- Gastrulation
-
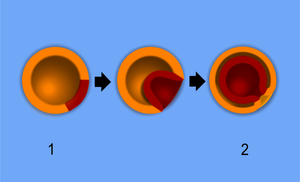 1 - Blastula, 2 - Gastrula; orange - Ektoderm, rot - Entoderm
1 - Blastula, 2 - Gastrula; orange - Ektoderm, rot - Entoderm
Gastrulation - von griech.: gaster (Magen) - bezeichnet eine Phase der Embryogenese der vielzelligen Tiere, zu denen auch der Mensch gehört. Dabei stülpt sich die Blastula ein und es kommt zur Ausbildung der Keimblätter. Vorausgegangen ist die Ausbildung des Blasenkeimes, die s.g. Blastulation. Dem britischen Entwicklungsbiologen Lewis Wolpert wird folgendes Zitat über die Gastrulation zugeschrieben: „Es ist nicht die Geburt, die Hochzeit oder der Tod, sondern die Gastrulation, welche in Wirklichkeit der wichtigste Zeitpunkt in deinem Leben ist.“
Inhaltsverzeichnis
Grundsätzlicher Vorgang
Die Gastrulation verläuft zwar bei allen vielzelligen Tieren (genauer gesagt bei den Gewebetieren, Eumetazoa) prinzipiell ähnlich, jedoch gibt es sehr deutliche Unterschiede sowohl beim Vorgang selbst als auch beim Ergebnis der Gastrulation. Im Laufe der Evolution haben sich die Vorgänge neuen Erfordernissen angepasst und vom Grundmuster abgewandelt, so dass sich gerade die Gastrulation der Säuger nur dann auf die gemeinsame Urform zurückführen lässt, wenn man die evolutionäre Abstammung von eierlegenden Tieren wie Vögeln und Reptilien beachtet und deren Embryologie als Bindeglied betrachtet.
Bilateralsymmetrische Tiere bilden drei Keimblätter aus (triploblastisch), das äußere Ektoderm, das innere Entoderm und das mittlere Mesoderm. Die dem veralteten, taxonomisch nicht korrekten Phylum der Radiata Hohltiere zugeordneten Cnidaria und Ctenophora bilden nur zwei Keimblätter aus (trifft nicht bei allen zu), nämlich Entoderm und Ektoderm (sie sind diploblastisch).
Ausgangslage ist die Blastula bei Vielzellern und niederen Säugetieren bzw. die Blastocyste bei höheren Säugetieren (u.a. beim Mensch), eine Hohlkugel aus einer Zellschicht, die, grundsätzlich und vereinfacht gesagt, zunächst in den zweischichtigen "Becherkeim", die Gastrula, umgebildet wird. Das Innere der beiden primären Keimblätter ist das Entoderm, das Äußere das Ektoderm. Eine Öffnung des Entoderms nach außen bezeichnet man als Urmund (Blastoporus), das Entoderm selbst stellt den Urdarm (Archenteron) dar. Die Entwicklung des Mesoderms erfolgt zeitgleich oder leicht zeitversetzt zur Bildung der beiden primären Keimblätter.
Die weitere Entwicklung des Urmunds teilt die bilateralsymmetrischen Tiere (Bilateria) in zwei Stammgruppen ein: Bei den Urmündern (Protostomier) bildet der Urmund auch tatsächlich den späteren Mund, während bei den Neumündern (Deuterostomier), zu denen die Wirbeltiere und auch der Mensch zählen, sich der Urmund mehr oder weniger deutlich zum After entwickelt, während der spätere Mund bei vollendeter Gastrulation auf der anderen Seite der Blastula "durchbricht".
Der Ablauf der Gastrulation weist wie schon erwähnt deutliche Unterschiede zwischen den Spezies auf, kann aber auf eine Reihe von Grundbewegungen zurückgeführt werden:
- Invagination, Einstülpung des prospektiven ("voraussichtlichen") Entoderms in den inneren, flüssigkeitsgefüllten Hohlraum (das Blastocoel) der Blastula: Durch Verformung von Zellen eines Poles der Blastula wird ein Teil der Außenwand eingestülpt; das Ergebnis sieht aus wie ein luftleerer, auf einer Seite eingedellter Fußball. Der innenliegende Anteil wird ab diesem Zeitpunkt als Entoderm, der außenliegende als Ektoderm bezeichnet. Der innere Hohlraum der Blastula, die primäre Leibeshöhle, wird bei diesem Vorgang eingeengt; die aus dem Entoderm bestehende "Delle" wird als Urdarm oder Archenteron bezeichnet.
