- Glatte Muskulatur
-
Die glatte Muskulatur ist das kontraktile Gewebe vieler Hohlorgane, Blut- und Lymphgefäße sowie anderer Strukturen bei Tieren und somit auch beim Menschen. Im Gegensatz zur quergestreiften Muskulatur ist sie nicht der willkürlichen Kontrolle unterworfen.
Eine Ausnahme von dieser Systematik bildet der Herzmuskel, der nicht aus glatter Muskulatur besteht, obwohl es sich beim Herzen um ein inneres Hohlorgan handelt.
Inhaltsverzeichnis
Embryologie
Wie auch die Skelettmuskelzellen sind die Zellen der glatten Muskulatur mesodermalen Ursprungs.
Histologie
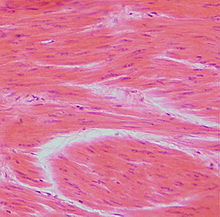 Zellen der Glatten Muskulatur
Zellen der Glatten Muskulatur
Die Zellen sind 30 bis 200 µm (in der Gebärmutter einer Schwangeren bis zu 500 µm) lang und 2 bis 10 µm breit und von einer Basallamina (Proteinschicht) umgeben.[1] Der längliche Zellkern liegt in der Mitte der Zelle. Im Bereich der Kernenden konzentrieren sich Zellorganellen wie Mitochondrien, Ribosomen sowie das raue endoplasmatische Retikulum und Glykogen-Einlagerungen.
Im Gegensatz zur quergestreiften Muskulatur, die der regelmäßigen Anordnung der Myofibrillen ihren Namen verdankt, existiert eine derartige Anordnung von Filamenten in der glatten Muskelzelle nicht. Lichtmikroskopisch erscheint das Zytoplasma homogen. Hauptsächliche Filamente sind Aktin, Myosin und zusätzlich Intermediärfilamente aus der Gruppe der Desmine (Desmin, Vimentin). Ein einzelnes Myosinfilament wird dabei von 13 bis 14 Aktinfilamenten zu einer Art „Bündel“ gruppiert. Die Aktin- und Intermediärfilamente befestigen sich an sogenannten „dense bodies“ (auch Verdichtungszone genannt), welche mit den Z-Scheiben der quergestreiften Muskulatur vergleichbar sind, und an Anheftungsplaques am Rand der Zelle. Diese Anordnung der kontraktilen Elemente bedingt eine stärkere Fähigkeit zur Verkürzung der Muskelzelle, als dies beim quergestreiften Muskel möglich ist.
Eine Besonderheit dieses Muskeltyps stellen die sogenannten Caveolae dar, Einstülpungen der Zellmembran (Sarkolemm), welche mit der extrazellulären Matrix in Kontakt stehen und vermutlich an der elektromechanischen Kopplung beteiligt sind.
Formen
Durch strukturelle Unterschiede und die daraus resultierenden funktionellen Unterschiede, ist eine Unterteilung des glatten Muskelgewebes in den Single unit- und den Multi unit-Typ möglich. Mischformen sind v. a. in der Gefäßmuskulatur häufig.
Single unit-Typ
Der Single unit-Typ bildet durch gap junctions (Nexus) elektrisch gekoppelte Zellverbände aus. Über die gap junctions findet ein Austausch von Ionen und Second-Messenger-Molekülen statt, was eine schnelle Ausbreitung der Erregung und somit eine nahezu synchrone Kontraktion des Zellverbandes („funktionelles Synzytium“) ermöglicht. Die Muskelzellen werden dabei nicht durch nervale Strukturen erregt, sondern durch die spontane Depolarisation morphologisch nicht abgrenzbarer Schrittmacherzellen (myogenener Tonus). Eine Modulation durch Fasern des vegetativen Nervensystems ist jedoch möglich.
Der Single unit-Typ kommt bevorzugt in der Muskulatur des Darms, der Gebärmutter (Uterus) und den Harnleitern (Ureter), teilweise aber auch in der muskulösen Wand größerer Gefäße vor.
Multi unit-Typ
Von den Nachbarzellen abhängige Kontraktionen finden beim Multi unit-Typ nicht bzw. nur sehr begrenzt statt. Jede Muskelzelle wird durch Nervenfasern des Vegetativen Nervensystems, welche in unmittelbarer Umgebung der Zelle aus Varikositäten Transmitter abgeben („en-passant-Synapse“), gesondert innerviert (neurogener Tonus).
Dieser Zelltyp kommt u. a. im Musculus arrector pili des Haares, den inneren Augenmuskeln, dem Samenleiter (Ductus deferens), im Bronchialsystem und in Gefäßen vor.
