminiferen, insbesondere zum Aufbau des eigentlichen „Körpers“ von Félix Dujardin. D’Orbigny war es auch, der mit der paläontologischen Erforschung der Foraminiferen begann, während zugleich vornehmlich britische Forscher erste ökologische Forschungen durchführten. [5]
Systematik
Adl et al. ordnen die Foraminiferen als eines der fünf Taxa innerhalb der Rhizaria ein, deren bei weitem größte Gruppe sie sind [8]. Auf der Basis von molekularbiologisch ermittelten Stammbäumen stellen die Foraminiferen die Schwestergruppe der Gromia dar und bilden entsprechend mit diesen wahrscheinlich ein gemeinsames Taxon.
Die innere Systematik der Gruppe hingegen gilt unter molekulargenetischen Gesichtspunkten noch als weitgehend unklar. Vor allem die Tatsache, dass die dazu notwendige DNA meist nur in unzureichender Menge gewonnen werden kann, da die meisten Foraminiferen sich in Labors nicht kultivieren lassen und nur von extrem wenigen Arten überhaupt DNA zur Verfügung steht, erschwert umfangreiche und repräsentative Untersuchungen. [9] Auch technische Hindernisse machen die Erstellung phylogenetischer Stammbäume zu schwierigen Aufgaben: Sogenannte Long-branch attraction-Artefakte führen im Rahmen der für Untersuchungen häufig gebrauchten SSU rDNA oft zu schweren statistischen Fehlern, daher konnte erst der Gebrauch experimenteller Markergene (Aktin-[10], RNA-Polymerase II-Gen [9]) die anfänglichen Ergebnisse stabilisieren. Deutlich zeichnet sich in allen Untersuchungen ab, dass die Ordnungen Allogromiida und Astrorhizida zusammen mit einigen, oft als Athalamidae geführten schalenlosen Foraminiferen eine paraphyletische Gruppe bilden sowie dass die Miliolida aus ihnen hervorgingen. Auch sind die Globigerinida ebenso wie die Buliminida wohl ein Teil der Rotaliida.
Alle bisherigen Systematiken basieren daher, auch aufgrund der großen Anzahl der ausschließlich durch ihre Gehäuse bekannten fossilen Arten, auf morphologischer Grundlage [11]. Die derzeit aktuelle umfassende Systematik der Foraminiferen geht auf Alfred R. Loeblich und Helen Tappan zurück und wurde 1992 vorgestellt. Sie diente auch als Grundlage für die modifizierte und ergänzte Systematik von Barun K. Sen Gupta aus dem Jahre 2002, die hier herangezogen wird. Die Foraminiferen werden dort als Klasse verstanden und in 16 Ordnungen untergliedert († = ausgestorben). [12] Sie umfassen rund 10.000 rezente und 40.000 fossil bekannte Arten[13] in rund 65 Überfamilien und 300 Familien, rund 150[14] davon rezent.
- Ordnung Allogromiida
- Ordnung Astrorhizida
- Ordnung Lituolida
- Ordnung Trochamminida
- Ordnung Textulariida
- Ordnung Fusulinida †
- Ordnung Miliolida
- Ordnung Carterinida
- Ordnung Spirillinida
- Ordnung Lagenida
- Ordnung Buliminida
- Ordnung Rotaliida
- Ordnung Globigerinida
- Ordnung Involutinida
- Ordnung Robertinida
- Ordnung Silicoloculinida
Einzelnachweise
- ↑ a b c d e f Klaus Hausmann, Norbert Hülsmann, Renate Radek: Protistology, 3. Auflage, 2003, ISBN 3-510-65208-8, S. 129-134
- ↑ Jan Pawlowski, Maria Holzmann, Cedric Berney, Jose Fahrni, Andrew J. Gooday, Tomas Cedhagen, Andrea Habura, Samuel S. Bowser: The evolution of early Foraminifera, In: Proceedings of the National Academy of Sciences of the USA, Vol. 100, S. 11494–11498, 2003
- ↑ B. Ziegler: Einführung in die Paläobiologie Teil 2 - Protisten, Spongien, etc., 1983, S. 35
- ↑ a b c d Susan T. Goldstein: Foraminifera: A Biological Overview In: Barun K. Sen Gupta (Hrsg.): Modern Foraminifera, 1999, S. 37-57, ISBN 0-412-82430-2
- ↑ a b c d e Klaus Nuglisch: Foraminiferen - marine Mikroorganismen, Wittenberg, 1985
- ↑ Spiegel online, zuletzt eingesehen am 11.01.2007.
- ↑ Barun K. Sen Gupta: Introduction to modern Foraminifera In: Barun K. Sen Gupta (Hrsg.): Modern Foraminifera, 1999, S. 3-6, ISBN 0-412-82430-2
- ↑ Adl, Sina M., Leander, Brian S., Simpson, Alastair G. B., Archibald, John M., Anderson, O. Roger., Bass, David, Bowser, Samuel S., Brugerolle, Guy, Farmer, Mark A., Karpov, Sergey, Kolisko, Martin, Lane, Christopher E., Lodge, Deborah J., Mann, David G., Meisterfeld, Ralf, Mendoza, Leonel, Moestrup, Øjvind, Mozley-Standridge, Sharon E., Smirnov, Alexey V. and Spiegel, Frederick (2007) Diversity, Nomenclature and Taxonomy of Protists, In: Systematic Biology, 56:4, 685
- ↑ a b David Longet, Jan Pawlowski: Higher-level phylogeny of Foraminifera inferred from the RNA polymerase II (RPB1) gene In: European Journal of Protistology, 43 (2007), S. 171–177
- ↑ Jerome Flakowski, Ignacio Bolivar, Jose Fahrni, Jan Pawlowski: Actin Phylogeny Of Foraminifera In: Journal of Foraminiferal Research, 2005, Band 35, S. 93-102
- ↑ Sina M. Adl, Alastair G. B. Simpson, Mark A. Farmer, Robert A. Andersen, O. Roger Anderson, John A. Barta, Samual S. Bowser, Guy Brugerolle, Robert A. Fensome, Suzanne Fredericq, Timothy Y. James, Sergei Karpov, Paul Kugrens, John Krug, Christopher E. Lane, Louise A. Lewis, Jean Lodge, Denis H. Lynn, David G. Mann, Richard M. McCourt, Leonel Mendoza, Øjvind Moestrup, Sharon E. Mozley-Standridge, Thoams A. Nerad, Carol A. Shearer, Alexey V. Smirnov, Frederick W. Spiegel, Max F. J. R. Taylor: The New Higher Level Classification of Eukaryotes with Emphasis on the Taxonomy of Protists. The Journal of Eukaryotic Microbiology 52 (5), 2005; Seite 418
- ↑ Barun K. Sen Gupta: Systematics of modern Foraminifera In: Barun K. Sen Gupta (Hrsg.): Modern Foraminifera, 1999, S. 7-37, ISBN 0-412-82430-2
- ↑ Samuel S. Bowser, Andrea Habura, Jan Pawlowski: Molecular evolution of Foraminifera In: Laura Katz Olson, Laura A. Katz, Debashish Bhattacharya: Genomics and Evolution of Microbial Eukaryotes, 2006, S. 78-94, ISBN 0-19-856974-2
- ↑ Pamela Hallock: Symbiont-bearing Foraminifera In: Barun K. Sen Gupta (Hrsg.): Modern Foraminifera, 2002, S. 123, ISBN 978-1-4020-0598-5
Weblinks
Wikimedia Foundation.