- Involution, Einrollen des prospektiven Entoderms
- Ingression (Immigration), Einwandern von Zellen des prospektiven Entoderms
- Delamination, Zellen der Blastula schnüren die Zellen des prospektiven Entoderms ins Blastocoel ab
- Epibolie, im Prinzip eine Invagination. Bei dotterreichen Eiern findet diese statt, indem das prospektive Ektoderm das prospektive Entoderm überwächst.
In der Regel überschneiden sich Gastrulation und Beginn folgender Prozesse wie die Neurulation.
Beschreibung der Gastrulation an Beispielen
Amphibien
Amphibien sind das klassische Beispiel für die sehr ursprüngliche Wirbeltierentwicklung. An einer bestimmten Stelle der Blastula erhöht sich die Rate der Zellteilung lokal, so dass sich ein Zellhaufen entwickelt, der sich wie eine Delle eines luftleeren Fußballs in die primäre Leibeshöhle (Blastocoel) der Blastula einstülpt. Ursache sind die erhöhte Teilungsrate, aber auch amöboide Bewegungen der Zellen selbst. Es entsteht eine doppelwandige Hohlkugel, deren innere Wand, das Entoderm, dort, wo sie sich eingestülpt hat, eine Öffnung nach außen aufweist, den Urmund. Die äußere Wand ist das Ektoderm.
Durch den Urmund ist nun das spätere hintere Körperende markiert, da sich der Urmund zum After entwickelt. Während das Entoderm noch einwächst, wachsen entlang der Mittellinie des Keims aus dem sich einstülpenden Entoderm beidseits dieser Mittellinie Zelllappen in den Raum zwischen Ento- und Ektoderm. Diese Lappen wachsen von der Rückseite des Keimes zwischen den primären Keimblättern beidseits bauchwärts um das Entoderm, den Urdarm, herum, bis sie sich auf der Mittellinie auf der Bauchseite treffen. Sie bilden das Mesoderm. Im Folgenden bilden und erweitern sich auf ganzer Länge Spalträume innerhalb des Mesoderms, die die sekundäre Leibeshöhle (Coelom) bilden.
Zum Ende der Gastrulation hat also der Embryo eine klare Orientierung durch eine Längsachse mit einem Vorder- und Hinterende, transversal durch die Mittellinie zwischen den Blättern des Mesoderms, und im Querschnitt durch Bauch- und Rückseite.
Der nächste Schritt der Wirbeltierentwicklung ist die Bildung der Chorda dorsalis als dorsaler, längsgerichteter Mesodermstrang, der sich aus dem Dach des Urdarms bildet, wo die beiden Mesodermblätter auseinanderwuchsen. Die Chorda induziert weitere grundlegende Prozesse wie die Neurulation.
Vögel
Während Amphibien auf die Fortpflanzung und Entwicklung im Wasser angewiesen sind, da ihre wasserdurchlässigen, empfindlichen Eier an Land vertrocknen würden, haben Vögel und Reptilien sich an die Bedingungen an Land angepasst und ihre Eier durch extraembryonale Membranen widerstandsfähiger gemacht. Ein bedeutender Fortschritt ist das Amnion, das den Embryo umgibt, weshalb Vögel und Reptilien gemeinsam mit den Säugetieren als Amnioten bezeichnet werden. Das Vogelei enthält im Unterschied zum Amphibienei eine riesige Eizelle, die fast nur aus Dotter besteht (bei einem unbefruchteten Hühnerei ist das Eigelb tatsächlich die Eizelle, während das Eiklar eine Nährlösung für den Embryo darstellt). Dadurch, dass die Zellen sich in Anwesenheit von Dotter nur eingeschränkt bzw. gar nicht teilen können, kann sich keine kugelförmige Blastula entwickeln, sondern nur eine flache, aus meroplastischer (partieller) Furchung entstandene Keimscheibe, die ventral offen auf dem animalen Pol der riesigen Dotterkugel sitzt und aus zwei übereinanderliegenden Zellschichten, dem Epiblast und dem Hypoblast, besteht, welche gemeinsam das Blastocoel einschließen (somit stellen sie die eigentliche Blastula dar). Am hinteren Pol dieser Keimscheibe bildet sich eine ausgezogene, lange Rinne, die Primitivrinne. Sie reicht ungefähr von der Mitte der halbkugelartigen Keimscheibe zum Keimende. In diese Rinne wandert nun von beiden Seiten Zellmaterial aus dem Epiblast in die primäre Leibeshöhle ein: Zunächst bilden Zellen auf dem Dottersack das Entoderm und verschieben dabei die Zellen des Hypoblasten nach unten, dann stülpen sich die in die Primitivrinne einsinkenden Zellen nach beiden Seiten um und bilden das zwischen den primären Keimblättern hängende Mesoderm. Die übrigen Zellen des Epiblast bilden das Ektoderm. Der Embryo besteht somit vollständig aus Zellen des Epiblasten. Der "Urdarm" besteht zunächst nur aus einem kleinen Raum zwischen Entoderm und Dotter und ist zum Dotter hin offen; die Darmanlage bildet sich jedoch noch, während der Embryo sich durch seitliche Faltung von der Dottermasse abhebt. Dabei schließt sich der Darm und reduziert seine Verbindung zum Dottersack auf einen Dotterstiel, der vorwiegend aus hypoblastischen Zellen besteht und sich in der Mitte des Embryos befindet (ähnlich der Nabelschnur bei Säugern).