Kontraktion
Der Kontraktionsmechanismus der glatten Muskulatur entspricht im Wesentlichen der Muskelkontraktion der quergestreiften Muskulatur. Der Gleitfilament-Mechanismus ist jedoch deutlich langsamer, verbraucht aber auch weniger Sauerstoff und weniger Energie in Form von ATP.
Ablauf
Kontraktionen der glatten Muskulatur können durch drei Mechanismen bewirkt werden, die zum Anstieg der intrazellularen Calciumkonzentration führen.
- Elektromechanische Kopplung: Durch Aktionspotentiale oder länger anhaltende Depolarisation der Zellmembran öffnen sich spannungsabhängige Calciumkanäle.
- Pharmakomechanische Kopplung: Rezeptorgesteuerte Calciumkanäle reagieren auf Botenstoffe wie Noradrenalin, Acetylcholin, Angiotensin, Histamin oder Arzneistoffe und öffnen sich. Zudem können rezeptorvermittelt weitere Calciumionen aus dem sarkoplasmatischen Reticulum freigesetzt werden.
- Mechanische Dehnung bewirkt die Öffnung dehnungsabhängiger Calciumkanäle.[1]
Durch die auslösenden Vorgänge kommt es zu einem Anstieg der Ca2+-Konzentration im Zytosol, bedingt durch hereinströmende Ionen aus dem Extrazellularraum und zu einem deutlich geringeren Anteil aus dem sarkoplasmatischen Retikulum. Bis zu vier Ca2+-Ionen binden sich an das Protein Calmodulin. Der entstandene Ca2+-Calmodulin-Komplex führt zu einer Aktivierung des Enzyms myosin light chain kinase (MLCK). Dieses Enzym spaltet in der aktivierten Form ATP und phosphoryliert die leichte Kette des Myosinmoleküls. Durch die Phosphorylierung kann der Querbrückenzyklus ablaufen.
Die Latenzzeit, welche die Zeitspanne zwischen aktivierendem Signal und Kontraktionsantwort des Muskels umfasst, beträgt bei diesem Muskeltyp rund 300 ms und kommt größtenteils durch die Diffusion des Calciums in der Zelle und den beschriebenen Mechanismus der Aktivierung zustande.
Insgesamt gesehen wird bei der glatten Muskulatur eine höhere Verkürzung erreicht als bei der quergestreiften, dies kommt durch die andere Anordnung der Myosinköpfchen zustande.
Relaxation
Zur Entspannung (Relaxation) kommt es durch einen sinkenden Ca2+-Spiegel, ausgelöst durch das Ausbleiben von Nervenreizen bzw. anderer erregender Vorgänge. Calcium wird durch Na+/Ca2+-Antiporter und Ca2+-ATPasen aus dem Innenraum der Zelle zurück in den Extrazellulärraum bzw. das sarkoplasmatische Retikulum transportiert. Der Ca2+-Calmodulin-Komplex dissoziiert und das Enzym Myosin-Leichte-Ketten-Phosphatase (MLCP) dephosphoryliert die leichten Ketten des Myosinmoleküls.
Stickstoffmonoxid (NO), das konstitutiv vom Gefäßendothel gebildet wird, kann in die benachbarten Glattmuskelzellen diffundieren und dort die lösliche Guanylatcyclase aktivieren. Der konsekutive Anstieg im cGMP-Spiegel führt zu einer Aktivierung der Proteinkinase G, die die Myosinphosphatase durch Phosphorylierung aktiviert und damit zur Relaxation der Glattmuskelzellen führt.
Funktion
Glatte Muskulatur kann aufgrund ihrer Struktur und der beschriebenen Vorgänge bei der Kontraktion einen langanhaltenden Tonus (tonische Dauerkontraktion) aufrechterhalten. Sowohl die Peristaltik in Magen, Darm und Harnwegen als auch die Blutdruckregulation in den Innenwänden der Arterien beruhen auf der Wirkung glatter Muskulatur. Während der Geburt ermöglicht sie die rhythmische Kontraktion (phasisch-rhythmische Kontraktion) der Gebärmutter (Wehen).
Matrixproduktion
Glatte Muskelzellen sind zur Synthese von Kollagen und anderen Bestandteilen der extrazellulären Matrix, wie z. B. Proteoglykanen, Elastin und Laminin, befähigt.
Siehe auch
Einzelnachweise
- ↑ a b Mutschler APP des Menschen ISBN 978-3-8047-2342-9
Kategorien:- Histologie der Binde- und Stützgewebe
- Glatte Muskulatur
Wikimedia Foundation.