Sobald alle drei Keimblätter gebildet sind, beginnen sie, zunächst das Ektoderm, dann das Entoderm, dann zwischen beiden hindurch das Mesoderm, um den riesigen Dotter herumzuwachsen, und werden zu extraembryonalen Membranen. Die hypoblastischen Zellen, die nach Abschluss dieses Vorgangs den Dotter umschließen, bilden ein vorübergehendes embryonales Hilfsorgan, den Dottersack, der den Embryo ernährt. Die äußeren Epiblasten bilden das Chorion, welches den Embryo und die anderen Membranen im extraembryonalen Coelom einschließt. Aus weiteren Zellen entsteht die Allantois, eine Ausbuchtung des Enddarms, die Abfallprodukte des embryonalen Stoffwechsels aufnimmt. Nur der kleine Teil oben auf dem Dotter, an dem die Keimblätter sich ausbildeten, wird sich als Embryo weiterentwickeln. Diese Unterscheidung findet sich auch in der Entwicklung der Säuger wieder.
Über die Gastrulation hinaus
Während der Embryo die Chorda dorsalis aus dem Mesoderm und das Neuralrohr aus dem Ektoderm bildet, finden noch zwei wesentliche Entwicklungen im extraembryonalen Gewebe statt. Der Dotter ist jetzt vom dreischichtigen Dottersack aus allen drei Keimblättern umwachsen. Zum einen falten sich Ektoderm und Mesoderm über dem Embryo nach oben, treffen sich in der Mitte, verbinden sich, und formen so eine schützende Höhle über dem Embryo, die Amnionhöhle, die den Embryo vor Austrocknung und mechanischen Erschütterungen schützen soll. Zum anderen entsteht im Mesoderm eine Höhle, so dass das Mesoderm vollständig in zwei Blätter zerfällt, zwischen denen die sekundäre Leibeshöhle oder Coelom entsteht. Sobald an der Grenze von intra- und extraembryonalen Gewebe, dort, wo sich die Amnionhöhle aufgefaltet hat, die Verbindung der intra- und extraembryonalen Coelome abreißt, entsteht eine Form des Embryos mit einer äußeren Hülle (von außen nach innen Ektoderm, Mesoderm), einer extraembryonalen Leibeshöhle, der Amnionhöhle (Mesoderm, Ektoderm, Amnionhöhle, Embryo) und darunter des Dottersacks (Mesoderm, Entoderm, nach oben offene Verbindung zum Urdarm des Embryos), die man genau so in der Embryonalentwicklung der Säuger wiederfindet, ohne aber direkt die Ähnlichkeit zum Grundmuster der Amphibien erkennen zu können.
Säugetiere und der Mensch
Verhältnis zur Embryonalentwicklung der Vögel
Gerade bei der frühen Embryonalentwicklung wird die stammesgeschichtliche Verwandtschaft der Säugetiere mit den eierlegenden Vögeln und Reptilien deutlich. Zwar legen Säugetiere (mit den Ursäugern als Ausnahme) keine Eier. Dennoch sind sowohl der Dottersack als auch Amnion, Chorion und Allantois immer noch integrale Bestandteile der Embryonalentwicklung: Die Evolution kann dieses Hilfsorgan nicht einfach wieder abschaffen, der Dottersack beinhaltet zwar kein Dotter mehr, aber dennoch laufen Prozesse wie die erste Blutbildung und das Entstehen der Urkeimzellen im Dottersack ab. Das ererbte genetische Programm wird also weiterentwickelt und teilweise umgewandelt. Gerade bei den höheren Säugetieren wie den Primaten sind einige Prozesse so abgewandelt, dass das Ergebnis zwar das gleiche, der Weg dorthin aber wesentlich abgekürzt ist.
Späte Blastozyste
Die Blastozyste ist - scheinbar im Gegensatz zu den Vögeln, aber wie bei der Blastula der Amphibien - eine Hohlkugel. Allerdings wird sich schon bald zeigen, dass sich auch die Säugetiere aus einer Keimscheibe entwickeln. Die Zellen der Blastozyste haben sich zu zwei Linien differenziert, zum äußeren Trophoblasten und zum Embryoblasten (für eine Erklärung siehe Blastozyste). Aus dem Trophoblasten wird das für Säugetiere typische embryonale Hilfsorgan entstehen, die Plazenta, während aus dem Embryoblasten, einem Zellhaufen an einer Stelle an der Innenseite der Hohlkugel, der eigentliche Embryo entsteht. Man kann sich vorstellen, dass der (nicht mehr vorhandene) Dotter im Hohlraum der Blastozyste liegt und der darauf liegende Embryoblast der nach unten offenen Keimscheibe der Vögel entspricht.
Zweiblättrige Keimscheibe
Beim Menschen erfolgt die Ausbildung zweier Keimblätter einfach dadurch, dass sich das Zellmaterial des Embryoblasten zum Einen zu einem epithelartigen, zylindrischen Verband differenziert, das Ektoderm, während sich zum Anderen an der Grenze der Keimscheibe gegen die Blastozystenhöhle eine Schicht kleinerer, vieleckiger Zellen, das Entoderm, ausbildet.
Während bei einigen Säugetieren (Hund und Katze) die Bildung der Amnionhöhle noch genau wie bei Vögeln über die Entstehung und Verschluss von Amnionfalten geschieht, ist dieser Prozess bei höher entwickelten Säugern, speziell beim Menschen, abgekürzt und vereinfacht: Im Ektoderm entstehen Spalträume, die konfluieren und die Amnionhöhle bilden. Sie ist nach oben durch eine Schicht dünner Zellen abgegrenzt, die auch aus dem Embryoblasten stammen, nach unten liegt das Ektoderm und somit der Rücken der Keimscheibe (achter Tag beim Menschen).
Aus dem Entoderm wandern Zellen aus, die entlang der Innenseite der Trophoblasten die Blastozystenhöhle auskleiden und so den primären Dottersack ausbilden (Mensch: Neunter Tag).
Die Implantation in der Gebärmutterschleimhaut und die weitere Entwicklung des Trophoblasten zu Zytotrophoblast und Synzytiotrophoblast, auf die in diesem Artikel nicht näher eingegangen wird, geschieht parallel und relativ unabhängig zur hier geschilderten Entwicklung des Embryoblasten, in den ersten Tagen schreitet sie in Bezug auf die Größe sogar wesentlich schneller voran. Das bedeutet, dass um den elften bis zwölften Tag (Mensch) herum der Trophoblast unverhältnismäßig zum Embryoblasten wächst. Er reißt zunächst einen Spaltraum zwischen der entodermalen Auskleidung des primären Dottersacks und dem Trophoblasten auf, der durch ein netzartiges, lockeres Gewebe gefüllt wird, das vom Entoderm abstammt (entodermales Retikulum).
Am dreizehnten Tag (Mensch) entsteht aus der Zone zwischen den beiden Keimblättern extraembryonales Mesoderm. Dieses Gewebe wächst in den oben geschilderten Spaltraum ein, umkleidet zum einen von außen das Gebilde aus Amnionhöhle, Dottersack und dazwischenliegender Keimscheibe, bildet zum anderen eine Schicht an der Innenseite des Trophoblasten. Die zwischen diesen beiden Blättern des extraembryonalen Mesoderms gebildete Höhle entspricht dem extraembryonalen Coelom und wird als Chorionhöhle bezeichnet. Der Embryo ist nur an einem dünnen mesodermalen Haftstiel aufgehängt. Die äußere Hülle aus Trophoblast und Mesoderm, aus der die Plazenta entstehen wird, nennt man Chorion.
Die nun am Ende der zweiten Woche (Mensch) erreichte Form mit der Keimscheibe zwischen Amnionhöhle und Dottersack, einer extraembryonalen Leibeshöhle und der Chorionhülle weist große Ähnlichkeiten mit dem Vogelembryo am Ende der Gastrulation auf, wobei beim Säugerembryo jetzt noch kein intraembryonales, eigentliches Mesoderm entstanden ist. Zwar ist im Vergleich zum Vogel der Weg zu dieser Entwicklungsstufe, was Form und Zeitpunkt der Entwicklungen betrifft, in vielen Punkten unterschiedlich, im Ergebnis aber sehr ähnlich. So lässt sich über die Vögel auch die Verbindung zur Entwicklung ursprünglicher Wirbeltiere wie den Amphibien erkennen.
Dreiblättrige Keimscheibe
In der dritten Woche der Embryonalentwicklung des Menschen bildet sich das intraembryonale Mesoderm, die dritte Keimscheibe, aus. Vom hinteren Pol der Keimscheibe entwickelt sich im Ektoderm eine längliche, schmale, rinnenartige Verdickung, der Primitivstreifen, der nach vorne wächst und etwa auf der Grenze vom hinteren zum mittleren Drittel des Embryos einen rundlichen Abschluss bildet, den Primitivknoten. Zellen wandern vom Ektoderm nach innen auf Primitivstreifen und -knoten zu, steigen ab und bilden eine neue Zellschicht zwischen Ektoderm und Entoderm, in der sie dann wieder zur Seite und nach vorne wandern. Mit Ausnahme eines kleinen runden Areals im vorderen Bereich (der Prächordalplatte) dehnt sich das Mesoderm in der gesamten Keimscheibe zwischen den beiden primären Keimblättern aus und gewinnt auch seitlich Anschluss an das schon länger bestehende extraembryonale Mesoderm.
Im weiteren Verlauf bildet sich aus dem Mesoderm entlang der Mittellinie die Chorda dorsalis, die wiederum weitere Vorgänge wie die Neurulation induziert. Das Mesoderm wird unter anderem die Somiten bilden.
Klinische Bezüge
Die Frühentwicklung der ersten zwei Wochen des menschlichen Keims ist gegenüber schädigenden Einflüssen relativ unempfindlich. Fehlbildungen des Keims und die meisten chromosomalen Aberrationen führen zu einem unbemerkten Abort.
Ganz anders ist die Gefährdung ab dem Beginn der dritten Woche (zwei Wochen nach Befruchtung, ca. vier Wochen nach der letzten Regel), sobald sich der Primitivstreifen (s.o.) bildet. Jedes Organ hat ab dann eine spezifische Phase, in der es für Teratogene besonders empfindlich ist. Speziell bei der Gastrulation gibt es zwei erwähnenswerte Krankheitsbilder: Sirenomelie und Steißbeinteratome.
Tritt eine Störung bei der Bildung des Primitivstreifens auf und ist als Folge nicht genug Mesoderm in der hinteren Hälfte des Embryos vorhanden, kann das sehr seltene Syndrom der Sirenomelie auftreten. Es umfasst Fehlbildungen wie das Zusammenwachsen der unteren Gliedmaßen, Störungen der unteren Wirbelsäule, fehlende Ausbildung der Nieren und Fehlbildung der Genitalorgane.
Reste des Primitivknotens können bei Neugeborenen Tumoren bilden, die Sakrokokzygealteratome oder Steißbeinteratome. Dieses Teratom ist der häufigste Tumor des Neugeborenen (1 auf 37000).
Literatur
- Gilbert, Scott F. (2000): "Developmental Biology", 6th ed., Sinauer Associates, Inc., Sunderland
- Gilbert, Scott F. and Raunio, Anne M. [eds.] (1997): "Embryology: Constructing the Organism", Sinauer Associates, Inc., Sunderland MA
- Schoenwolf, Gary C. (2001): "Laboratory Studies of Vertebrate and Invertebrate Embryos", 8th ed., Prentice Hall, New Jersey
- Wolpert, Lewis et al. (1998): "Principles of Development", Oxford University Press, Oxford - New York - Tokyo
- Scharf, Karl-Heinz und Weber, Wilhelm (1990) : "Materialien für den Sekundarbereich II Biologie: Fortpflanzung und Entwicklung" SCHROEDEL-Verlag, Hannover
- Sadler, Thomas W. (2003), Übersetzung von Ulrich Drews: "Medizinische Embryologie", Thieme Verlag, Stuttgart
- Ax, Peter: "Das System der Metazoa", Spektrum Verlag, Bde 1-3
Wikimedia Foundation.